

Assessment of Equine Influenza Virus Status in the Republic of Korea from 2020 to 2022
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. RNA Extraction and RT-PCR
2.3. Hemagglutination Inhibition (HI) Assay
2.4. Data Analysis
3. Results
3.1. PCR Detection of Equine Influenza Virus
3.2. Detection of EIV Antibody
3.2.1. Seropositivity Rates and Mean Antibody Titers According to Year
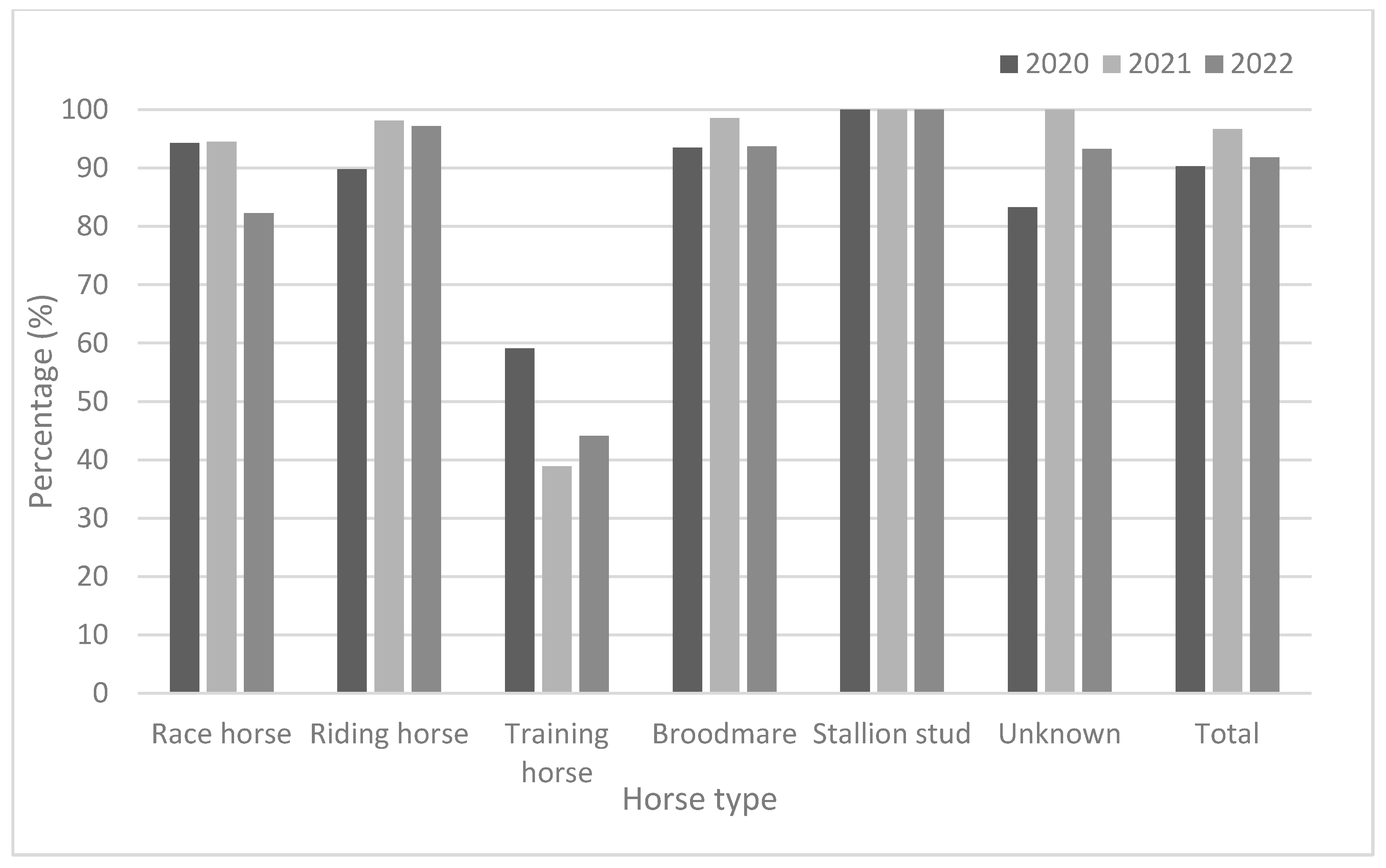
3.2.2. Seropositivity Rates and Mean Antibody Titers According to Horse Type
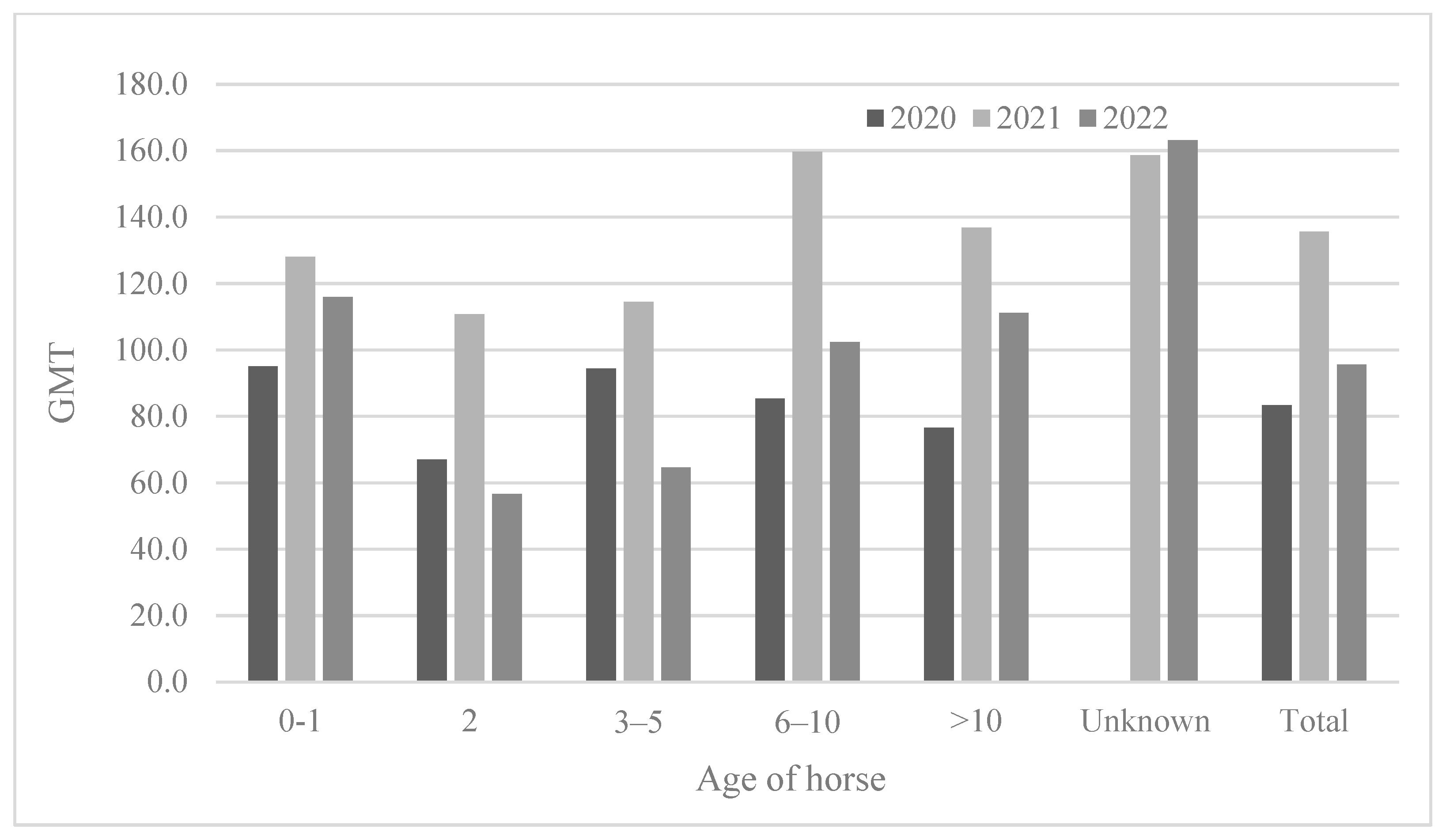
3.2.3. Seropositivity Rates and Mean Antibody Titers According to Horse Age
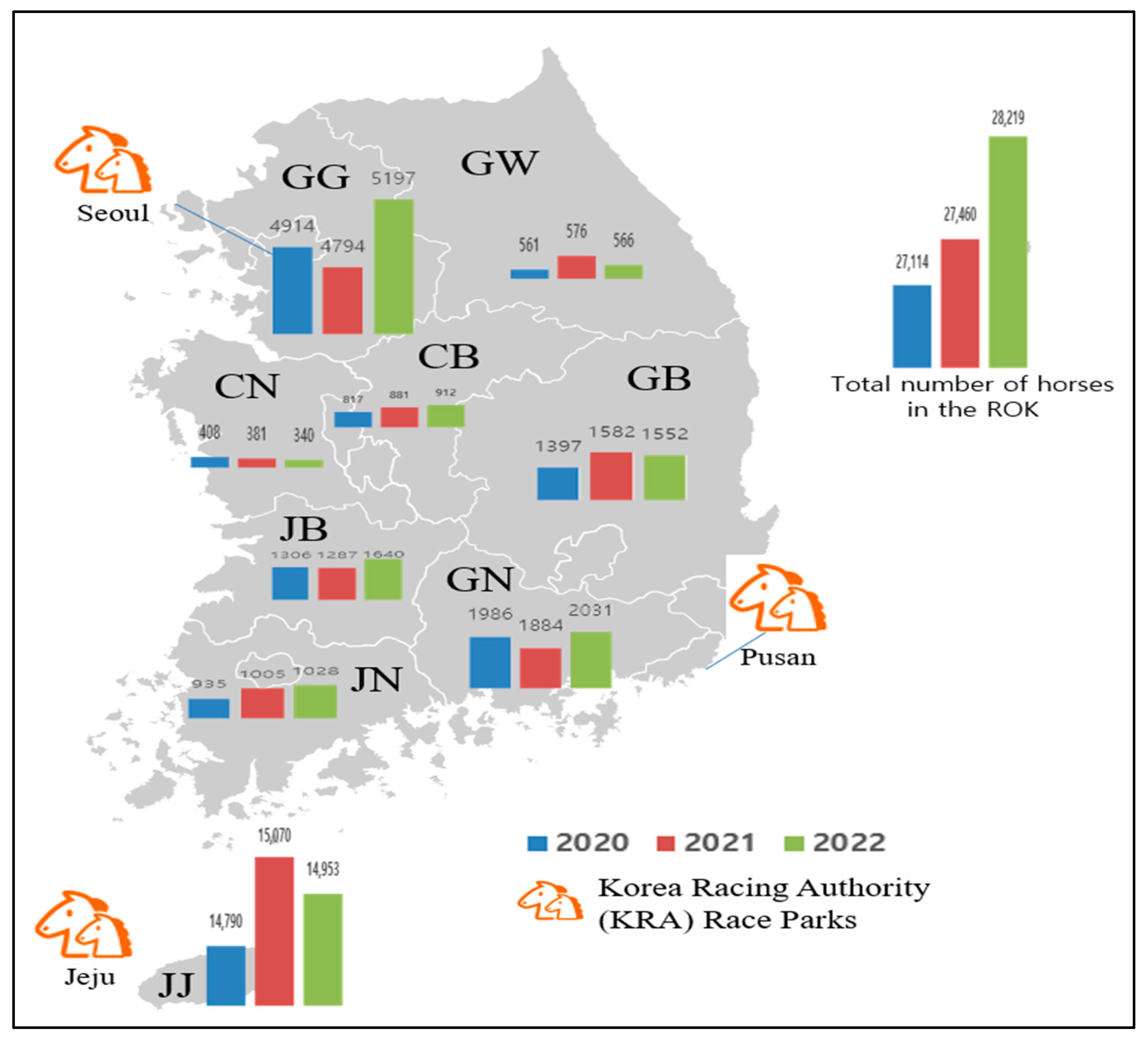
3.2.4. Seropositivity Rates and Mean Antibody Titers According to Province
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hemida, M.G.; Perera, R.A.P.; Chu, D.K.W.; Alnaeem, A.A.; Peiris, M. Evidence of equine influenza A (H3N8) activity in horses from Eastern and Central Saudi Arabia: 2013–2015. Equine Vet. J. 2019, 51, 218–221. [Google Scholar] [CrossRef]
