
Human Metapneumovirus-Induced Host microRNA Expression Impairs the Interferon Response in Macrophages and Epithelial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Virus Stocks
2.3. Viral Infection
2.4. Flow Cytometry
2.5. Nucleofection
2.6. RNA Extraction
2.7. Quantitative Real-Time Reverse Transcription-PCR (qRT-PCR)
2.8. Statistical Analysis
3. Results
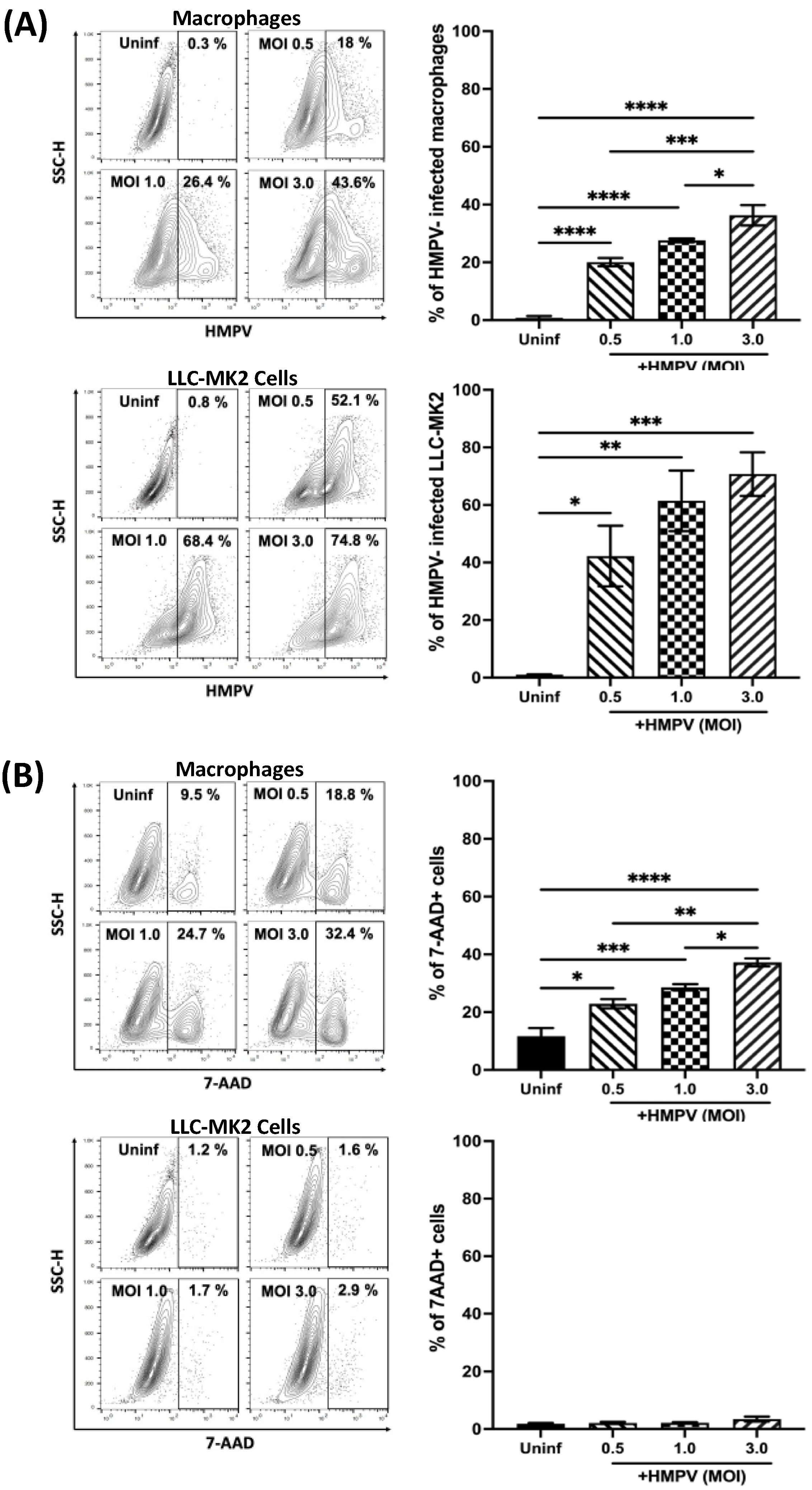
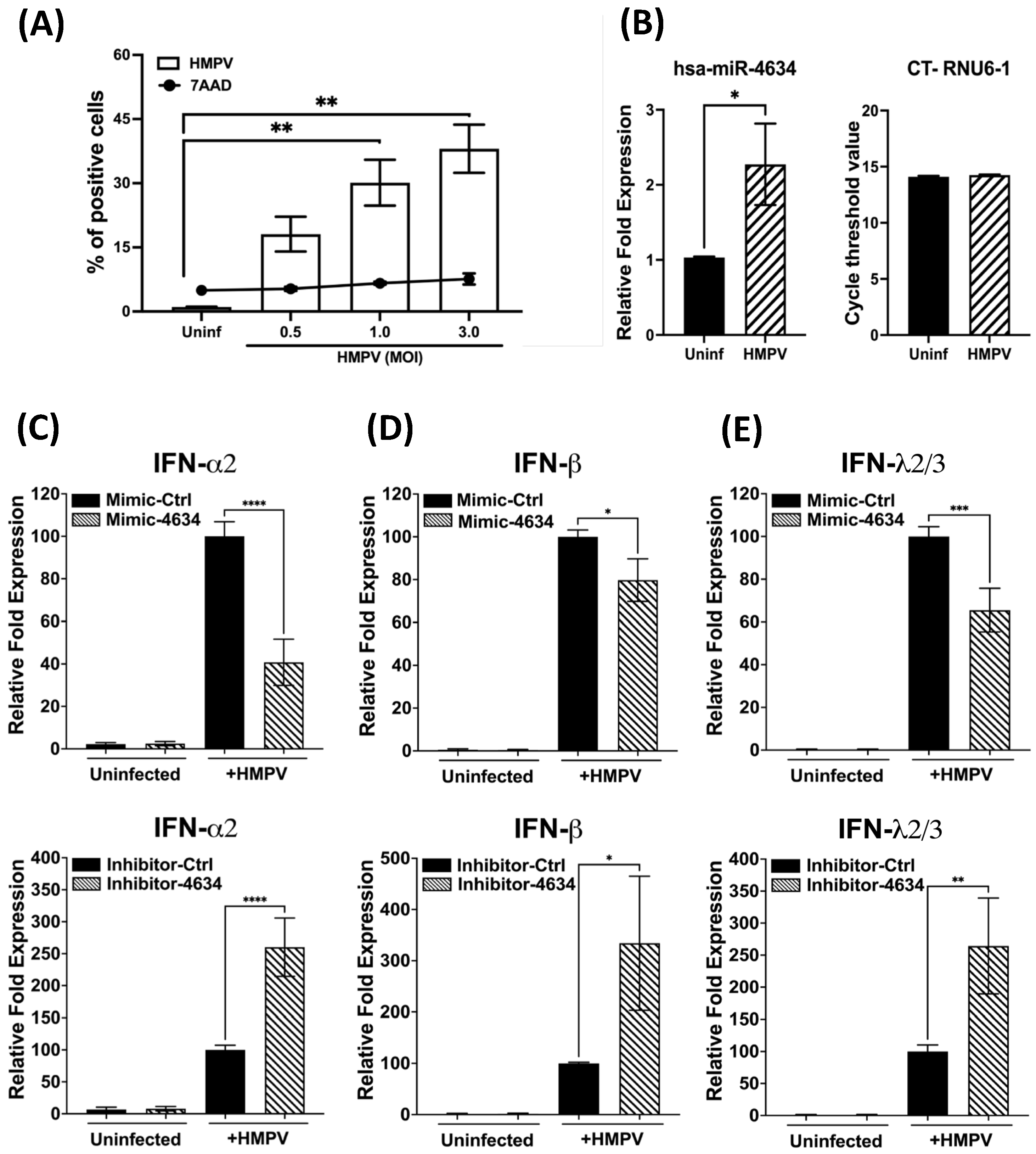
3.1. Suseptibility of Macrophages to HMPV Infection
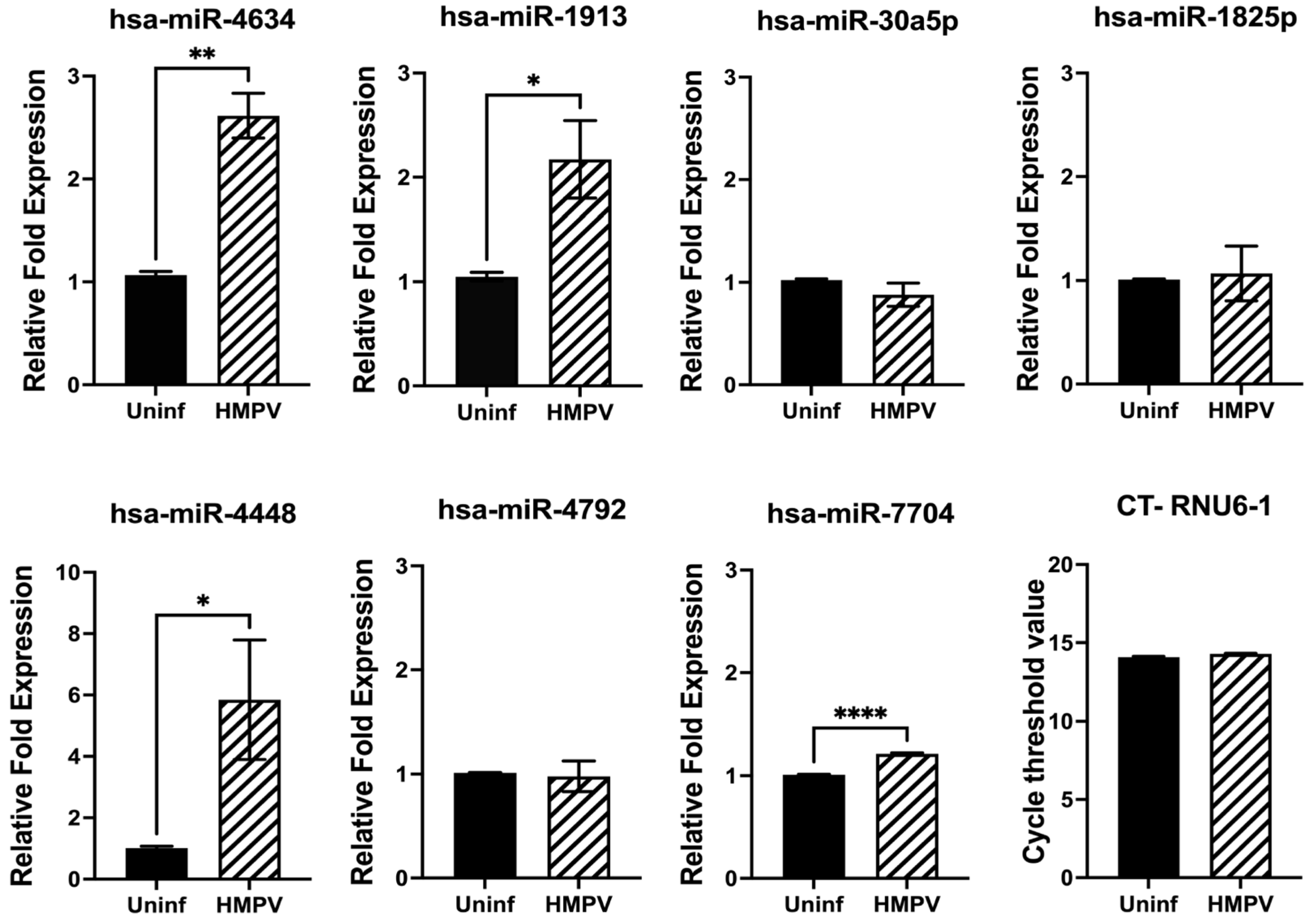
3.2. miRNA Expression in HMPV-Infected Macrophages
3.3. Functional Analysis of hsa-miR-4634 in Macrophages Infected with HMPV
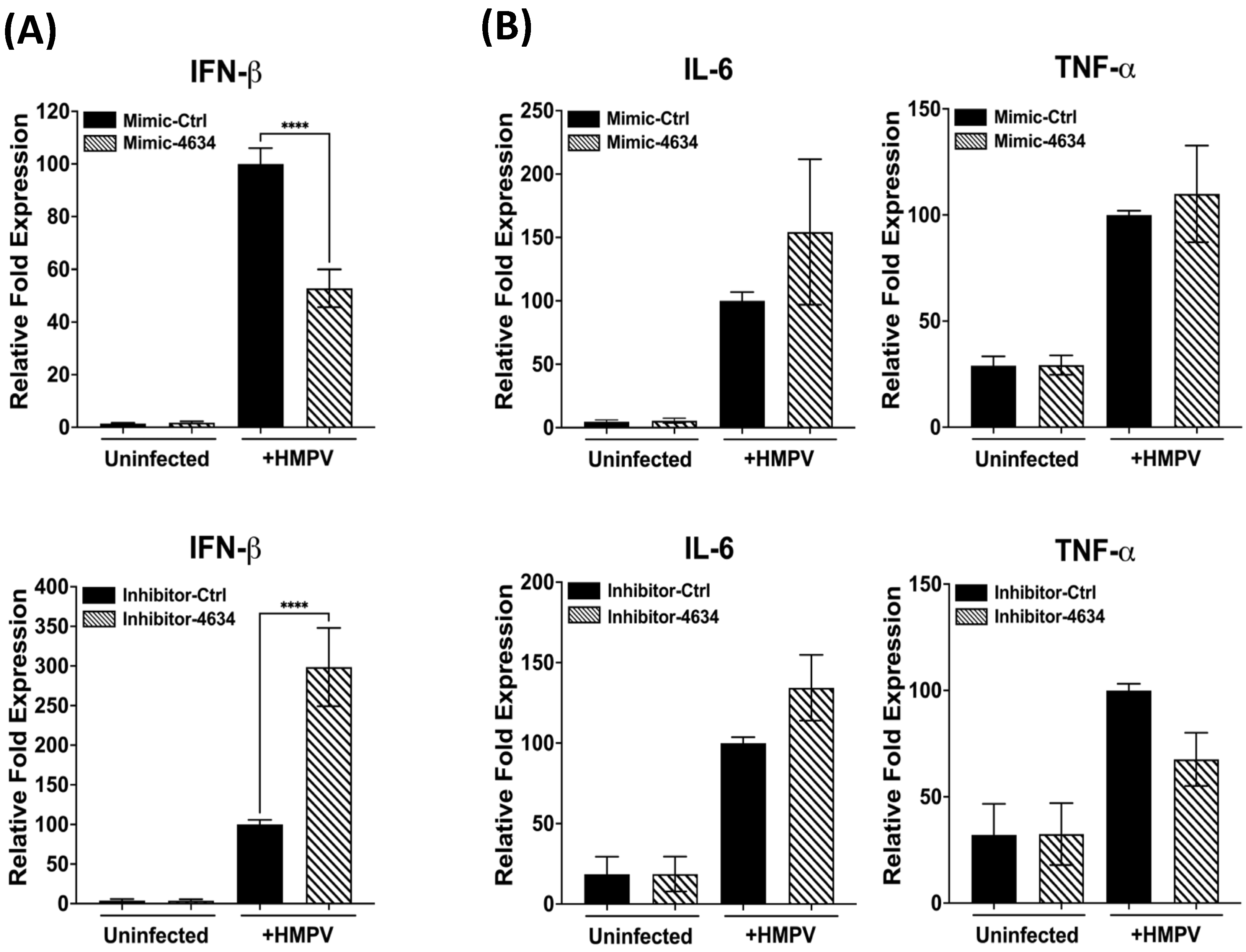
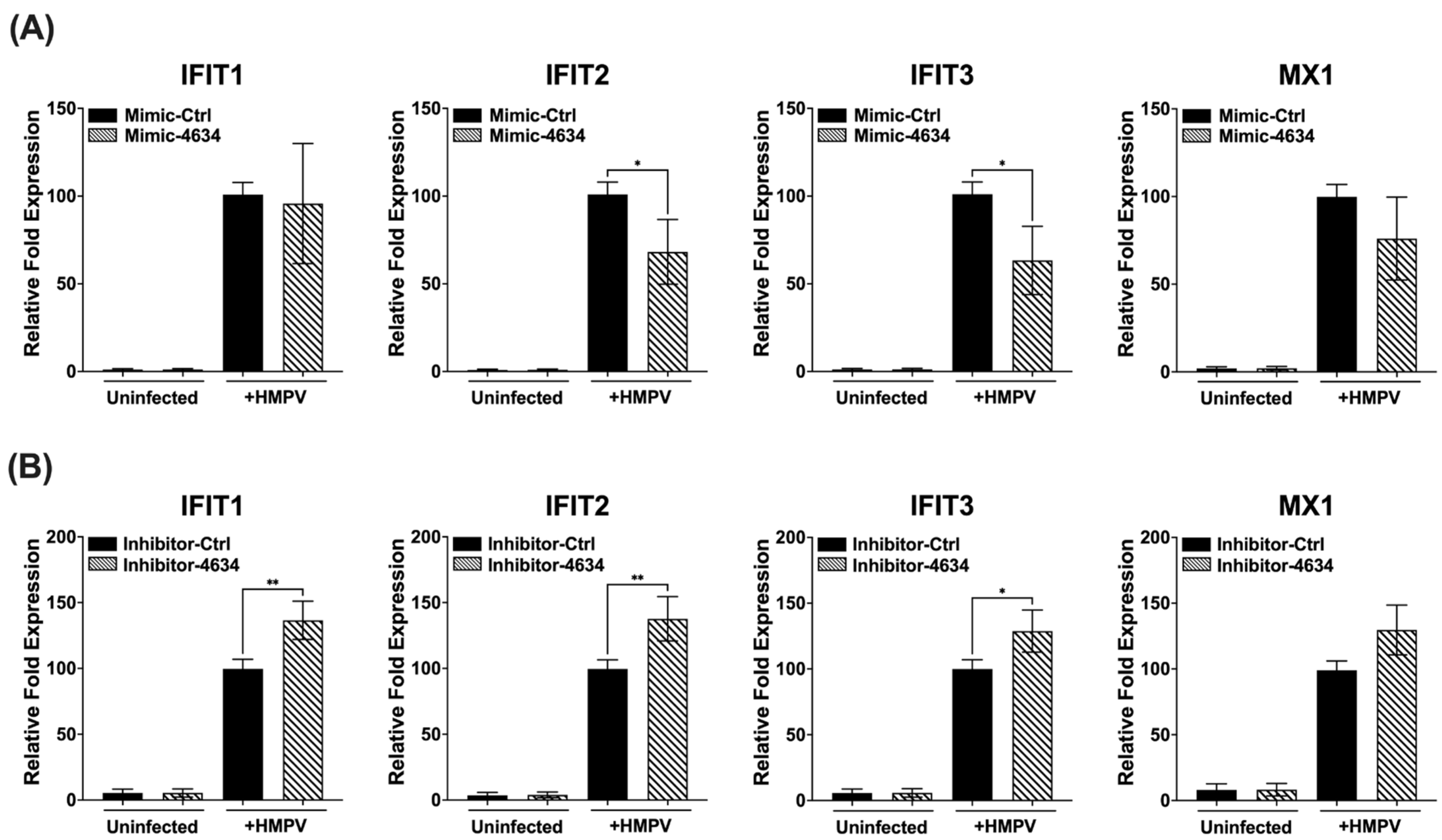
3.4. Interferon-Stimulated Gene (ISG) Expression in Macrophages Treated with Mimic and Inhibitor of hsa-miR-4634
3.5. Functional Analysis of hsa-miR-4634 in Epithelial Cells Infected with HMPV
3.6. hsa-miR-4634 Regulates the Viral Replication in Macrophages and Epithelial Cells
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gálvez, N.M.S.; Andrade, C.A.; Pacheco, G.A.; Soto, J.A.; Stranger, V.; Rivera, T.; Vásquez, A.E.; Kalergis, A.M. Host Components That Modulate the Disease Caused by hMPV. Viruses 2021, 13, 519. [Google Scholar] [CrossRef] [PubMed]
- Rima, B.; Collins, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.A.; Lee, B.; Maisner, A.; Rota, P.; Wang, L.; et al. ICTV Virus Taxonomy Profile: Pneumoviridae. J. Gen. Virol. 2017, 98, 2912–2913. [Google Scholar] [CrossRef] [PubMed]
- Kahn, J.S. Epidemiology of Human Metapneumovirus. Clin. Microbiol. Rev. 2006, 19, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Hasvold, J.; Sjoding, M.; Pohl, K.; Cooke, C.R.; Hyzy, R.C. The role of human metapneumovirus in the critically ill adult patient. J. Crit. Care 2016, 31, 233–237. [Google Scholar] [CrossRef]
- Van Den Hoogen, B.G.; De Jong, J.C.; Groen, J.; Kuiken, T.; De Groot, R.; Fouchier, R.A.M.; Osterhaus, A.D.M.E. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat. Med. 2001, 7, 719–724. [Google Scholar] [CrossRef]
