
The Isolation and Characterization of Bacteriophages Infecting Avian Pathogenic Escherichia coli O1, O2 and O78 Strains

 , ,
, , 
 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria and Media
2.2. Sample Collection and Processing
2.3. Sample Enrichment and Phage Recovery
2.4. Purification and Phage Research Stock Preparation
2.5. Host Range and Lytic Activity
2.6. pH Tolerance
2.7. Transmission Electron Microscopy
2.8. DNA Extraction and Whole-Genome Sequencing
2.9. Bioinformatic Analysis of Sequencing Data
3. Results
3.1. Phage Isolation, Host Range and Lytic Activity
3.2. pH Tolerance
3.3. Phage Virion Morphology and Genomic Organization
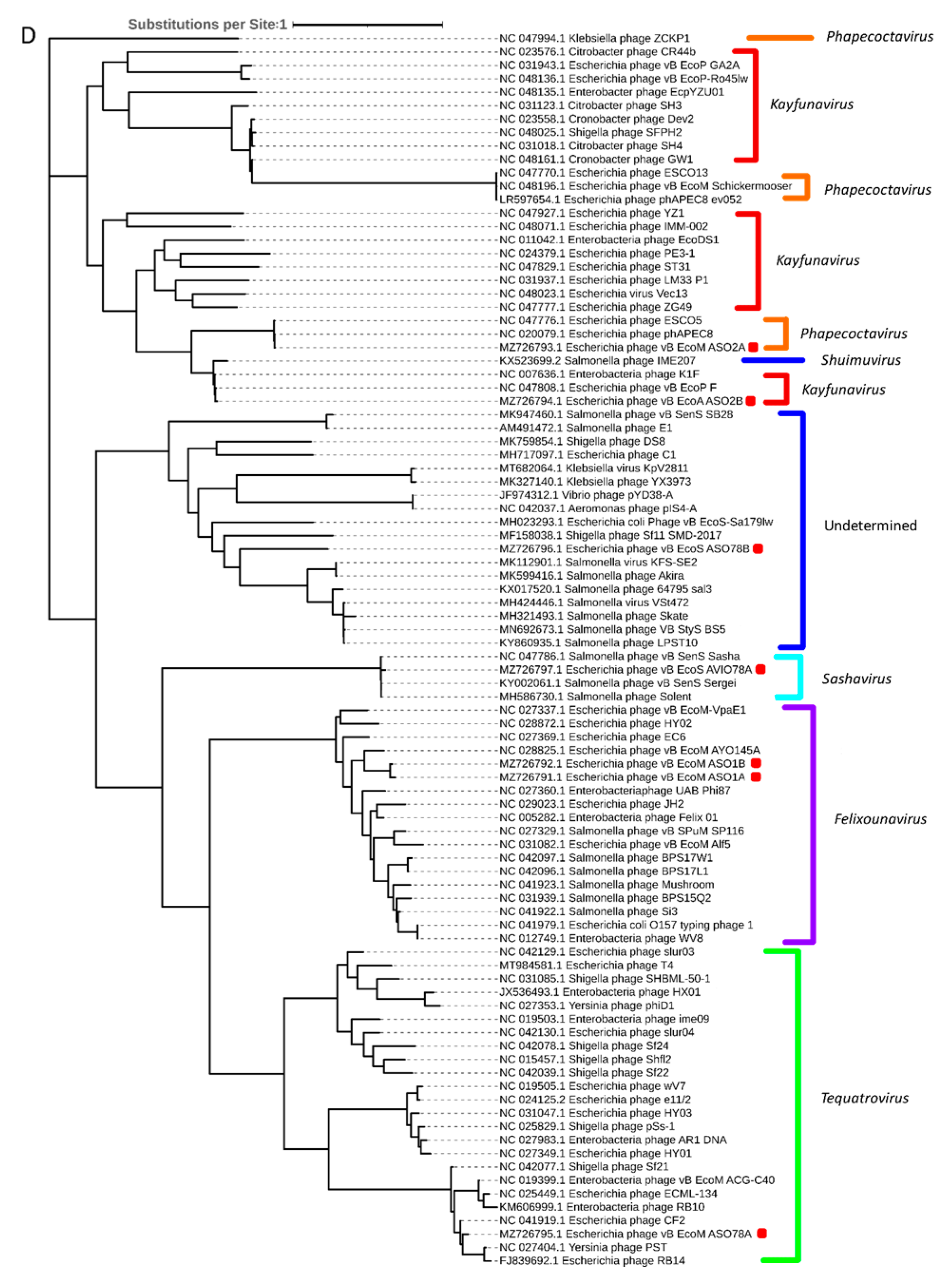
3.4. Comparative and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McPeake, S.; Smyth, J.; Ball, H. Characterisation of avian pathogenic Escherichia coli (APEC) associated with colisepticaemia compared to faecal isolates from healthy birds. Vet. Microbiol. 2005, 110, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Koutsianos, D.; Athanasiou, L.V.; Mossialos, D.; Franzo, G.; Cecchinato, M.; Koutoulis, K.C. Investigation of Serotype Prevalence of Escherichia coli Strains Isolated from Layer Poultry in Greece and Interactions with Other Infectious Agents. Vet. Sci. 2022, 9, 152. [Google Scholar] [CrossRef]
- La Ragione, R.; Woodward, M.J. Virulence factors of Escherichia coli serotypes associated with avian colisepticaemia. Res. Vet. Sci. 2002, 73, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Lutful Kabir, S. Avian colibacillosis and salmonellosis: A closer look at epidemiology, pathogenesis, diagnosis, control and public health concerns. Int. J. Environ. Res. Public Health 2010, 7, 89–114. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Boerlin, P.; Daignault, D.; Dozois, C.M.; Dutil, L.; Galanakis, C.; Reid-Smith, R.J.; Tellier, P.-P.; Tellis, P.A.; Ziebell, K. Food reservoir for Escherichia coli causing urinary tract infections. Emerg. Infect. Dis. 2010, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, C.R.; Prussing, C.; Boerlin, P.; Daignault, D.; Dutil, L.; Reid-Smith, R.J.; Zhanel, G.G.; Manges, A.R. Chicken as reservoir for extraintestinal pathogenic Escherichia coli in humans, Canada. Emerg. Infect. Dis. 2012, 18, 415–421. [Google Scholar] [CrossRef]
- Stromberg, Z.R.; Johnson, J.R.; Fairbrother, J.M.; Kilbourne, J.; Van Goor, A.; Curtiss 3rd, R.; Mellata, M. Evaluation of Escherichia coli isolates from healthy chickens to determine their potential risk to poultry and human health. PLoS ONE 2017, 12, e0180599. [Google Scholar] [CrossRef]
- Health Canada. Categorization of Antimicrobial Drugs Based on Importance in Human Medicine. 2009. Available online: https://www.canada.ca/en/health-canada/services/drugs-health-products/veterinary-drugs/antimicrobial-resistance/categorization-antimicrobial-drugs-based-importance-human-medicine.html (accessed on 10 November 2022).
- Chicken Farmers of Canada. Canadian Chicken Industry Reduces Antimicrobial Use. 2018. Available online: https://www.chickenfarmers.ca/wp-content/uploads/2018/10/AMU-Magazine-insides_ENG-Issue2.pdf (accessed on 24 August 2022).