- Whitlock, F.; Murcia, P.R.; Newton, J.R. A review on equine influenza from a human influenza perspective. Viruses 2022, 14, 1312. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Kass, P.H.; Mapes, S.; Wademan, C.; Akana, N.; Barnett, C.; MacKenzie, C.; Vaala, W. Voluntary surveillance program for equine influenza virus in the United States from 2010 to 2013. J. Vet. Intern. Med. 2015, 29, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.M. Equine influenza. Cold Spring Harb. Perspect. Med. 2022, 4, a038331. [Google Scholar] [CrossRef]
- Sovinova, O.; Tumova, B.; Pouska, F.; Nemec, J. Isolation of a virus causing respiratory disease in horses. Acta Virol. 1958, 2, 52–61. [Google Scholar]
- Paillot, R.; Hannant, D.; Kydd, J.H.; Daly, J.M. Vaccination against equine influenza: Quid novi? Vaccine 2006, 24, 4047–4061. [Google Scholar] [CrossRef]
- Oladunni, F.S.; Oseni, S.O.; Martinez-Sobrido, L.; Chambers, T.M. Equine influenza virus and vaccines. Viruses 2021, 13, 1657. [Google Scholar] [CrossRef]
- Waddell, G.H.; Teigland, M.B.; Sigel, M.M. A new influenza virus associated with equine respiratory disease. J. Am. Vet. Med. Assoc. 1963, 143, 587–590. [Google Scholar] [PubMed]