- Divarathna, M.V.M.; Rafeek, R.A.M.; Noordeen, F. A review on epidemiology and impact of human metapneumovirus infections in children using TIAB search strategy on PubMed and PubMed Central articles. Rev. Med. Virol. 2020, 30, e2090. [Google Scholar] [CrossRef]
- Edwards, K.M.; Zhu, Y.; Griffin, M.R.; Weinberg, G.A.; Hall, C.B.; Szilagyi, P.G.; Staat, M.A.; Iwane, M.; Prill, M.M.; Williams, J.V. Burden of Human Metapneumovirus Infection in Young Children. N. Engl. J. Med. 2013, 368, 633–643. [Google Scholar] [CrossRef]
- Kuiken, T.; Van Den Hoogen, B.G.; Van Riel, D.A.J.; Laman, J.D.; Van Amerongen, G.; Sprong, L.; Fouchier, R.A.M.; Osterhaus, A.D.M.E. Experimental Human Metapneumovirus Infection of Cynomolgus Macaques (Macaca fascicularis) Results in Virus Replication in Ciliated Epithelial Cells and Pneumocytes with Associated Lesions throughout the Respiratory Tract. Am. J. Pathol. 2004, 164, 1893–1900. [Google Scholar] [CrossRef]
- Loevenich, S.; Spahn, A.S.; Rian, K.; Boyartchuk, V.; Anthonsen, M.W. Human Metapneumovirus Induces IRF1 via TANK-Binding Kinase 1 and Type I IFN. Front. Immunol. 2021, 12, 563336. [Google Scholar] [CrossRef]
- Keppler, S.J.; Rosenits, K.; Koegl, T.; Vucikuja, S.; Aichele, P. Signal 3 cytokines as modulators of primary immune responses during infections: The interplay of type I IFN and IL-12 in CD8 T cell responses. PLoS ONE 2012, 7, e40865. [Google Scholar] [CrossRef]
- Hou, F.; Xiao, K.; Tang, L.; Xie, L. Diversity of Macrophages in Lung Homeostasis and Diseases. Front. Immunol. 2021, 12, 753940. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.D.; Jeong, D.; Chung, D.H. Development and Functions of Alveolar Macrophages. Mol. Cells 2021, 44, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Liu, T.; Spetch, L.; Kolli, D.; Garofalo, R.P.; Casola, A. Airway epithelial cell response to human metapneumovirus infection. Virology 2007, 368, 91–101. [Google Scholar] [CrossRef]
- Guerrero-Plata, A.; Casola, A.; Garofalo, R.P. Human metapneumovirus induces a profile of lung cytokines distinct from that of respiratory syncytial virus. J. Virol. 2005, 79, 14992–14997. [Google Scholar] [CrossRef] [PubMed]