- Christensen, H.; Bachmeier, J.; Bisgaard, M. New strategies to prevent and control avian pathogenic Escherichia coli (APEC). Avian Pathol. 2021, 50, 370–381. [Google Scholar] [CrossRef]
- Tarabees, R.; El-Sayed, M.S.; Shehata, A.A.; Diab, M.S. Effects of the probiotic candidate E. faecalis-1, the poulvac E. coli vaccine, and their combination on growth performance, caecal microbial composition, immune response, and protection against E. coli O78 challenge in broiler chickens. Probiot. Antimicrob. Proteins 2020, 12, 860–872. [Google Scholar]
- Uotani, Y.; Kitahara, R.; Imai, T.; Tsutsumi, N.; Sasakawa, C.; Nagai, S.; Nagano, T. Efficacy of an avian colibacillosis live vaccine for layer breeder in Japan. J. Vet. Med. Sci. 2017, 79, 1215–1219. [Google Scholar] [CrossRef][Green Version]
- Nagano, T.; Kitahara, R.; Nagai, S. An attenuated mutant of avian pathogenic Escherichia coli serovar O78: A possible live vaccine strain for prevention of avian colibacillosis. Microbiol. Immunol. 2012, 56, 605–612. [Google Scholar] [CrossRef]
- La Ragione, R.; Woodward, M.; Kumar, M.; Rodenberg, J.; Fan, H.; Wales, A.; Karaca, K. Efficacy of a live attenuated Escherichia coli O78: K80 vaccine in chickens and turkeys. Avian Dis. 2013, 57, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Guabiraba, R.; Schouler, C. Avian colibacillosis: Still many black holes. FEMS Microbiol. Lett. 2015, 362, fnv118. [Google Scholar] [CrossRef] [PubMed]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage applications for food production and processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef]
- Loc-Carrillo, C.; Abedon, S. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162–173. [Google Scholar] [CrossRef]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe Rev. 2019, 25, 219–232. [Google Scholar] [CrossRef]
- Huff, W.; Huff, G.; Rath, N.; Balog, J.; Donoghue, A. Alternatives to antibiotics: Utilization of bacteriophage to treat colibacillosis and prevent foodborne pathogens. Poult. Sci. 2005, 84, 655–659. [Google Scholar] [CrossRef]
- El-Gohary, F.; Huff, W.; Huff, G.; Rath, N.; Zhou, Z.; Donoghue, A. Environmental augmentation with bacteriophage prevents colibacillosis in broiler chickens. Poult. Sci. 2014, 93, 2788–2792. [Google Scholar] [CrossRef]
- Jamalludeen, N.; She, Y.-M.; Lingohr, E.; Griffiths, M. Isolation and characterization of virulent bacteriophages against Escherichia coli serogroups O1, O2, and O78. Poult. Sci. 2009, 88, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; McAllister, T.; Xu, Y.; Johnson, R.; Stephens, T.; Stanford, K. Prevalence and impact of bacteriophages on the presence of Escherichia coli O157: H7 in feedlot cattle and their environment. Appl. Environ. Microbiol. 2009, 75, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Niu, Y.D.; Stanford, K.; McAllister, T.A.; Ackermann, H.-W. Characterization of 4 T1-like lytic bacteriophages that lyse Shiga-toxin Escherichia coli O157: H7. Can. J. Microbiol. 2012, 58, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.D.; Johnson, R.P.; Xu, Y.; McAllister, T.A.; Sharma, R.; Louie, M.; Stanford, K. Host range and lytic capability of four bacteriophages against bovine and clinical human isolates of Shiga toxin-producing Escherichia coli O157:H7. J. Appl. Microbiol. 2009, 107, 646–656. [Google Scholar] [CrossRef]