- Kim, E.J.; Kim, B.H.; Yang, S.; Choi, E.J.; Shin, Y.J.; Song, J.Y.; Shin, Y.K. Antibody responses after vaccination against influenza in the Republic of Korea in 2013. J. Vet. Med. Sci. 2015, 77, 1517–1521. [Google Scholar] [CrossRef][Green Version]
- Mino, S.; Mojsiejczuk, L.; Guo, W.; Zhang, H.; Qi, T.; Du, C.; Zhang, X.; Wang, J.; Campos, R.; Wang, X. Equine influenza virus in Asia: Phlogeographic pattern and molecular features reveal circulation of an autochthonous lineage. J. Virol. 2019, 93, e00116-19. [Google Scholar] [CrossRef]
- Na, W.; Yeom, M.; Yuk, H.; Moon, H.; Kang, B.K.; Song, D. Influenza virus vaccine for neglected hosts: Horses and dogs. Clin. Exp. Vaccine Res. 2016, 5, 117–124. [Google Scholar] [CrossRef] [PubMed]
- WOAH. WOAH Expert Surveillance Panel on Equine Influenza Vaccine Composition. 2022. Available online: http://www.woah.int/ourscientific-expertise/specific-information-and44recommendations/equine-influenza/ (accessed on 10 October 2020).
