
Evolution of Swine Influenza Virus H3N2 in Vaccinated and Nonvaccinated Pigs after Previous Natural H1N1 Infection

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell and Viruses
2.2. Experimental Design
2.3. Humoral Response against SIV
2.4. Pathological Analyses in Lung and Immunohistochemistry to Detect SIV
2.5. SIV RNA Detection
2.6. Whole Influenza Genome Sequence by NGS
2.7. De Novo Assambly, Mapping, and Variant Calling
2.8. Nucleotide Diversity (π)
2.9. Lolliplot and 3D Structural Representation of SIV Proteins
2.10. Satatistical Analysis
3. Results
3.1. Detection of Natural SIV Circulation in Both Experimental Groups at Vaccination Day
3.2. Identification of H1N1 Subtype as Natural Circulating Virus before Challenge
3.3. Humoral Response againts SIV after Vaccination and H3N2 Challenge
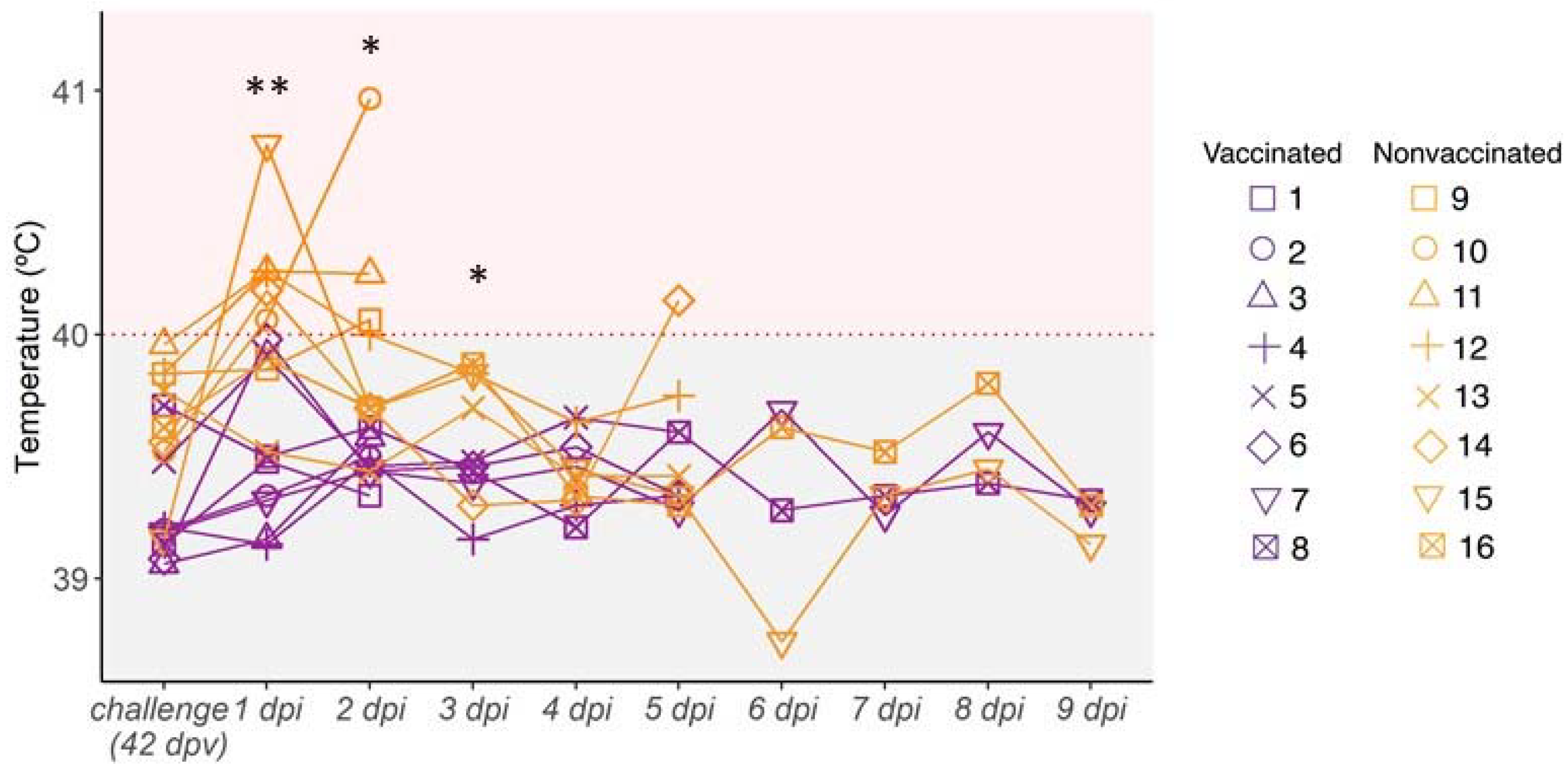
3.4. Kinetics of Rectal Temperature after SIV Challenge
3.5. SIV Genome Detection after Challenge
3.6. Lung Gross Lesions, Histopathological Assessment, and Immunohistochemistry to Detect SIV Antigen