- Niu, Y.D.; Stanford, K.; Kropinski, A.M.; Ackermann, H.W.; Johnson, R.P.; She, Y.M.; Ahmed, R.; Villegas, A.; McAllister, T.A. Genomic, proteomic and physiological characterization of a T5-like bacteriophage for control of Shiga toxin-producing Escherichia coli O157:H7. PLoS ONE 2012, 7, e34585. [Google Scholar] [CrossRef]
- Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of bacteriophages using the small drop plaque assay system. Methods Mol. Biol. 2009, 501, 81–85. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Köster, J.; Rahmann, S. Snakemake—A scalable bioinformatics workflow engine. Bioinformatics 2012, 28, 2520–2522. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, E.P.; Eddy, S.R. Infernal 1.1: 100-fold faster RNA homology searches. Bioinformatics 2013, 29, 2933–2935. [Google Scholar] [CrossRef] [PubMed]
- Kingsford, C.L.; Ayanbule, K.; Salzberg, S.L. Rapid, accurate, computational discovery of Rho-independent transcription terminators illuminates their relationship to DNA uptake. Genome Biol. 2007, 8, R22. [Google Scholar] [CrossRef]
- Mitra, A.; Kesarwani, A.K.; Pal, D.; Nagaraja, V. WebGeSTer DB—A transcription terminator database. Nucleic Acids Res. 2011, 39, D129–D135. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Mohamad Razali, R.; Bougouffa, S.; Radovanovic, A.; Bajic, V.B. bTSSfinder: A novel tool for the prediction of promoters in cyanobacteria and Escherichia coli. Bioinformatics 2017, 33, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. ABRicate: Mass Screening of Contigs for Antimicrobial and Virulence Genes: Github 2018. Available online: https://github.com/tseemann/abricate (accessed on 20 August 2023).
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.-M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, D.M.; Waller, A.S.; Yamada, T.; Bork, P.; Mushegian, A.R.; Koonin, E.V. Orthologous gene clusters and taxon signature genes for viruses of prokaryotes. J. Bacteriol. 2013, 195, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC—A novel tool to calculate the intergenomic similarities of prokaryote-infecting viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A roadmap for genome-based phage taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Turner, D.; Reynolds, D.; Seto, D.; Mahadevan, P. CoreGenes3. 5: A webserver for the determination of core genes from sets of viral and small bacterial genomes. BMC Res. Notes 2013, 6, 140. [Google Scholar] [CrossRef]
- Sullivan, M.; Petty, N.; Beatson, S. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Kropinski, A.M.; Prangishvili, D.; Lavigne, R.J.E.m. Position paper: The creation of a rational scheme for the nomenclature of viruses of Bacteria and Archaea. Environ. Microbiol. 2009, 11, 2775–2777. [Google Scholar] [CrossRef]
- Ackermann, H.-W. Phage classification and characterization. In Bacteriophages; Springer: New York, NY, USA, 2009; pp. 127–140. [Google Scholar]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef]
- Wintachai, P.; Surachat, K.; Singkhamanan, K. Isolation and Characterization of a Novel Autographiviridae Phage and Its Combined Effect with Tigecycline in Controlling Multidrug-Resistant Acinetobacter baumannii-Associated Skin and Soft Tissue Infections. Viruses 2022, 14, 194. [Google Scholar] [CrossRef]
