
Infection of Pro- and Anti-Inflammatory Macrophages by Wild Type and Vaccine Strains of Measles Virus: NLRP3 Inflammasome Activation Independent of Virus Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Viruses, Virus Infection and Inflammasome Activation
2.3. Flow Cytometry
2.4. Plaque Assay for Infectious Virus
2.5. Cytokine and LDH Assays
2.6. Statistical Analysis
2.7. Ethical Statement
3. Results
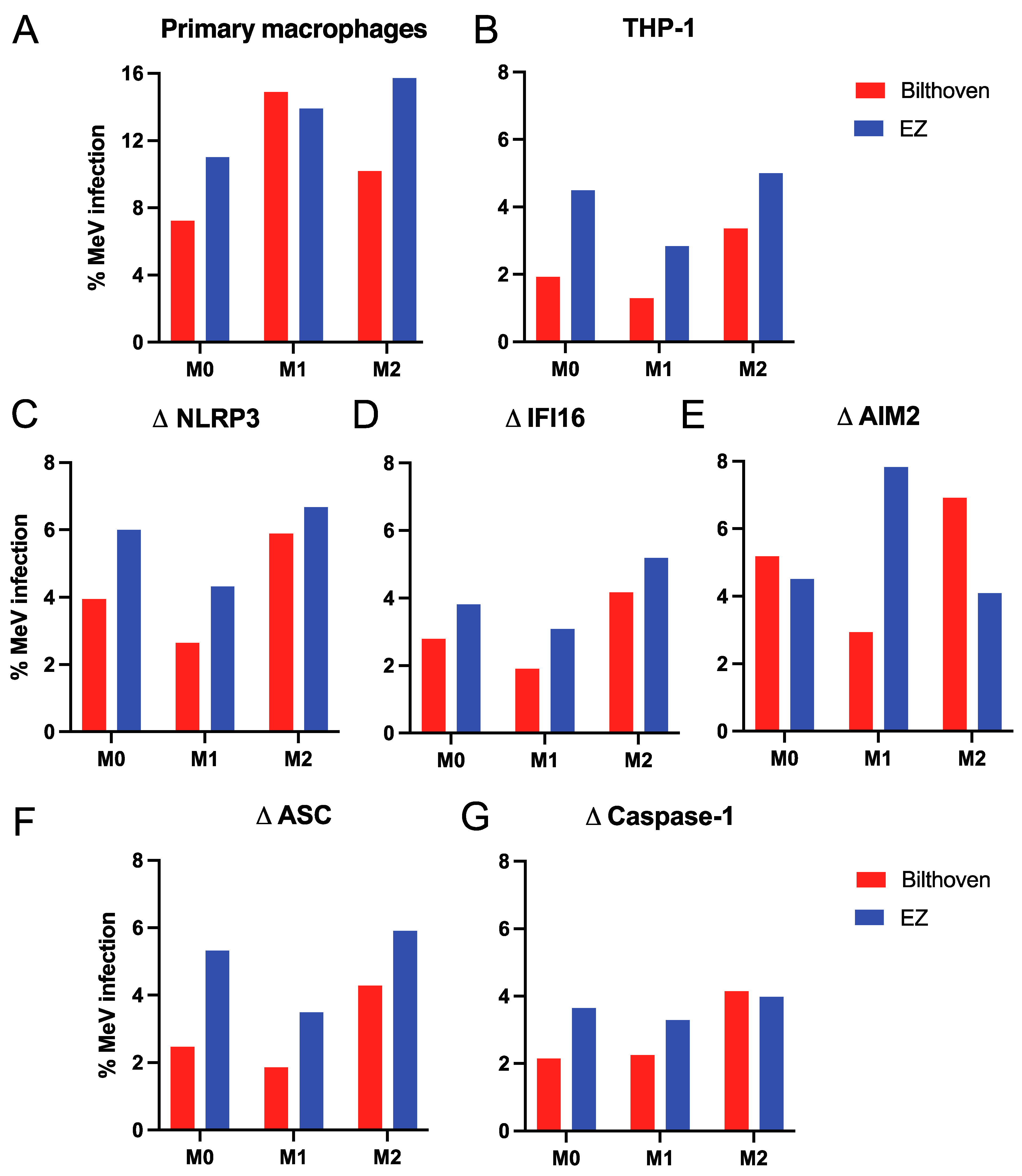
3.1. Infection of Differentiated Primary Human Macrophages and THP-1 Cells with WT and Vaccine Strains of MeV
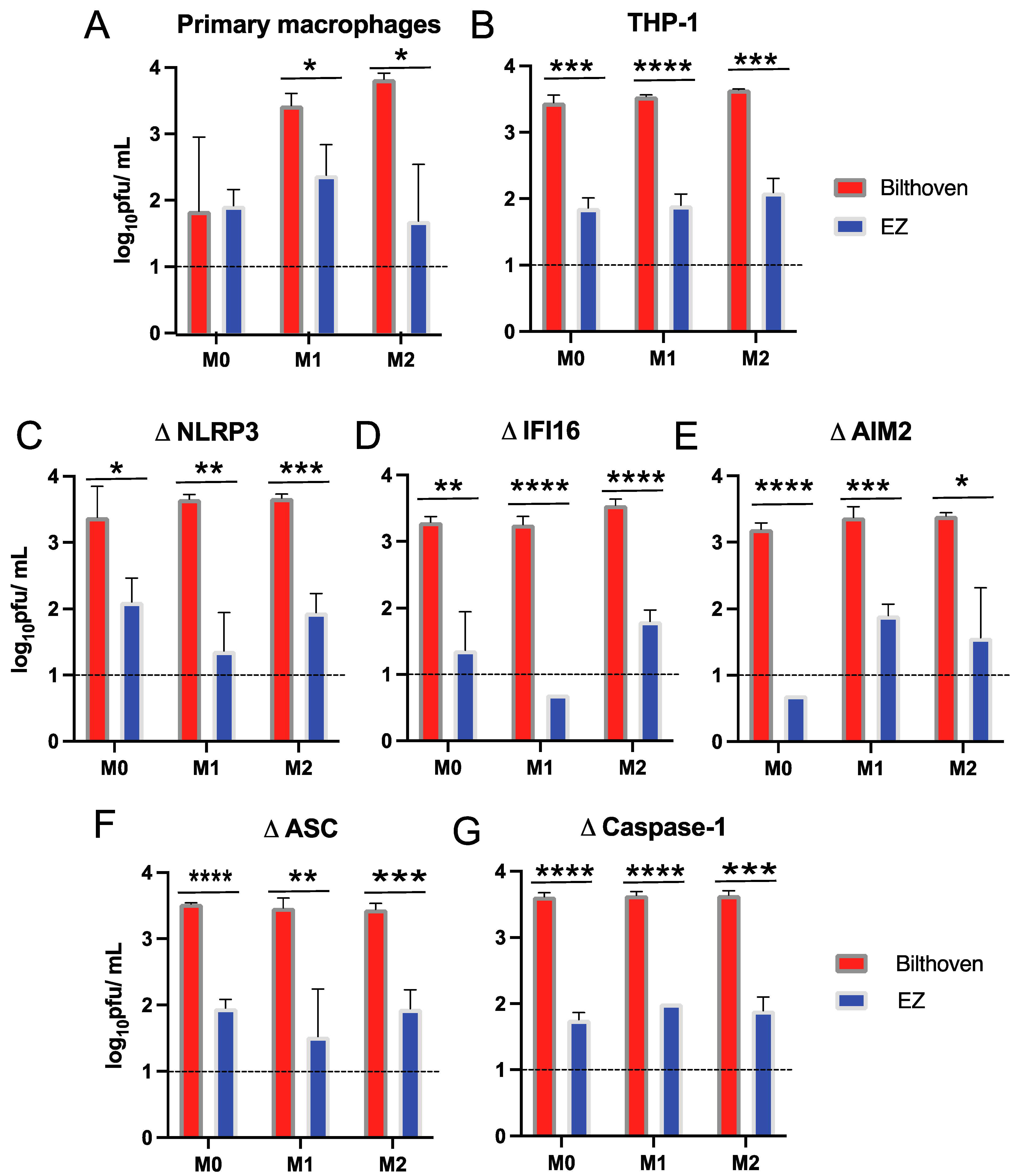
3.2. Production of Infectious Virus by Differentiated Primary Human Macrophages and THP-1 Cells Infected by WT and Vaccine Strains of MeV
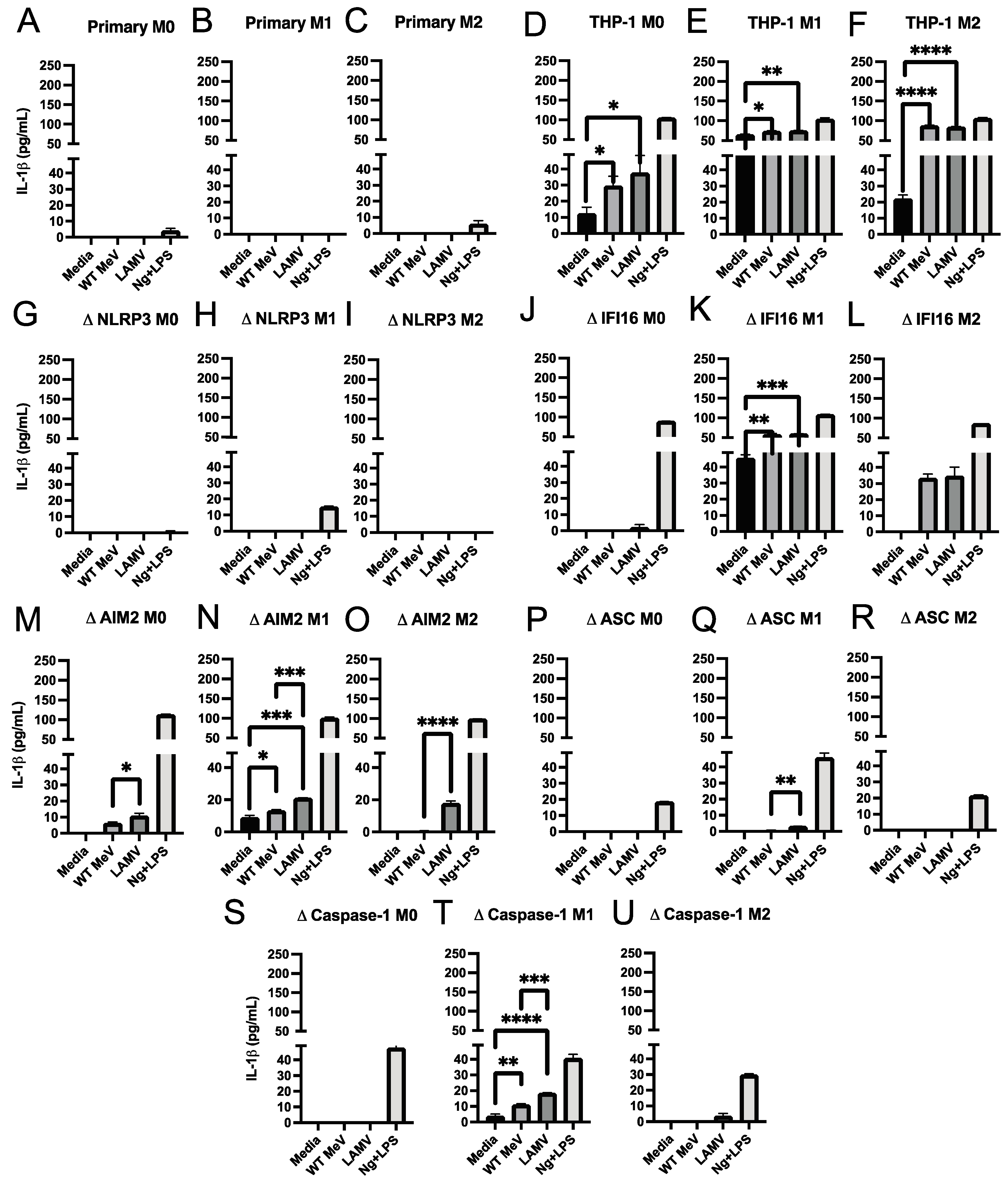
3.3. Inflammasome-Induced Release of IL-18 and IL-1β from Macrophages Infected with LAMV and WT Strains of MeV
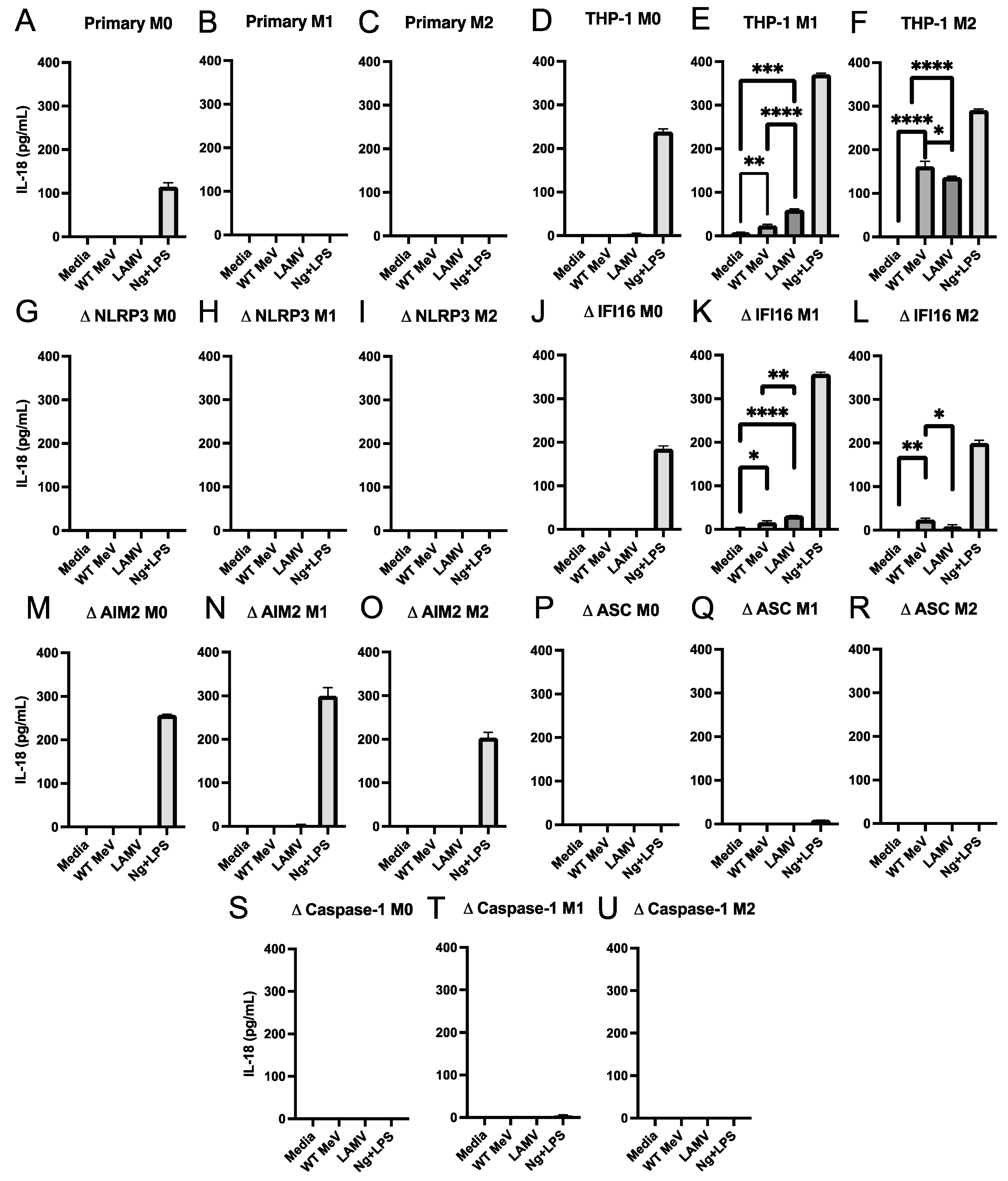
3.4. Effect of Inflammasome Component Deficiencies on MeV-Induced Release of IL-18 and IL-1β from THP-1 Macrophages
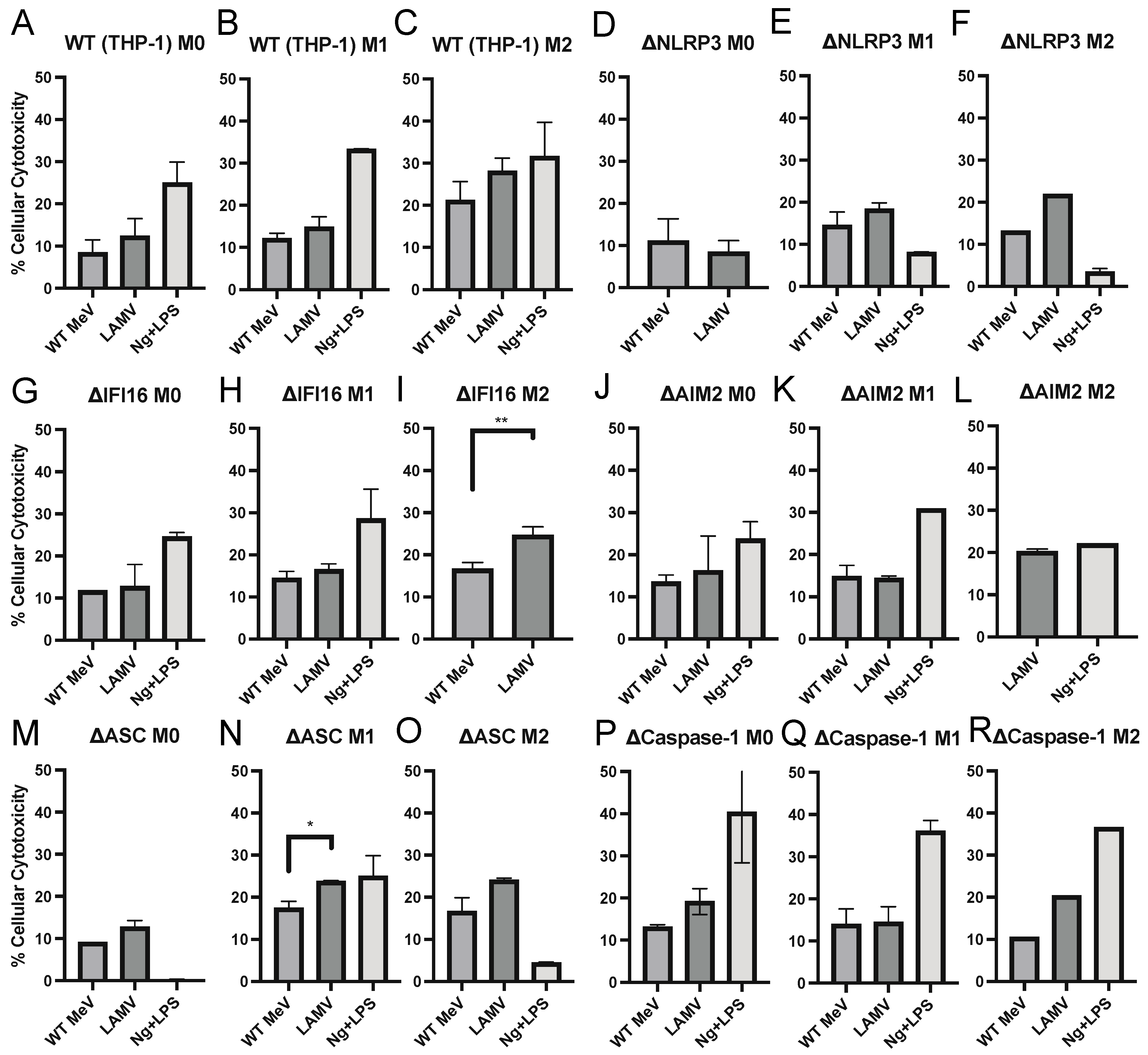
3.5. Cell Death of MeV-Infected Differentiated THP-1 Cells
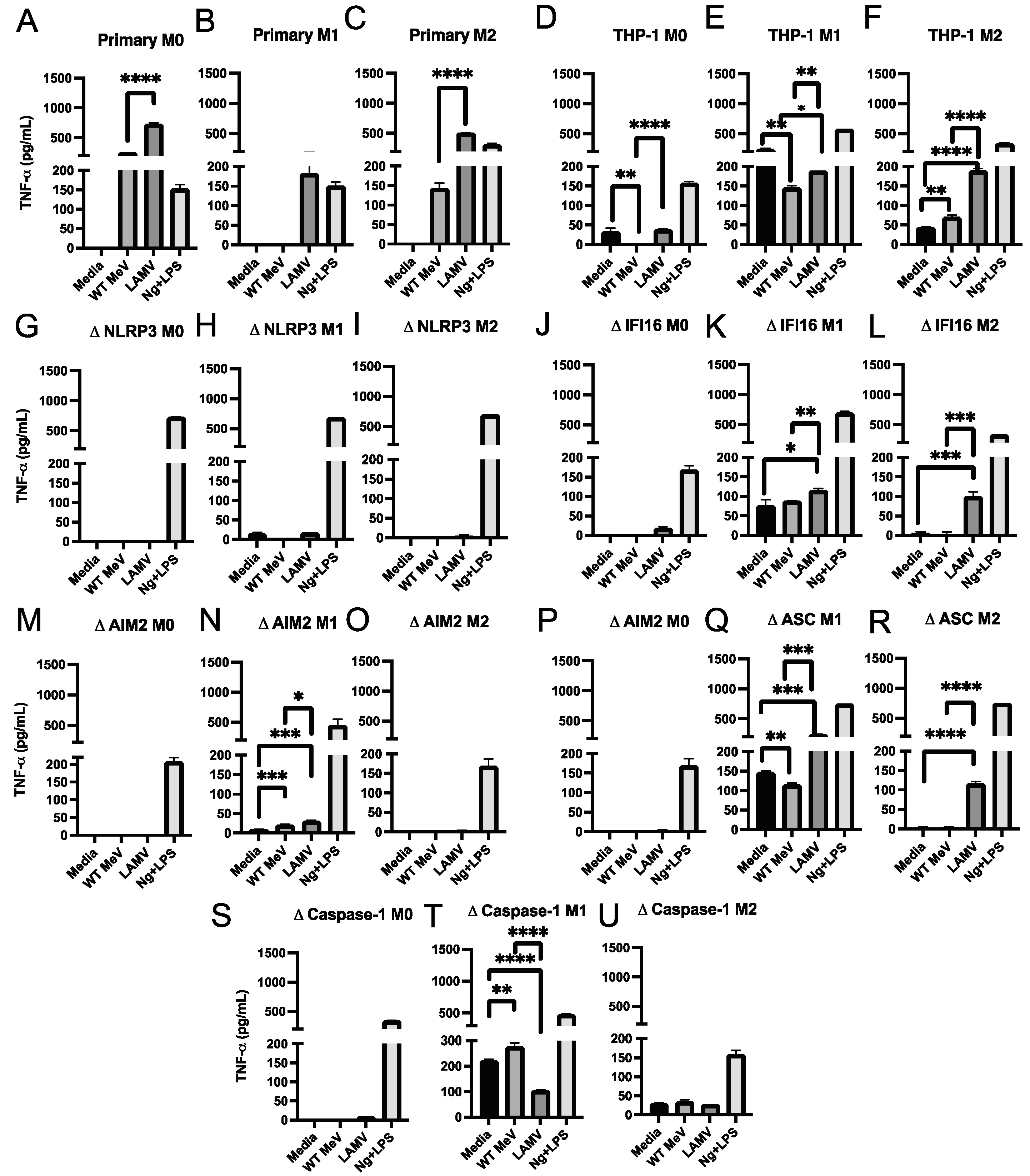
3.6. Production of IL-6 and TNFα by MeV-Infected Macrophages
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patel, M.K.; Antoni, S.; Nedelec, Y.; Sodha, S.; Menning, L.; Ogbuanu, I.U.; Gacic Dobo, M. The Changing Global Epidemiology of Measles, 2013–2018. J. Infect. Dis. 2020, 222, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Panum, P. Observations made during the epidemic of measles on the Faroe Islands in the year 1846. Med. Class. 1938, 3, 829–886. [Google Scholar]
- Mina, M.J.; Kula, T.; Leng, Y.; Li, M.; de Vries, R.D.; Knip, M.; Siljander, H.; Rewers, M.; Choy, D.F.; Wilson, M.S.; et al. Measles virus infection diminishes preexisting antibodies that offer protection from other pathogens. Science 2019, 366, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.J.; Metcalf, C.J.; de Swart, R.L.; Osterhaus, A.D.; Grenfell, B.T. Vaccines. Long-term measles-induced immunomodulation increases overall childhood infectious disease mortality. Science 2015, 348, 694–699. [Google Scholar] [CrossRef]