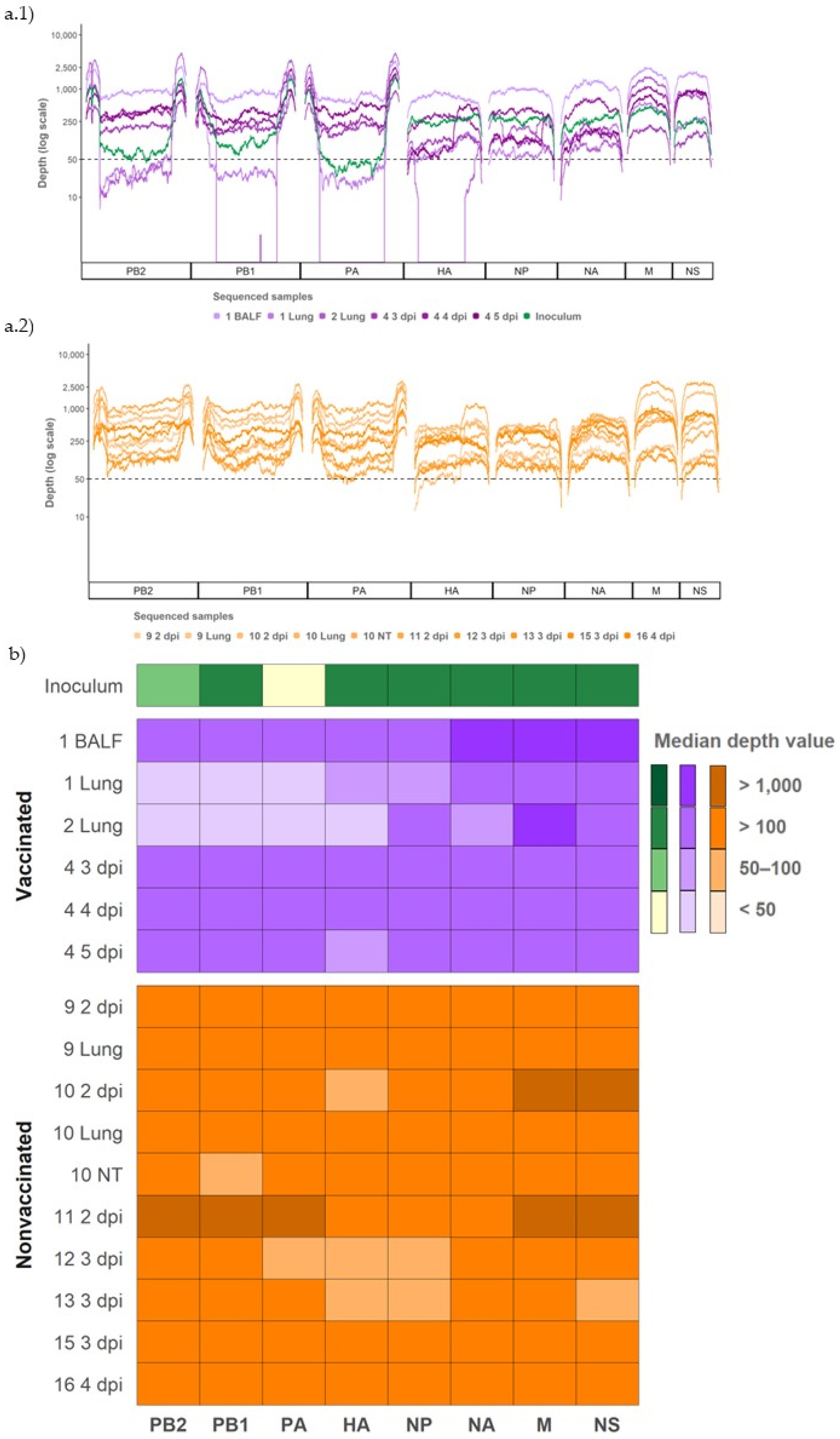
3.7. Whole SIV Genome Sequences Obtained from Vaccinated and Nonvaccinated Challenged Animals
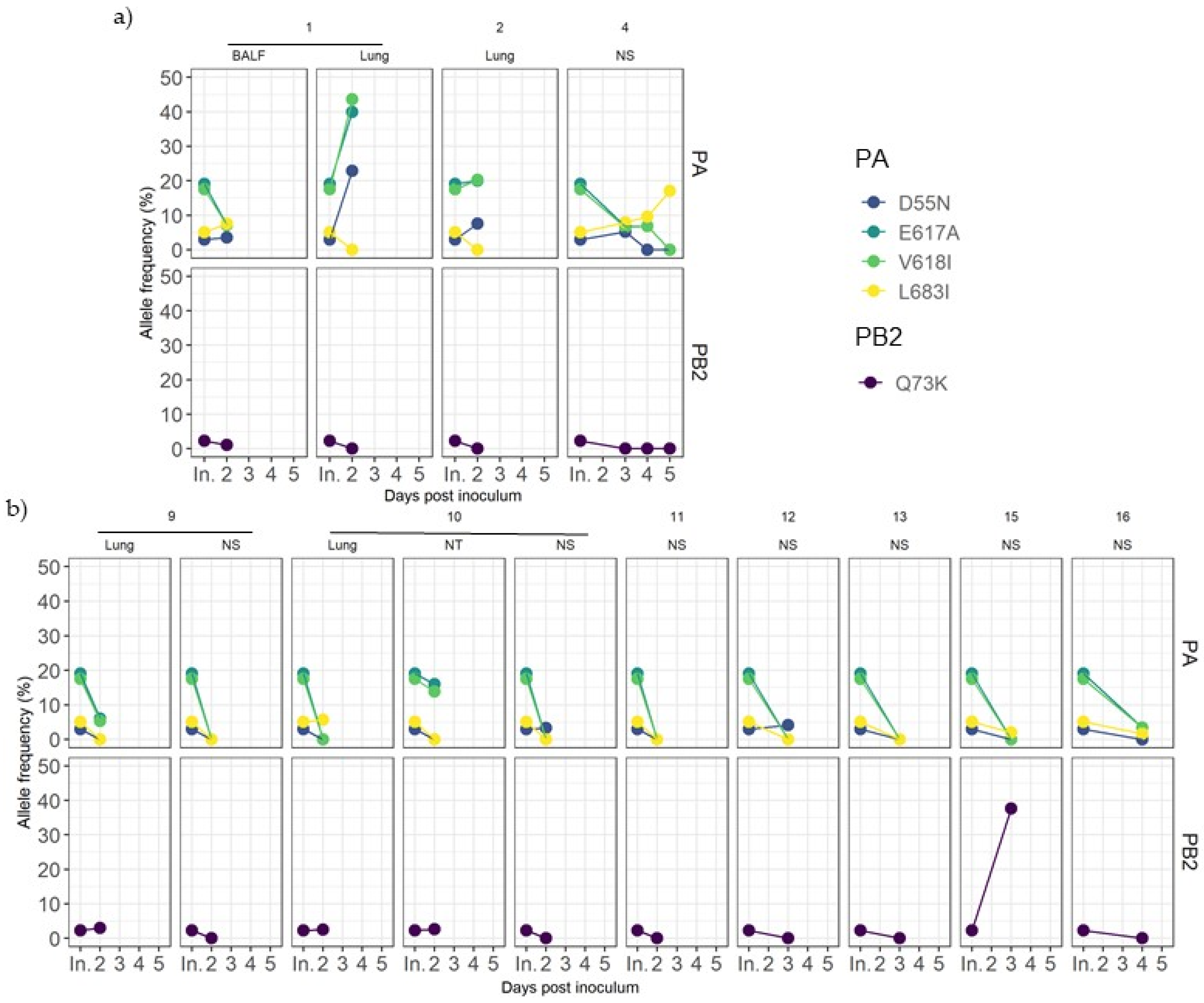
3.8. Evolution of Variants Found in Inoculum
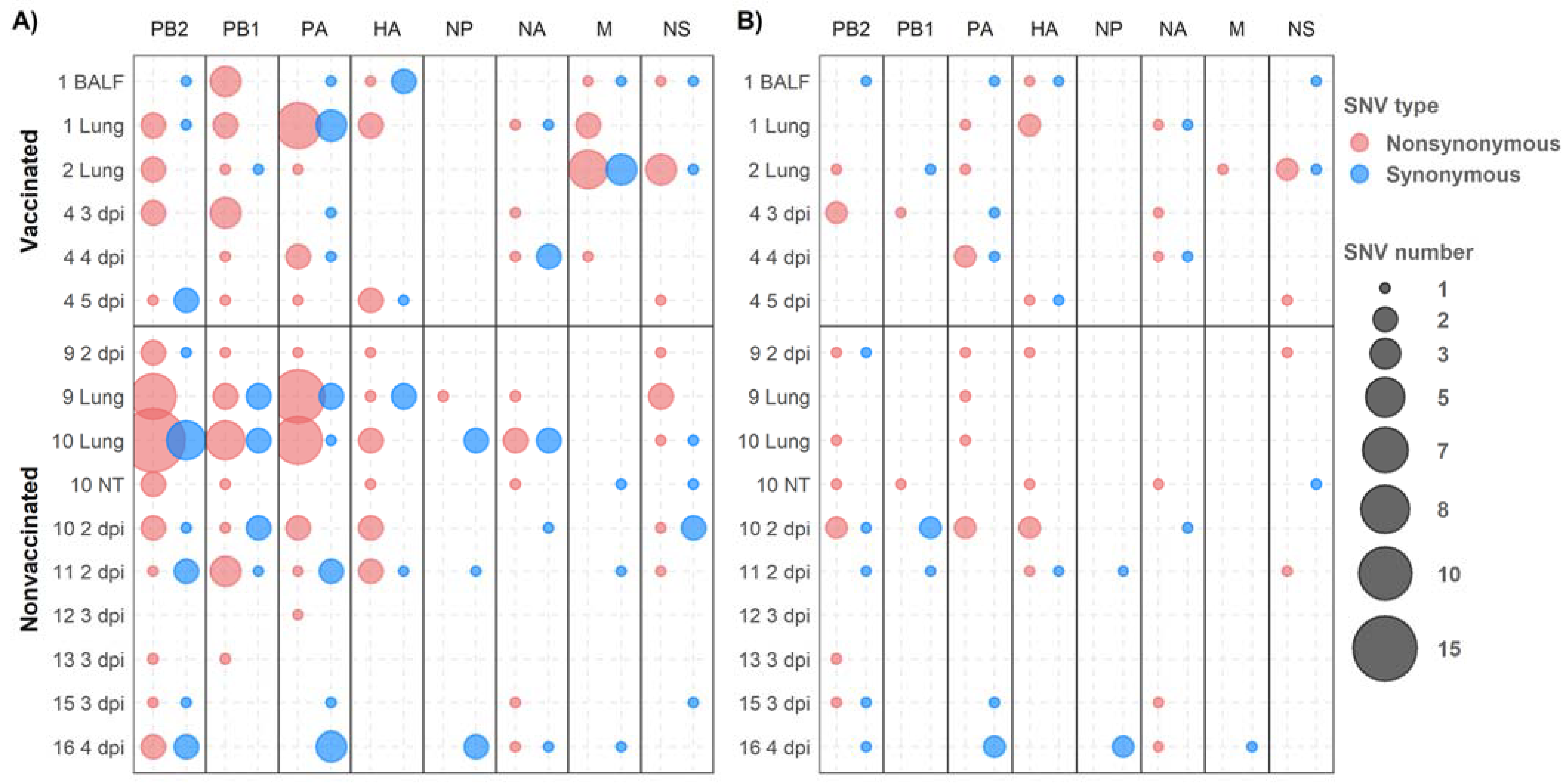
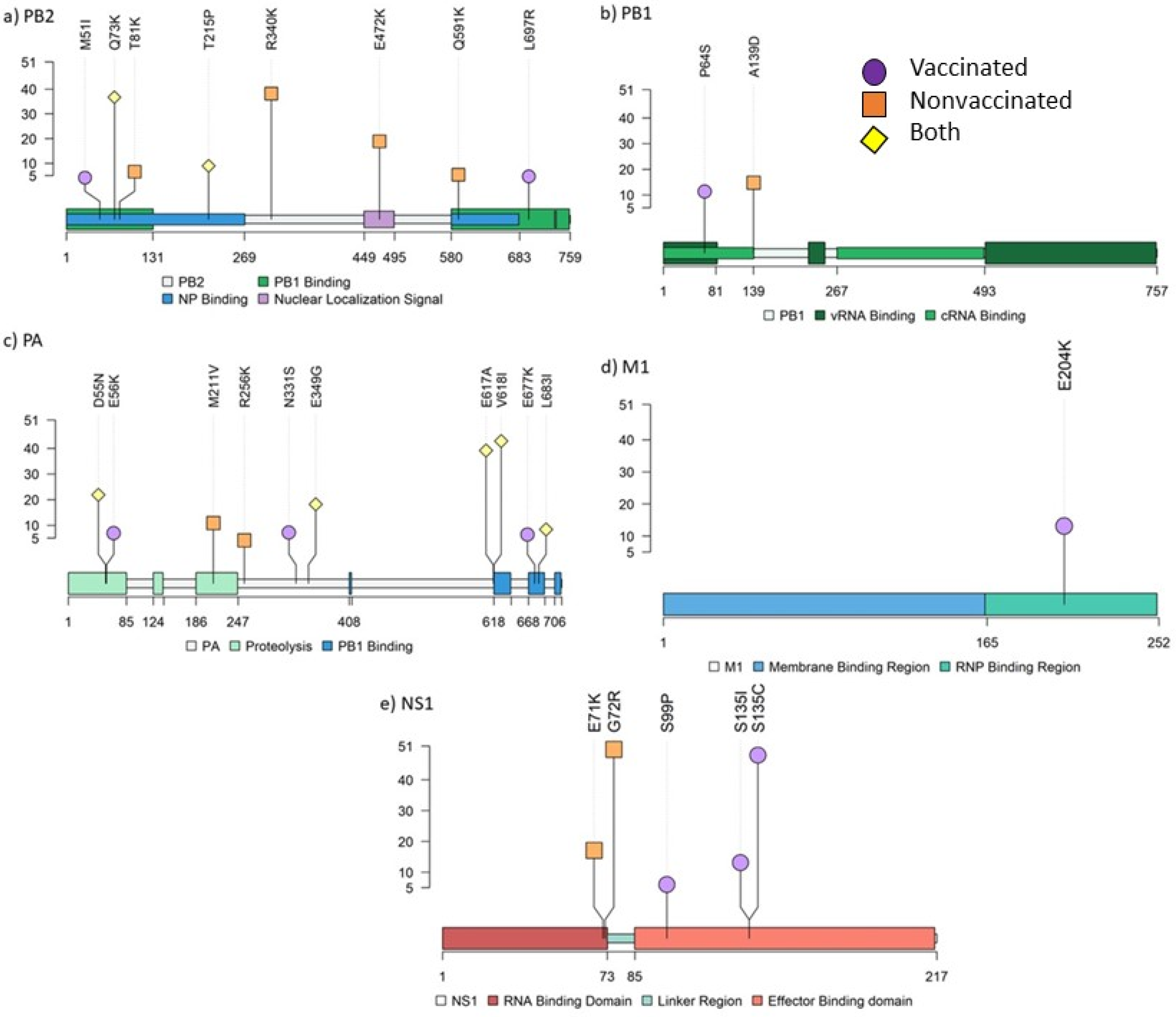
3.9. Detection of De Novo SIV Variants in Sequenced Samples Collected from Vaccinated and Nonvaccinated Animals
3.10. Genetic Diversity in Viral Populations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Organisation for Animal Health. Swine Influenza. Available online: https://www.oie.int/en/disease/swine-influenza/ (accessed on 22 February 2022).