- Malmo, J.; Moe, N.; Krokstad, S.; Ryan, L.; Loevenich, S.; Johnsen, I.B.; Espevik, T.; Nordbø, S.A.; Døllner, H.; Anthonsen, M.W. Cytokine Profiles in Human Metapneumovirus Infected Children: Identification of Genes Involved in the Antiviral Response and Pathogenesis. PLoS ONE 2016, 11, e0155484. [Google Scholar] [CrossRef] [PubMed]
- Banete, A.; Barilo, J.; Whittaker, R.; Basta, S. The Activated Macrophage—A Tough Fortress for Virus Invasion: How Viruses Strike Back. Front. Microbiol. 2021, 12, 803427. [Google Scholar] [CrossRef]
- Schneider, C.; Nobs, S.P.; Heer, A.K.; Kurrer, M.; Klinke, G.; Van Rooijen, N.; Vogel, J.; Kopf, M. Alveolar Macrophages Are Essential for Protection from Respiratory Failure and Associated Morbidity following Influenza Virus Infection. PLoS Pathog. 2014, 10, e1004053. [Google Scholar] [CrossRef]
- Somerville, L.; Cardani, A.; Braciale, T.J. Alveolar Macrophages in Influenza A Infection Guarding the Castle with Sleeping Dragons. Infect. Dis. Ther. 2020, 1–3. [Google Scholar] [CrossRef]
- Banos-Lara Mdel, R.; Harvey, L.; Mendoza, A.; Simms, D.; Chouljenko, V.N.; Wakamatsu, N.; Kousoulas, K.G.; Guerrero-Plata, A. Impact and regulation of lambda interferon response in human metapneumovirus infection. J. Virol. 2015, 89, 730–742. [Google Scholar] [CrossRef]
- Kolli, D.; Gupta, M.R.; Sbrana, E.; Velayutham, T.S.; Chao, H.; Casola, A.; Garofalo, R.P. Alveolar Macrophages Contribute to the Pathogenesis of Human Metapneumovirus Infection while Protecting against Respiratory Syncytial Virus Infection. Am. J. Respir. Cell Mol. Biol. 2014, 51, 502–515. [Google Scholar] [CrossRef]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. Methods Mol. Biol. 2017, 1509, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Bulati, M.; Miceli, V.; Amodio, N.; Conaldi, P.G. Non-Coding RNAs: Strategy for Viruses’ Offensive. Non-Coding RNA 2020, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Baños-Lara, M.D.R.; Zabaleta, J.; Garai, J.; Baddoo, M.; Guerrero-Plata, A. Comparative analysis of miRNA profile in human dendritic cells infected with respiratory syncytial virus and human metapneumovirus. BMC Res. Notes 2018, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Espinoza, I.; Banos-Lara, M.D.R.; Guerrero-Plata, A. The Importance of miRNA Identification During Respiratory Viral Infections. J. Cell Immunol. 2021, 3, 207–214. [Google Scholar] [CrossRef]