- Nelson, A.N.; Lin, W.W.; Shivakoti, R.; Putnam, N.E.; Mangus, L.; Adams, R.J.; Hauer, D.; Baxter, V.K.; Griffin, D.E. Association of persistent wild-type measles virus RNA with long-term humoral immunity in rhesus macaques. JCI Insight 2020, 5, e134992. [Google Scholar] [CrossRef]
- Lin, W.H.; Kouyos, R.D.; Adams, R.J.; Grenfell, B.T.; Griffin, D.E. Prolonged persistence of measles virus RNA is characteristic of primary infection dynamics. Proc. Natl. Acad. Sci. USA 2012, 109, 14989–14994. [Google Scholar] [CrossRef]
- Amanna, I.J.; Carlson, N.E.; Slifka, M.K. Duration of humoral immunity to common viral and vaccine antigens. N. Engl. J. Med. 2007, 357, 1903–1915. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef]
- Shivakoti, R.; Hauer, D.; Adams, R.J.; Lin, W.H.; Duprex, W.P.; de Swart, R.L.; Griffin, D.E. Limited in vivo production of type I or type III interferon after infection of macaques with vaccine or wild-type strains of measles virus. J. Interferon Cytokine Res. 2015, 35, 292–301. [Google Scholar] [CrossRef]
- Fontana, J.M.; Bankamp, B.; Bellini, W.J.; Rota, P.A. Regulation of interferon signaling by the C and V proteins from attenuated and wild-type strains of measles virus. Virology 2008, 374, 71–81. [Google Scholar] [CrossRef]
- Zilliox, M.J.; Moss, W.J.; Griffin, D.E. Gene expression changes in peripheral blood mononuclear cells during measles virus infection. Clin. Vaccine Immunol. 2007, 14, 918–923. [Google Scholar] [CrossRef]
- Phillips, R.S.; Enwonwu, C.O.; Okolo, S.; Hassan, A. Metabolic effects of acute measles in chronically malnourished Nigerian children. J. Nutr. Biochem. 2004, 15, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Sato, T.A.; Katayama, A.; Higuchi, K.; Shichijo, K.; Tsuchiya, T.; Takayama, N.; Takeuchi, Y.; Abe, T.; Okabe, N.; et al. Comparative analysis of host responses related to immunosuppression between measles patients and vaccine recipients with live attenuated measles vaccines. Arch. Virol. 2001, 146, 859–874. [Google Scholar] [CrossRef]