- Cullinane, A.; Elton, D.; Mumford, J. Equine influenza surveillance and control. Influenza Other Respir. Viruses 2010, 4, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Lee, C.H.; Song, J.Y.; Shin, Y.K.; Moon, J.S.; Choi, Y.G.; Kim, H.P. Serological survey on equine influenza viruses in Korea. Korean J. Vet. Med. 2010, 34, 45–51. [Google Scholar]
- Cho, M.S.; Lee, J.Y.; Lee, S.K.; Song, J.Y.; Lee, J.; Hyun, B.H.; Cho, S.D.; Ouh, I.O. Antibody responses after vaccination against equine influenza in Korea in 2016–2018. Korean J. Vet. Res. 2019, 59, 151–155. [Google Scholar] [CrossRef]
- Ouh, I.O.; Kim, M.J.; Choi, H.; Yang, H.S.; Lee, S.K.; Moon, S.Y.; Yun, S.; Hyun, B.H.; Cho, S.D.; Lee, Y.H. Antibody responses after vaccination against equine influenza virus in the Republic of Korea in 2019. J. Prev. Vet. Med. 2020, 44, 132–135. [Google Scholar] [CrossRef]
- Na, W.; Kang, B.; Kim, H.I.; Hong, M.; Park, S.J.; Jeoung, H.Y.; An, D.J.; Moon, H.; Kim, J.K.; Song, D. Isolation and genetic characterization of naturally NS-truncated H3N8 equine influenza virus in South Korea. Epidemiol. Infect. 2014, 142, 759–766. [Google Scholar] [CrossRef]
- Statistics Korea. Korean Statistical Information Service. Available online: http://kosis.kr (accessed on 26 July 2023).