- Adriaenssens, E.M.; Sullivan, M.B.; Knezevic, P.; van Zyl, L.J.; Sarkar, B.; Dutilh, B.E.; Alfenas-Zerbini, P.; Łobocka, M.; Tong, Y.; Brister, J.R. Taxonomy of prokaryotic viruses: 2018-2019 update from the ICTV Bacterial and Archaeal Viruses Subcommittee. Arch. Virol. 2020, 165, 1253–1260. [Google Scholar] [CrossRef]
- Pires, D.P.; Costa, A.R.; Pinto, G.; Meneses, L.; Azeredo, J. Current challenges and future opportunities of phage therapy. FEMS Microbiol. Rev. 2020, 44, 684–700. [Google Scholar] [CrossRef]
- Oliveira, A.; Sillankorva, S.; Quinta, R.; Henriques, A.; Sereno, R.; Azeredo, J. Isolation and characterization of bacteriophages for avian pathogenic E. coli strains. J. Appl. Microbiol. 2009, 106, 1919–1927. [Google Scholar] [CrossRef]
- Kathayat, D.; Helmy, Y.A.; Deblais, L.; Rajashekara, G. Novel small molecules affecting cell membrane as potential therapeutics for avian pathogenic Escherichia coli. Sci. Rep. 2018, 8, 15329. [Google Scholar] [CrossRef]
- Deng, S.; Xu, Q.; Fu, Y.; Liang, L.; Wu, Y.; Peng, F.; Gao, M. Genomic analysis of a novel phage infecting the Turkey pathogen Escherichia coli APEC O78 and its endolysin activity. Viruses 2021, 13, 1034. [Google Scholar] [CrossRef]
- Philipson, C.W.; Voegtly, L.J.; Lueder, M.R.; Long, K.A.; Rice, G.K.; Frey, K.G.; Biswas, B.; Cer, R.Z.; Hamilton, T.; Bishop-Lilly, K.A. Characterizing phage genomes for therapeutic applications. Viruses 2018, 10, 188. [Google Scholar] [CrossRef]
- Gohar, M.; Smith, R.; Hyun, J.; Mehina, N.; Abdelsalam, R.; Fairbrother, J.; Faizal, A.C.M.; Niu, D. Use of bacteriophage cocktail in the prevention of colibacillosis in laying hens. In Proceedings of the 72nd Annual Conference of the Canadian Society of Microbiologists, Halifax, NS, Canada, 25–28 June 2023. [Google Scholar]
- Plackett, B. Why big pharma has abandoned antibiotics. Nature 2020, 586, S50. [Google Scholar] [CrossRef]
- Parfitt, T. Georgia: An unlikely stronghold for bacteriophage therapy. Lancet 2005, 365, 2166–2167. [Google Scholar] [CrossRef]
- Torres-Acosta, M.A.; Clavijo, V.; Vaglio, C.; González-Barrios, A.F.; Vives-Flórez, M.J.; Rito-Palomares, M. Economic evaluation of the development of a phage therapy product for the control of Salmonella in poultry. Biotechnol. Prog. 2019, 35, e2852. [Google Scholar] [CrossRef]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Azam, A.H.; Tanji, Y. Bacteriophage-host arm race: An update on the mechanism of phage resistance in bacteria and revenge of the phage with the perspective for phage therapy. Appl. Microbiol. Biotechnol. 2019, 103, 2121–2131. [Google Scholar] [CrossRef]
- Abdelsattar, A.S.; Dawooud, A.; Rezk, N.; Makky, S.; Safwat, A.; Richards, P.J.; El-Shibiny, A. How to train your phage: The recent efforts in phage training. Biologics 2021, 1, 70–88. [Google Scholar] [CrossRef]
- Haggård-Ljungquist, E.; Halling, C.; Calendar, R. DNA sequences of the tail fiber genes of bacteriophage P2: Evidence for horizontal transfer of tail fiber genes among unrelated bacteriophages. J. Bacteriol. 1992, 174, 1462–1477. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phages | APEC Strains (Serogroup) | |||||