- Lin, W.W.; Nelson, A.N.; Ryon, J.J.; Moss, W.J.; Griffin, D.E. Plasma Cytokines and Chemokines in Zambian Children With Measles: Innate Responses and Association With HIV-1 Coinfection and In-Hospital Mortality. J. Infect. Dis. 2017, 215, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.W.; Moran, E.; Adams, R.J.; Sievers, R.E.; Hauer, D.; Godin, S.; Griffin, D.E. A durable protective immune response to wild-type measles virus infection of macaques is due to viral replication and spread in lymphoid tissues. Sci. Transl. Med. 2020, 12, eaax7799. [Google Scholar] [CrossRef]
- Condack, C.; Grivel, J.C.; Devaux, P.; Margolis, L.; Cattaneo, R. Measles virus vaccine attenuation: Suboptimal infection of lymphatic tissue and tropism alteration. J. Infect. Dis. 2007, 196, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Esolen, L.M.; Ward, B.J.; Moench, T.R.; Griffin, D.E. Infection of monocytes during measles. J. Infect. Dis. 1993, 168, 47–52. [Google Scholar] [CrossRef]
- Lemon, K.; de Vries, R.D.; Mesman, A.W.; McQuaid, S.; van Amerongen, G.; Yuksel, S.; Ludlow, M.; Rennick, L.J.; Kuiken, T.; Rima, B.K.; et al. Early target cells of measles virus after aerosol infection of non-human primates. PLoS Pathog. 2011, 7, e1001263. [Google Scholar] [CrossRef]
- Laksono, B.M.; Fortugno, P.; Nijmeijer, B.M.; de Vries, R.D.; Cordisco, S.; Kuiken, T.; Geijtenbeek, T.B.H.; Duprex, W.P.; Brancati, F.; de Swart, R.L. Measles skin rash: Infection of lymphoid and myeloid cells in the dermis precedes viral dissemination to the epidermis. PLoS Pathog. 2020, 16, e1008253. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.V.; McQuaid, S.; Penalva, R.; Ludlow, M.; Duprex, W.P.; Rima, B.K. Macrophages and Dendritic Cells Are the Predominant Cells Infected in Measles in Humans. mSphere 2018, 3, e00570-17. [Google Scholar] [CrossRef] [PubMed]
- Rennick, L.J.; de Vries, R.D.; Carsillo, T.J.; Lemon, K.; van Amerongen, G.; Ludlow, M.; Nguyen, D.T.; Yuksel, S.; Verburgh, R.J.; Haddock, P.; et al. Live-attenuated measles virus vaccine targets dendritic cells and macrophages in muscle of nonhuman primates. J. Virol. 2015, 89, 2192–2200. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Inflammasome activation and regulation: Toward a better understanding of complex mechanisms. Cell Discov. 2020, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Guarda, G.; Zenger, M.; Yazdi, A.S.; Schroder, K.; Ferrero, I.; Menu, P.; Tardivel, A.; Mattmann, C.; Tschopp, J. Differential expression of NLRP3 among hematopoietic cells. J. Immunol. 2011, 186, 2529–2534. [Google Scholar] [CrossRef] [PubMed]