- Biacchesi, S.; Skiadopoulos, M.H.; Tran, K.C.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Recovery of human metapneumovirus from cDNA: Optimization of growth in vitro and expression of additional genes. Virology 2004, 321, 247–259. [Google Scholar] [CrossRef]
- Guerrero-Plata, A.; Baron, S.; Poast, J.S.; Adegboyega, P.A.; Casola, A.; Garofalo, R.P. Activity and Regulation of Alpha Interferon in Respiratory Syncytial Virus and Human Metapneumovirus Experimental Infections. J. Virol. 2005, 79, 10190–10199. [Google Scholar] [CrossRef]
- Guerrero-Plata, A.; Casola, A.; Suarez, G.; Yu, X.; Spetch, L.; Peeples, M.E.; Garofalo, R.P. Differential Response of Dendritic Cells to Human Metapneumovirus and Respiratory Syncytial Virus. Am. J. Respir. Cell Mol. Biol. 2006, 34, 320–329. [Google Scholar] [CrossRef]
- Deffrasnes, C.; Côté, S.; Boivin, G. Analysis of replication kinetics of the human metapneumovirus in different cell lines by real-time PCR. J. Clin. Microbiol. 2005, 43, 488–490. [Google Scholar] [CrossRef]
- Martínez-Espinoza, I. THP-1-derevided macrophages infected with HMPV at MOIs of 5.0 and 10.0 induced 42% and 43.4% cell mortality, respectively. 2023. Unpublished work. [Google Scholar]
- Martínez-Espinoza, I. Interfeon alpha-2 was not detected in THP-1-derived macrophages infected with HMPV. 2023. Unpublished work. [Google Scholar]
- Mcnab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Klimyte, E.M.; Smith, S.E.; Oreste, P.; Lembo, D.; Dutch, R.E. Inhibition of Human Metapneumovirus Binding to Heparan Sulfate Blocks Infection in Human Lung Cells and Airway Tissues. J. Virol. 2016, 90, 9237–9250. [Google Scholar] [CrossRef]
- Preston, F.M.; Straub, C.P.; Ramirez, R.; Mahalingam, S.; Spann, K.M. siRNA against the G gene of human metapneumovirus. Virol. J. 2012, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Liu, T.; Shan, Y.; Li, K.; Garofalo, R.P.; Casola, A. Human Metapneumovirus Glycoprotein G Inhibits Innate Immune Responses. PLoS Pathog. 2008, 4, e1000077. [Google Scholar] [CrossRef]