- Li, Y.; Robertson, I. The Epidemiology of Swine Influenza. Anim. Dis. 2021, 1, 21. [Google Scholar] [CrossRef] [PubMed]
- Breen, M.; Nogales, A.; Baker, S.F.; Martínez-Sobrido, L. Replication-Competent Influenza A Viruses Expressing Reporter Genes. Viruses 2016, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Skehel, J.J.; Wiley, D.C. Receptor Binding and Membrane Fusion in Virus Entry: The Influenza Hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef]
- Varghese, J.N.; McKimm-Breschkin, J.L.; Caldwell, J.B.; Kortt, A.A.; Colman, P.M. The Structure of the Complex between Influenza Virus Neuraminidase and Sialic Acid, the Viral Receptor. Proteins 1992, 14, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Aramini, J.M.; Ma, L.-C.; Krug, R.M.; Arnold, E. Structures of Influenza A Proteins and Insights into Antiviral Drug Targets. Nat. Struct. Mol. Biol. 2010, 17, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.S.; Russell, C.A.; Langat, P.; Anderson, T.K.; Berger, K.; Bielejec, F.; Burke, D.F.; Dudas, G.; Fonville, J.M.; Fouchier, R.A.M.; et al. The Global Antigenic Diversity of Swine Influenza A Viruses. Elife 2016, 5, e12217. [Google Scholar] [CrossRef] [PubMed]
- Holzer, B.; Martini, V.; Edmans, M.; Tchilian, E. T and B Cell Immune Responses to Influenza Viruses in Pigs. Front. Immunol. 2019, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Nichol, K.L.; Treanor, J.J. Vaccines for Seasonal and Pandemic Influenza. J. Infect. Dis. 2006, 194, S111–S118. [Google Scholar] [CrossRef]
- Ma, W.; Richt, J.A. Swine Influenza Vaccines: Current Status and Future Perspectives. Anim. Health Res. Rev. 2010, 11, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Thacker, E.; Janke, B. Swine Influenza Virus: Zoonotic Potential and Vaccination Strategies for the Control of Avian and Swine Influenzas. J. Infect. Dis. 2008, 197, S19–S24. [Google Scholar] [CrossRef]
- López-Valiñas, Á.; Sisteré-Oró, M.; López-Serrano, S.; Baioni, L.; Darji, A.; Chiapponi, C.; Segalés, J.; Ganges, L.; Núñez, J.I. Identification and Characterization of Swine Influenza Virus H1n1 Variants Generated in Vaccinated and Nonvaccinated, Challenged Pigs. Viruses 2021, 13, 2087. [Google Scholar] [CrossRef] [PubMed]
- Fulton, B.O.; Sachs, D.; Beaty, S.M.; Won, S.T.; Lee, B.; Palese, P.; Heaton, N.S. Mutational Analysis of Measles Virus Suggests Constraints on Antigenic Variation of the Glycoproteins. Cell Rep. 2015, 11, 1331. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S. Pathogen Evolution and the Immunological Niche. Ann. N. Y. Acad. Sci. 2014, 1320, 1. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Sheldon, J.; Perales, C. Viral Quasispecies Evolution. Microbiol. Mol. Biol. Rev. 2012, 76, 159–216. [Google Scholar] [CrossRef]
- Shao, W.; Li, X.; Goraya, M.U.; Wang, S.; Chen, J.L. Evolution of Influenza a Virus by Mutation and Re-Assortment. Int. J. Mol. Sci. 2017, 18, 1650. [Google Scholar] [CrossRef]