- Kummer, J.A.; Broekhuizen, R.; Everett, H.; Agostini, L.; Kuijk, L.; Martinon, F.; van Bruggen, R.; Tschopp, J. Inflammasome components NALP 1 and 3 show distinct but separate expression profiles in human tissues suggesting a site-specific role in the inflammatory response. J. Histochem. Cytochem. 2007, 55, 443–452. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; de Sa, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Goncalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218, e20201707. [Google Scholar] [CrossRef]
- Karaba, A.H.; Figueroa, A.; Massaccesi, G.; Botto, S.; DeFilippis, V.R.; Cox, A.L. Herpes simplex virus type 1 inflammasome activation in proinflammatory human macrophages is dependent on NLRP3, ASC, and caspase-1. PLoS ONE 2020, 15, e0229570. [Google Scholar] [CrossRef]
- Shrivastava, G.; Leon-Juarez, M.; Garcia-Cordero, J.; Meza-Sanchez, D.E.; Cedillo-Barron, L. Inflammasomes and its importance in viral infections. Immunol. Res. 2016, 64, 1101–1117. [Google Scholar] [CrossRef]
- Shrivastava, G.; Valenzuela Leon, P.C.; Calvo, E. Inflammasome Fuels Dengue Severity. Front. Cell. Infect. Microbiol. 2020, 10, 489. [Google Scholar] [CrossRef]
- Chen, I.Y.; Ichinohe, T. Response of host inflammasomes to viral infection. Trends Microbiol. 2015, 23, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Chattergoon, M.A.; Latanich, R.; Quinn, J.; Winter, M.E.; Buckheit, R.W., 3rd; Blankson, J.N.; Pardoll, D.; Cox, A.L. HIV and HCV activate the inflammasome in monocytes and macrophages via endosomal Toll-like receptors without induction of type 1 interferon. PLoS Pathog. 2014, 10, e1004082. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- Botto, S.; Abraham, J.; Mizuno, N.; Pryke, K.; Gall, B.; Landais, I.; Streblow, D.N.; Fruh, K.J.; DeFilippis, V.R. Human Cytomegalovirus Immediate Early 86-kDa Protein Blocks Transcription and Induces Degradation of the Immature Interleukin-1beta Protein during Virion-Mediated Activation of the AIM2 Inflammasome. mBio 2019, 10, e02510-18. [Google Scholar] [CrossRef]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef]