- Dinwiddie, D.L.; Harrod, K.S. Human metapneumovirus inhibits IFN-alpha signaling through inhibition of STAT1 phosphorylation. Am. J. Respir. Cell Mol. Biol. 2008, 38, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Soto, J.A.; Gálvez, N.M.S.; Benavente, F.M.; Pizarro-Ortega, M.S.; Lay, M.K.; Riedel, C.; Bueno, S.M.; Gonzalez, P.A.; Kalergis, A.M. Human Metapneumovirus: Mechanisms and Molecular Targets Used by the Virus to Avoid the Immune System. Front. Immunol. 2018, 9, 2466. [Google Scholar] [CrossRef]
- Loevenich, S.; Malmo, J.; Liberg, A.M.; Sherstova, T.; Li, Y.; Rian, K.; Johnsen, I.B.; Anthonsen, M.W. Cell-Type-Specific Transcription of Innate Immune Regulators in response to HMPV Infection. Mediat. Inflamm. 2019, 2019, 4964239. [Google Scholar] [CrossRef] [PubMed]
- McMichael, T.M.; Zhang, Y.; Kenney, A.D.; Zhang, L.; Zani, A.; Lu, M.; Chemudupati, M.; Li, J.; Yount, J.S. IFITM3 Restricts Human Metapneumovirus Infection. J. Infect. Dis. 2018, 218, 1582–1591. [Google Scholar] [CrossRef]
- Nao, N.; Sato, K.; Yamagishi, J.; Tahara, M.; Nakatsu, Y.; Seki, F.; Katoh, H.; Ohnuma, A.; Shirogane, Y.; Hayashi, M.; et al. Consensus and variations in cell line specificity among human metapneumovirus strains. PLoS ONE 2019, 14, e0215822. [Google Scholar] [CrossRef]
- Kumagai, Y.; Takeuchi, O.; Kato, H.; Kumar, H.; Matsui, K.; Morii, E.; Aozasa, K.; Kawai, T.; Akira, S. Alveolar Macrophages Are the Primary Interferon-α Producer in Pulmonary Infection with RNA Viruses. Immunity 2007, 27, 240–252. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. miRTarBase update 2022: An informative resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2022, 50, D222–D230. [Google Scholar] [CrossRef]
- Shimomura, A.; Shiino, S.; Kawauchi, J.; Takizawa, S.; Sakamoto, H.; Matsuzaki, J.; Ono, M.; Takeshita, F.; Niida, S.; Shimizu, C.; et al. Novel combination of serum microRNA for detecting breast cancer in the early stage. Cancer Sci. 2016, 107, 326–334. [Google Scholar] [CrossRef]