- Ministry of Agriculture, Food and Rural Affairs. Livestock Product Safety Management System. Available online: http://www.lpsms.go.kr (accessed on 26 July 2023).
- Lee, E.; Kim, E.J.; Kim, B.H.; Song, J.Y.; Cho, I.S.; Shin, Y.K. Multiplex RT-PCR detection of H3N2 influenza A virus in dogs. Mol. Cell. Probes 2016, 30, 56–60. [Google Scholar] [CrossRef]
- Shigh, R.K.; Dhama, K.; Karthik, K.; Khandia, R.; Munjal, A.; Khurana, S.K.; Chakraborty, S.; Malik, Y.S.; Virmani, N.; Singh, R.; et al. A comprehensive review of equine influenza virus: Etiology, epidemiology, pathobiology, and advances in diagnostics, vaccines, and control strategies. Front. Microbiol. 2018, 9, 1941. [Google Scholar] [CrossRef]
- Cowled, B.; Ward, M.P.; Hamilton, S.; Garner, G. The equine influenza epidemic in Australia: Spatial and temporal descriptive analyses of a large propagating epidemic. Prev. Vet. Med. 2009, 92, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.M.; Lai, A.C.K.; Binns, M.M.; Chambers, T.M.; Barrandeguy, M. Antigenic and genetic evolution of equine H3N8 influenza A viruses. J. Gen. Virol. 1996, 77, 661–671. [Google Scholar] [CrossRef]
- Timoney, P.J. Factors influencing the international spread of equine diseases. Vet. Clin. N. Am. Equine Pract. 2000, 16, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Shittu, I.; Meseko, C.A.; Sulaiman, L.P.; Inuwa, B.; Mustapha, M.; Zakariya, P.S.; Muhammad, A.A.; Muhammad, U.; Atuman, Y.J.; Barde, I.J.; et al. Fatal multiple outbreaks of equine influenza H3N8 in Nigeria, 2019: The first introduction of Florida clade 1 to West Africa. Vet. Microbiol. 2020, 248, 108820. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiao, Y.; Meng, F.; Sun, F.; Chen, M.; Cheng, Z.; Chen, Y.; Liu, S.; Chen, H. Emergence of H3N8 equine influenza virus in donkeys in China in 2017. Vet. Microbiol. 2018, 214, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Barbic, L.; Madic, J.; Turk, N.; Daly, J. Vaccine failure caused an outbreak of equine influenza in Croatia. Vet. Micobiol. 2009, 133, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Elia, G.; Decaro, N.; Trani, L.D.; Lorusso, E.; Campolo, M.; Desario, C.; Parisi, A.; Cavaliere, N.; Buonavoglia, C. An outbreak of equine influenza virus in vaccinated horses in Italy is due to an H3N8 strain related to recent North American representatives of the Florida sub-lineage. Vet. Microbiol. 2007, 121, 56–563. [Google Scholar] [CrossRef]
- Yamanaka, T.; Niwa, H.; Tsujimura, K.; Kondo, T.; Matsumura, T. Epidemic of equine influenza among vaccinated racehorses in Japan in 2007. J. Vet. Med. Sci. 2008, 70, 623–625. [Google Scholar] [CrossRef]
- Gildea, S.; Garvey, M.; Lyons, P.; Lyons, R.; Gahan, J.; Walsh, C.; Cullinane, A. Multifocal equine influenza outbreak with vaccination breakdown in thoroughbred racehorses. Pathogens 2018, 7, 43. [Google Scholar] [CrossRef]
- Favaro, P.F.; Fernandes, W.R.; Reischak, D.; Brandao, P.E.; Silva, S.O.S.; Richtzenhain, L.J. Evolution of equine influenza viruses (H3N8) during a Brazilian outbreak, 2015. Braz. J. Microbiol. 2018, 49, 336–346. [Google Scholar] [CrossRef]
- Park, A.W.; Wood, J.L.N.; Daly, J.M.; Newton, J.R.; Glass, K.; Henley, W.; Mumford, J.A.; Grenfell, B.T. The effects of strain heterology on the epidemiology of equine influenza in a vaccinated population. Proc. Biol. Sci. 2004, 271, 1547–1555. [Google Scholar] [CrossRef]
- Alaql, F.A.; Alhafufi, A.N.; Kasem, S.; Alhammad, Y.M.O.; Albaqshi, H.; Alyousaf, A.; Alsubaie, F.M.; Alghamdi, A.N.; Abdel-Moneim, A.S.; Alharbi, S.A. Full-length genome of the equine influenza A virus Subtype H3N8 from 2019 outbreak in Saudi Arabia. Animals 2022, 12, 2720. [Google Scholar] [CrossRef]
- Horsepia. Available online: http://horsepia.com (accessed on 26 July 2023).