- Surdziel, E.; Clay, I.; Nigsch, F.; Thiemeyer, A.; Allard, C.; Hoffman, G.; Reece-Hoyes, J.S.; Phadke, T.; Gambert, R.; Keller, C.G.; et al. Multidimensional pooled shRNA screens in human THP-1 cells identify candidate modulators of macrophage polarization. PLoS ONE 2017, 12, e0183679. [Google Scholar] [CrossRef]
- Ono, N.; Tatsuo, H.; Hidaka, Y.; Aoki, T.; Minagawa, H.; Yanagi, Y. Measles viruses on throat swabs from measles patients use signaling lymphocytic activation molecule (CDw150) but not CD46 as a cellular receptor. J. Virol. 2001, 75, 4399–4401. [Google Scholar] [CrossRef]
- Mariathasan, S.; Weiss, D.S.; Newton, K.; McBride, J.; O’Rourke, K.; Roose-Girma, M.; Lee, W.P.; Weinrauch, Y.; Monack, D.M.; Dixit, V.M. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 2006, 440, 228–232. [Google Scholar] [CrossRef]
- Joerink, M.; Rindsjo, E.; van Riel, B.; Alm, J.; Papadogiannakis, N. Placental macrophage (Hofbauer cell) polarization is independent of maternal allergen-sensitization and presence of chorioamnionitis. Placenta 2011, 32, 380–385. [Google Scholar] [CrossRef]
- Lin, W.W.; Tsay, A.J.; Lalime, E.N.; Pekosz, A.; Griffin, D.E. Primary differentiated respiratory epithelial cells respond to apical measles virus infection by shedding multinucleated giant cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2013264118. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, H.; Brankin, B.; Brady, C.; Cosby, S.L. Wild-type measles virus infection upregulates poliovirus receptor-related 4 and causes apoptosis in brain endothelial cells by induction of tumor necrosis factor-related apoptosis-inducing ligand. J. Neuropathol. Exp. Neurol. 2013, 72, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Andres, O.; Obojes, K.; Kim, K.S.; ter Meulen, V.; Schneider-Schaulies, J. CD46- and CD150-independent endothelial cell infection with wild-type measles viruses. J. Gen. Virol. 2003, 84, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.; Tanaka, Y.; Kitagawa, Y.; Koide, N.; Naiki, Y.; Morita, N.; Gotoh, B.; Yokochi, T. Sendai Virus V Protein Inhibits the Secretion of Interleukin-1beta by Preventing NLRP3 Inflammasome Assembly. J. Virol. 2018, 92, e00842-18. [Google Scholar] [CrossRef]