- Liu, S.; Zang, H.; Zheng, H.; Wang, W.; Wen, Q.; Zhan, Y.; Yang, Y.; Ning, Y.; Wang, H.; Fan, S. miR-4634 augments the anti-tumor effects of RAD001 and associates well with clinical prognosis of non-small cell lung cancer. Sci. Rep. 2020, 10, 13079. [Google Scholar] [CrossRef] [PubMed]
- Kaba, M.; Pirinççi, N.; Demir, M.; Kaba, S.; Oztuzcu, S.; Verep, S. The relationship between microRNAs and bladder cancer: Are microRNAs useful to predict bladder cancer in suspicious patients? Int. Urol. Nephrol. 2023, 55, 2483–2491. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Guo, R.; Zhang, L. Downregulation of lncRNA FOXD2-AS1 Confers Radiosensitivity to Gastric Cancer Cells via miR-1913/SETD1A Axis. Cytogenet. Genome Res. 2022, 162, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, L.; Yang, P.; Lu, Y.; Lin, S.; Yuan, G. The exosomal transfer of human bone marrow mesenchymal stem cell-derived miR-1913 inhibits osteosarcoma progression by targeting NRSN2. Am. J. Transl. Res. 2021, 13, 10178–10192. [Google Scholar] [PubMed]
- Byun, Y.J.; Piao, X.M.; Jeong, P.; Kang, H.W.; Seo, S.P.; Moon, S.K.; Lee, J.Y.; Choi, Y.H.; Lee, H.Y.; Kim, W.T.; et al. Urinary microRNA-1913 to microRNA-3659 expression ratio as a non-invasive diagnostic biomarker for prostate cancer. Investig. Clin. Urol. 2021, 62, 340–348. [Google Scholar] [CrossRef]
- Zhang, C.; Dang, D.; Liu, C.; Wang, Y.; Cong, X. Identification of tumor mutation burden-related hub genes and the underlying mechanism in melanoma. J. Cancer 2021, 12, 2440–2449. [Google Scholar] [CrossRef]
- Bai, M.; Che, Y.; Lu, K.; Fu, L. Analysis of deubiquitinase OTUD5 as a biomarker and therapeutic target for cervical cancer by bioinformatic analysis. PeerJ 2020, 8, e9146. [Google Scholar] [CrossRef]
- Stverakova, T.; Baranova, I.; Mikyskova, P.; Gajdosova, B.; Vosmikova, H.; Laco, J.; Palicka, V.; Parova, H. Selection of endogenous control and identification of significant microRNA deregulations in cervical cancer. Front. Oncol. 2023, 13, 1143691. [Google Scholar] [CrossRef]
- Zhang, D.; Yang, Y.; Kang, Y.; Xie, D.; Zhang, X.; Hao, J. Dysregulated expression of microRNA involved in resistance to osimertinib in EGFR mutant non-small cell lung cancer cells. J. Thorac. Dis. 2023, 15, 1978–1993. [Google Scholar] [CrossRef]