- Nemoto, M.; Reedy, S.E.; Yano, T.; Suzuki, K.; Fukuda, S.; Garvey, M.; Kambayashi, Y.; Bannai, H.; Tsujimura, K.; Yamanaka, T.; et al. Antigenic comparison of H3N8 equine influenza viruses belonging to Florida sublineage clade 1 between vaccine strains and North American strains isolated in 2021–2022. Arch. Virol. 2023, 168, 94. [Google Scholar] [CrossRef] [PubMed]
- OIE. Chapter 2.5.7 Equine influenza. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2014. [Google Scholar]
- Abdelwhab, E.M.; Mettenleither, T.C. Zoonotic Animal Influenza Virus and Potential Mixing Vessel Hosts. Viruses 2023, 15, 980. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Number of Collected Samples | ||
|---|---|---|---|
| Blood | Nasal Swab | Lung | |
| 2020 | 1323 | 509 | 157 |
| 2021 | 1312 | 660 | 266 |
| 2022 | 1346 | 636 | 50 |
| Total | 3981 | 1805 | 473 |
| Year | Number of Tested | Number of Positive Horse | Positive Rate (%) | GMT |
|---|---|---|---|---|
| 2020 | 1323 | 1195 | 90.3 | 83.4 |
| 2021 | 1312 | 1269 | 96.7 | 135.7 |
| 2022 | 1346 | 1235 | 91.8 | 95.6 |
| Total | 3981 | 3699 | 92.9 | 103.1 |
| Type | 2020 | 2021 | 2022 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | |
| Race horse | 262 | 247 | 94.3 | 118.3 | 360 | 351 | 97.5 | 124.3 | 305 | 251 | 82.3 | 50.9 |
| Riding horse | 725 | 651 | 89.8 | 80.1 | 685 | 672 | 98.1 | 148.5 | 722 | 702 | 97.2 | 122.2 |
| Training horse | 44 | 26 | 59.1 | 112.0 | 18 | 7 | 38.9 | 78.0 | 34 | 15 | 44.1 | 67.0 |
| Broodmare | 261 | 244 | 93.5 | 63.6 | 210 | 207 | 98.6 | 103.3 | 238 | 223 | 93.7 | 95.9 |
| Stallion stud | 7 | 7 | 100 | 64.0 | 7 | 7 | 100 | 128.0 | 2 | 2 | 100 | 90.5 |
| Unknown | 24 | 20 | 83.3 | 81.6 | 32 | 32 | 100 | 189.0 | 45 | 42 | 93.3 | 76.7 |
| Total | 1323 | 1195 | 90.3 | 83.4 | 1312 | 1269 | 96.7 | 135.7 | 1346 | 1235 | 91.8 | 103.1 |
| Age | 2020 | 2021 | 2022 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | |
| 0–1 | 33 | 14 | 42.4 | 95.1 | 16 | 8 | 50.0 | 128.0 | 24 | 7 | 29.2 | 115.9 |
| 2 | 41 | 30 | 73.2 | 67.0 | 32 | 24 | 75.0 | 110.8 | 23 | 17 | 73.9 | 56.6 |
| 3–5 | 339 | 312 | 92.0 | 94.4 | 369 | 361 | 97.8 | 114.5 | 355 | 304 | 85.6 | 64.6 |
| 6–10 | 371 | 342 | 92.2 | 85.3 | 352 | 341 | 96.9 | 159.7 | 335 | 314 | 93.7 | 102.4 |
| >10 | 539 | 497 | 92.2 | 76.6 | 501 | 493 | 98.4 | 136.9 | 569 | 553 | 97.2 | 111.2 |
| Unknown | 0 | 0 | 0 | 0 | 42 | 42 | 100 | 158.6 | 40 | 40 | 100 | 163.1 |