- Komune, N.; Ichinohe, T.; Ito, M.; Yanagi, Y. Measles virus V protein inhibits NLRP3 inflammasome-mediated interleukin-1beta secretion. J. Virol. 2011, 85, 13019–13026. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host cell death and inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- de Vries, R.D.; Lemon, K.; Ludlow, M.; McQuaid, S.; Yuksel, S.; van Amerongen, G.; Rennick, L.J.; Rima, B.K.; Osterhaus, A.D.; de Swart, R.L.; et al. In vivo tropism of attenuated and pathogenic measles virus expressing green fluorescent protein in macaques. J. Virol. 2010, 84, 4714–4724. [Google Scholar] [CrossRef]
- Takeuchi, K.; Nagata, N.; Kato, S.I.; Ami, Y.; Suzaki, Y.; Suzuki, T.; Sato, Y.; Tsunetsugu-Yokota, Y.; Mori, K.; Van Nguyen, N.; et al. Wild-type measles virus with the hemagglutinin protein of the edmonston vaccine strain retains wild-type tropism in macaques. J. Virol. 2012, 86, 3027–3037. [Google Scholar] [CrossRef]
- Okamoto, Y.; Vricella, L.A.; Moss, W.J.; Griffin, D.E. Immature CD4+CD8+ thymocytes are preferentially infected by measles virus in human thymic organ cultures. PLoS ONE 2012, 7, e45999. [Google Scholar] [CrossRef]
- Parks, C.L.; Lerch, R.A.; Walpita, P.; Wang, H.P.; Sidhu, M.S.; Udem, S.A. Comparison of predicted amino acid sequences of measles virus strains in the Edmonston vaccine lineage. J. Virol. 2001, 75, 910–920. [Google Scholar] [CrossRef]
- Bankamp, B.; Takeda, M.; Zhang, Y.; Xu, W.; Rota, P.A. Genetic characterization of measles vaccine strains. J. Infect. Dis. 2011, 204 (Suppl. 1), S533–S548. [Google Scholar] [CrossRef]
- Bankamp, B.; Hodge, G.; McChesney, M.B.; Bellini, W.J.; Rota, P.A. Genetic changes that affect the virulence of measles virus in a rhesus macaque model. Virology 2008, 373, 39–50. [Google Scholar] [CrossRef]
- Nakatsu, Y.; Takeda, M.; Iwasaki, M.; Yanagi, Y. A highly attenuated measles virus vaccine strain encodes a fully functional C protein. J. Virol. 2009, 83, 11996–12001. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Ohgimoto, S.; Sharma, L.B.; Kurazono, S.; Ayata, M.; Komase, K.; Takeda, M.; Takeuchi, K.; Ihara, T.; Ogura, H. Reduced ability of hemagglutinin of the CAM-70 measles virus vaccine strain to use receptors CD46 and SLAM. Vaccine 2009, 27, 3838–3848. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Sakaguchi, T.; Li, Y.; Kobune, F.; Kato, A.; Nagai, Y. The genome nucleotide sequence of a contemporary wild strain of measles virus and its comparison with the classical Edmonston strain genome. Virology 1999, 256, 340–350. [Google Scholar] [CrossRef]
- Takeda, M.; Ohno, S.; Tahara, M.; Takeuchi, H.; Shirogane, Y.; Ohmura, H.; Nakamura, T.; Yanagi, Y. Measles viruses possessing the polymerase protein genes of the Edmonston vaccine strain exhibit attenuated gene expression and growth in cultured cells and SLAM knock-in mice. J. Virol. 2008, 82, 11979–11984. [Google Scholar] [CrossRef]
- Takeuchi, K.; Miyajima, N.; Kobune, F.; Tashiro, M. Comparative nucleotide sequence analyses of the entire genomes of B95a cell-isolated and vero cell-isolated measles viruses from the same patient. Virus Genes 2000, 20, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Druelle, J.; Sellin, C.I.; Waku-Kouomou, D.; Horvat, B.; Wild, F.T. Wild type measles virus attenuation independent of type I IFN. Virol. J. 2008, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Hayward, J.A.; Mathur, A.; Ngo, C.; Man, S.M. Cytosolic Recognition of Microbes and Pathogens: Inflammasomes in Action. Microbiol. Mol. Biol. Rev. 2018, 82, e00015-18. [Google Scholar] [CrossRef]
- Netea, M.G.; van de Veerdonk, F.L.; van der Meer, J.W.; Dinarello, C.A.; Joosten, L.A. Inflammasome-independent regulation of IL-1-family cytokines. Annu. Rev. Immunol. 2015, 33, 49–77. [Google Scholar] [CrossRef] [PubMed]
- Matikainen, S.; Nyman, T.A.; Cypryk, W. Function and Regulation of Noncanonical Caspase-4/5/11 Inflammasome. J. Immunol. 2020, 204, 3063–3069. [Google Scholar] [CrossRef]
- Vigano, E.; Diamond, C.E.; Spreafico, R.; Balachander, A.; Sobota, R.M.; Mortellaro, A. Human caspase-4 and caspase-5 regulate the one-step non-canonical inflammasome activation in monocytes. Nat. Commun. 2015, 6, 8761. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, W.; Ma, X.; Wu, J.; Qin, X.; Cao, X.; Zhou, J.; Jin, L.; He, J.; Zheng, H.; et al. Peste Des Petits Ruminants Virus N Protein Is a Critical Proinflammation Factor That Promotes MyD88 and NLRP3 Complex Assembly. J. Virol. 2022, 96, e0030922. [Google Scholar] [CrossRef]