- Suzuki, K.; Yokoi, A.; Yoshida, K.; Kato, T.; Ochiya, T.; Yamamoto, Y.; Kajiyama, H. Preoperative serum microRNAs as potential prognostic biomarkers in ovarian clear cell carcinoma. J. Gynecol. Oncol. 2023, 34, e34. [Google Scholar] [CrossRef]
- Garcia-Marcos, M.; Ghosh, P.; Farquhar, M.G. GIV/Girdin transmits signals from multiple receptors by triggering trimeric G protein activation. J. Biol. Chem. 2015, 290, 6697–6704. [Google Scholar] [CrossRef] [PubMed]
- Mittal, Y.; Pavlova, Y.; Garcia-Marcos, M.; Ghosh, P. Src homology domain 2-containing protein-tyrosine phosphatase-1 (SHP-1) binds and dephosphorylates G(alpha)-interacting, vesicle-associated protein (GIV)/Girdin and attenuates the GIV-phosphatidylinositol 3-kinase (PI3K)-Akt signaling pathway. J. Biol. Chem. 2011, 286, 32404–32415. [Google Scholar] [CrossRef] [PubMed]
- Swanson, L.; Katkar, G.D.; Tam, J.; Pranadinata, R.F.; Chareddy, Y.; Coates, J.; Anandachar, M.S.; Castillo, V.; Olson, J.; Nizet, V.; et al. TLR4 signaling and macrophage inflammatory responses are dampened by GIV/Girdin. Proc. Natl. Acad. Sci. USA 2020, 117, 26895–26906. [Google Scholar] [CrossRef]
- Jarret, A.; McFarland, A.P.; Horner, S.M.; Kell, A.; Schwerk, J.; Hong, M.; Badil, S.; Joslyn, R.C.; Baker, D.P.; Carrington, M.; et al. Hepatitis-C-virus-induced microRNAs dampen interferon-mediated antiviral signaling. Nat. Med. 2016, 22, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.Q.; Li, L.; Wang, B.H.; Ali, A.F.; Li, W. Profiles and predictive value of cytokines in children with human metapneumovirus pneumonia. Virol. J. 2022, 19, 214. [Google Scholar] [CrossRef]
- Deng, J.; Ptashkin, R.N.; Wang, Q.; Liu, G.; Zhang, G.; Lee, I.; Lee, Y.S.; Bao, X. Human metapneumovirus infection induces significant changes in small noncoding RNA expression in airway epithelial cells. Mol. Ther. Nucleic Acids 2014, 3, e163. [Google Scholar] [CrossRef] [PubMed]