| Total | 1323 | 1195 | 90.3 | 83.4 | 1312 | 1269 | 96.7 | 135.7 | 1346 | 1235 | 91.8 | 95.6 |
| Province * | 2020 | 2021 | 2022 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | No. Tested | No. of Positive Horse | Positive Rate (%) | GMT | |
| Gangwon (GW) | 50 | 48 | 96.0 | 66.8 | 47 | 47 | 100 | 171.9 | 57 | 57 | 100 | 128 |
| Gyeonggi (GG) | 462 | 433 | 93.7 | 84.7 | 460 | 447 | 97.2 | 111.8 | 477 | 429 | 89.9 | 83.1 |
| Chungbuk (CB) | 61 | 50 | 82.0 | 88.0 | 28 | 27 | 96.4 | 134.7 | 18 | 18 | 100 | 128 |
| Chungnam (CN) | 33 | 21 | 63.6 | 31.0 | 36 | 35 | 97.2 | 227.3 | 46 | 45 | 97.8 | 166.3 |
| Gyeongbuk (GB) | 42 | 40 | 95.2 | 102.2 | 55 | 55 | 100 | 150.8 | 52 | 48 | 92.3 | 81.8 |
| Gyeongnam (GN) | 86 | 78 | 90.7 | 95.5 | 148 | 147 | 99.3 | 189.3 | 153 | 142 | 92.8 | 78.6 |
| Jeonbuk (JB) | 72 | 60 | 83.3 | 80.6 | 50 | 50 | 100 | 191.3 | 61 | 57 | 93.4 | 71.4 |
| Jeonnam (JN) | 29 | 28 | 96.6 | 99.9 | 48 | 48 | 100 | 209.1 | 44 | 43 | 97.7 | 176.7 |
| Jeju Island (JJ) ** | 488 | 437 | 89.5 | 83.4 | 440 | 413 | 93.9 | 124.6 | 438 | 396 | 90.4 | 105.4 |
| Total | 1323 | 1195 | 90.3 | 83.4 | 1312 | 1269 | 96.7 | 135.7 | 1346 | 1235 | 91.8 | 95.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, S.-I.; Kim, M.J.; Kim, M.-J.; Lee, S.-K.; Yang, H.-S.; Kwon, M.; Lim, E.H.; Ouh, I.-O.; Kim, E.-J.; Hyun, B.-H.; et al. Assessment of Equine Influenza Virus Status in the Republic of Korea from 2020 to 2022. Viruses 2023, 15, 2135. https://doi.org/10.3390/v15102135
Lim S-I, Kim MJ, Kim M-J, Lee S-K, Yang H-S, Kwon M, Lim EH, Ouh I-O, Kim E-J, Hyun B-H, et al. Assessment of Equine Influenza Virus Status in the Republic of Korea from 2020 to 2022. Viruses. 2023; 15(10):2135. https://doi.org/10.3390/v15102135
Chicago/Turabian StyleLim, Seong-In, Min Ji Kim, Min-Ji Kim, Sang-Kyu Lee, Hyoung-Seok Yang, MiJung Kwon, Eui Hyeon Lim, In-Ohk Ouh, Eun-Jung Kim, Bang-Hun Hyun, and et al. 2023. "Assessment of Equine Influenza Virus Status in the Republic of Korea from 2020 to 2022" Viruses 15, no. 10: 2135. https://doi.org/10.3390/v15102135
APA StyleLim, S.-I., Kim, M. J., Kim, M.-J., Lee, S.-K., Yang, H.-S., Kwon, M., Lim, E. H., Ouh, I.-O., Kim, E.-J., Hyun, B.-H., & Lee, Y.-H. (2023). Assessment of Equine Influenza Virus Status in the Republic of Korea from 2020 to 2022. Viruses, 15(10), 2135. https://doi.org/10.3390/v15102135