- Donohue, R.C.; Pfaller, C.K.; Cattaneo, R. Cyclical adaptation of measles virus quasispecies to epithelial and lymphocytic cells: To V, or not to V. PLoS Pathog. 2019, 15, e1007605. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef]
- Vecchie, A.; Bonaventura, A.; Toldo, S.; Dagna, L.; Dinarello, C.A.; Abbate, A. IL-18 and infections: Is there a role for targeted therapies? J. Cell. Physiol. 2021, 236, 1638–1657. [Google Scholar] [CrossRef]
- Nelson, A.N.; Putnam, N.; Hauer, D.; Baxter, V.K.; Adams, R.J.; Griffin, D.E. Evolution of T Cell Responses during Measles Virus Infection and RNA Clearance. Sci. Rep. 2017, 7, 11474. [Google Scholar] [CrossRef]
- Deng, J.; Yu, X.Q.; Wang, P.H. Inflammasome activation and Th17 responses. Mol. Immunol. 2019, 107, 142–164. [Google Scholar] [CrossRef]
- Ichinohe, T.; Pang, I.K.; Iwasaki, A. Influenza virus activates inflammasomes via its intracellular M2 ion channel. Nat. Immunol. 2010, 11, 404–410. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suwanmanee, S.; Ghimire, S.; Edwards, J.; Griffin, D.E. Infection of Pro- and Anti-Inflammatory Macrophages by Wild Type and Vaccine Strains of Measles Virus: NLRP3 Inflammasome Activation Independent of Virus Production. Viruses 2023, 15, 260. https://doi.org/10.3390/v15020260
Suwanmanee S, Ghimire S, Edwards J, Griffin DE. Infection of Pro- and Anti-Inflammatory Macrophages by Wild Type and Vaccine Strains of Measles Virus: NLRP3 Inflammasome Activation Independent of Virus Production. Viruses. 2023; 15(2):260. https://doi.org/10.3390/v15020260
Chicago/Turabian StyleSuwanmanee, San, Shristi Ghimire, Jerome Edwards, and Diane E. Griffin. 2023. "Infection of Pro- and Anti-Inflammatory Macrophages by Wild Type and Vaccine Strains of Measles Virus: NLRP3 Inflammasome Activation Independent of Virus Production" Viruses 15, no. 2: 260. https://doi.org/10.3390/v15020260
APA StyleSuwanmanee, S., Ghimire, S., Edwards, J., & Griffin, D. E. (2023). Infection of Pro- and Anti-Inflammatory Macrophages by Wild Type and Vaccine Strains of Measles Virus: NLRP3 Inflammasome Activation Independent of Virus Production. Viruses, 15(2), 260. https://doi.org/10.3390/v15020260