- Koltsida, O.; Hausding, M.; Stavropoulos, A.; Koch, S.; Tzelepis, G.; Ubel, C.; Kotenko, S.V.; Sideras, P.; Lehr, H.A.; Tepe, M.; et al. IL-28A (IFN-λ2) modulates lung DC function to promote Th1 immune skewing and suppress allergic airway disease. EMBO Mol. Med. 2011, 3, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Miah, S.M.; Wilson, Z.S.; Erick, T.K.; Banh, C.; Brossay, L. Role of type I interferon receptor signaling on NK cell development and functions. PLoS ONE 2014, 9, e111302. [Google Scholar] [CrossRef]
- Seo, Y.J.; Hahm, B. Type I interferon modulates the battle of host immune system against viruses. Adv. Appl. Microbiol. 2010, 73, 83–101. [Google Scholar] [CrossRef]
- Kaplan, A.; Lee, M.W.; Wolf, A.J.; Limon, J.J.; Becker, C.A.; Ding, M.; Murali, R.; Lee, E.Y.; Liu, G.Y.; Wong, G.C.L.; et al. Direct Antimicrobial Activity of IFN-β. J. Immunol. 2017, 198, 4036–4045. [Google Scholar] [CrossRef]
- Chrysanthopoulou, A.; Kambas, K.; Stakos, D.; Mitroulis, I.; Mitsios, A.; Vidali, V.; Angelidou, I.; Bochenek, M.; Arelaki, S.; Arampatzioglou, A.; et al. Interferon lambda1/IL-29 and inorganic polyphosphate are novel regulators of neutrophil-driven thromboinflammation. J. Pathol. 2017, 243, 111–122. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Espinoza, I.; Bungwon, A.D.; Guerrero-Plata, A. Human Metapneumovirus-Induced Host microRNA Expression Impairs the Interferon Response in Macrophages and Epithelial Cells. Viruses 2023, 15, 2272. https://doi.org/10.3390/v15112272
Martínez-Espinoza I, Bungwon AD, Guerrero-Plata A. Human Metapneumovirus-Induced Host microRNA Expression Impairs the Interferon Response in Macrophages and Epithelial Cells. Viruses. 2023; 15(11):2272. https://doi.org/10.3390/v15112272
Chicago/Turabian StyleMartínez-Espinoza, Iván, Anang D. Bungwon, and Antonieta Guerrero-Plata. 2023. "Human Metapneumovirus-Induced Host microRNA Expression Impairs the Interferon Response in Macrophages and Epithelial Cells" Viruses 15, no. 11: 2272. https://doi.org/10.3390/v15112272
APA StyleMartínez-Espinoza, I., Bungwon, A. D., & Guerrero-Plata, A. (2023). Human Metapneumovirus-Induced Host microRNA Expression Impairs the Interferon Response in Macrophages and Epithelial Cells. Viruses, 15(11), 2272. https://doi.org/10.3390/v15112272