|---|---|---|---|---|---|---|
| ECL20834 (O1) | ECL20885 (O1) | ECL21443 (O2) | ECL22102 (O78) | ECL20719 (O78) | ECL23026 (O78) | |
| ASO1A | − | +++ | ++ | − | − | − |
| ASO1B | − | +++ | ++ | − | − | − |
| ASO2A | − | ++ | +++ | − | ++ | − |
| ASO2B | − | ++ | +++ | − | − | − |
| ASO78A | + | − | ++ | +++ | +++ | +++ |
| ASO78B | − | − | − | ± | +++ | +++ |
| AVIO78A | − | − | − | + | +++ | +++ |
| ASO78A + ASO78B | NA | NA | NA | +++ | +++ | +++ |
| ASO78A + AVIO78A | NA | NA | NA | +++ | +++ | +++ |
| ASO78B + AVIO78A | NA | NA | NA | ++ | +++ | +++ |
| ASO78A + ASO78B + AVIO78A | NA | NA | NA | +++ | +++ | +++ |
| Phage |
Assigned morphotype | Head Diameter (nm) | Tail Sheath Length (nm) | Tail Width (nm) |
| ASO1A | Myovirus | 61.4 ± 1.3 | 125.7 ± 3.3 | 20.2 ± 0.9 |
| ASO1B | Myovirus | 69.6 ± 2.4 | 108.1 ± 2.1 | 17.6 ± 0.6 |
| ASO2A | Myovirus | 83.4 ± 3.6 | 88.4 ± 1.3 | 17.8 ± 2.0 |
| ASO2B | Podovirus | 53.2 ± 0.8 | 10.6 ± 4.5 | 12.9 ± 3.4 |
| ASO78A | Myovirus | 97.3 ± 5.0 | 115.3 ± 6.4 | 27.3 ± 4.6 |
| ASO78B | Siphovirus | 61.8 ± 5.1 | 107.4 ± 4.8 | 10.6 ± 0 |
| AVIO78A | Siphovirus | 63.3 ± 2.4 | 157.6 ± 3.2 | 10.4 ± 0.60 |
| Genus | Phage | Genbank Accession Number | Genome Size (Kbp) | CDS | G+C% | tRNA | Promoters | Terminators |
| Felixounavirus | ASO1A | MZ726791 | 87.613 | 126 | 38.8 | 25 | 18 | 19 |
| Felixounavirus | ASO1B | MZ726792 | 89.496 | 127 | 38.8 | 26 | 19 | 20 |
| Phapecoctavirus | ASO2A | MZ726793 | 151.661 | 277 | 39.0 | 12 | 9 | 55 |
| Tequatrovirus | ASO78A | MZ726795 | 166.374 | 270 | 35.4 | 10 | 81 | 57 |
| Kayfunavirus | ASO2B | MZ726794 | 39.917 | 48 | 49.7 | 0 | 11 | 11 |
| Undetermined | ASO78B | MZ726796 | 46.233 | 81 | 46.6 | 2 | 5 | 15 |
| Sashavirus | AVIO78A | MZ726797 | 57.881 | 105 | 43.7 | 0 | 7 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, K.R.; Bumunang, E.W.; Schlechte, J.; Waldner, M.; Anany, H.; Walker, M.; MacLean, K.; Stanford, K.; Fairbrother, J.M.; Alexander, T.W.; et al. The Isolation and Characterization of Bacteriophages Infecting Avian Pathogenic Escherichia coli O1, O2 and O78 Strains. Viruses 2023, 15, 2095. https://doi.org/10.3390/v15102095
Smith KR, Bumunang EW, Schlechte J, Waldner M, Anany H, Walker M, MacLean K, Stanford K, Fairbrother JM, Alexander TW, et al. The Isolation and Characterization of Bacteriophages Infecting Avian Pathogenic Escherichia coli O1, O2 and O78 Strains. Viruses. 2023; 15(10):2095. https://doi.org/10.3390/v15102095
Chicago/Turabian StyleSmith, Kat R., Emmanuel W. Bumunang, Jared Schlechte, Matthew Waldner, Hany Anany, Matthew Walker, Kellie MacLean, Kim Stanford, John M. Fairbrother, Trevor W. Alexander, and et al. 2023. "The Isolation and Characterization of Bacteriophages Infecting Avian Pathogenic Escherichia coli O1, O2 and O78 Strains" Viruses 15, no. 10: 2095. https://doi.org/10.3390/v15102095
APA StyleSmith, K. R., Bumunang, E. W., Schlechte, J., Waldner, M., Anany, H., Walker, M., MacLean, K., Stanford, K., Fairbrother, J. M., Alexander, T. W., McAllister, T. A., Abdul-Careem, M. F., & Niu, Y. D. (2023). The Isolation and Characterization of Bacteriophages Infecting Avian Pathogenic Escherichia coli O1, O2 and O78 Strains. Viruses, 15(10), 2095. https://doi.org/10.3390/v15102095