- Doud, M.B.; Lee, J.M.; Bloom, J.D. How Single Mutations Affect Viral Escape from Broad and Narrow Antibodies to H1 Influenza Hemagglutinin. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Prachanronarong, K.L.; Canale, A.S.; Liu, P.; Somasundaran, M.; Hou, S.; Poh, Y.-P.; Han, T.; Zhu, Q.; Renzette, N.; Zeldovich, K.B.; et al. Mutations in Influenza A Virus Neuraminidase and Hemagglutinin Confer Resistance against a Broadly Neutralizing Hemagglutinin Stem Antibody. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Kirkpatrick Roubidoux, E.; McMahon, M.; Carreño, J.M.; Capuano, C.; Jiang, K.; Simon, V.; van Bakel, H.; Wilson, P.; Krammer, F. Identification and Characterization of Novel Antibody Epitopes on the N2 Neuraminidase. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Krammer, F. The Human Antibody Response to Influenza A Virus Infection and Vaccination. Nat. Rev. Immunol. 2019, 19, 383–397. [Google Scholar] [CrossRef]
- Park, J.K.; Xiao, Y.; Ramuta, M.D.; Rosas, L.A.; Fong, S.; Matthews, A.M.; Freeman, A.D.; Gouzoulis, M.A.; Batchenkova, N.A.; Yang, X.; et al. Pre-Existing Immunity to Influenza Virus Hemagglutinin Stalk Might Drive Selection for Antibody-Escape Mutant Viruses in a Human Challenge Model. Nat. Med. 2020, 26, 1240–1246. [Google Scholar] [CrossRef]
- Zhou, N.N.; Senne, D.A.; Landgraf, J.S.; Swenson, S.L.; Erickson, G.; Rossow, K.; Liu, L.; Yoon, K.; Krauss, S.; Webster, R.G. Genetic Reassortment of Avian, Swine, and Human Influenza A Viruses in American Pigs. J. Virol. 1999, 73, 8851–8856. [Google Scholar] [CrossRef] [PubMed]
- Baratelli, M.; Córdoba, L.; Pérez, L.J.; Maldonado, J.; Fraile, L.; Núñez, J.I.; Montoya, M. Genetic Characterization of Influenza A Viruses Circulating in Pigs and Isolated in North-East Spain during the Period 2006–2007. Res. Vet. Sci. 2014, 96, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Janke, B.H.; Richt, J.A. Chapter 3 Swine Influenza Viruses: A North American Perspective. Adv Virus Res 2008, 72, 127–154. [Google Scholar] [CrossRef]
- Garten, R.J.; Davis, C.T.; Russell, C.A.; Shu, B.; Lindstrom, S.; Balish, A.; Sessions, W.M.; Xu, X.; Skepner, E.; Deyde, V.; et al. Antigenic and Genetic Characteristics of Swine-Origin 2009 A(H1N1) Influenza Viruses Circulating in Humans. Science 2009, 325, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Kahn, R.E.; Richt, J.A. The Pig as a Mixing Vessel for Influenza Viruses: Human and Veterinary Implications. J. Mol. Genet. Med. 2009, 3, 158. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty per Cent Endpoints. Antioch Rev. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Busquets, N.; Segalés, J.; Córdoba, L.; Mussá, T.; Crisci, E.; Martín-Valls, G.E.; Simon-Grifé, M.; Pérez-Simó, M.; Pérez-Maíllo, M.; Núñez, J.I.; et al. Experimental Infection with H1N1 European Swine Influenza Virus Protects Pigs from an Infection with the 2009 Pandemic H1N1 Human Influenza Virus. Vet. Res. 2010, 41, 74. [Google Scholar] [CrossRef]
- López-Serrano, S.; Cordoba, L.; Pérez-Maillo, M.; Pleguezuelos, P.; Remarque, E.J.; Ebensen, T.; Guzmán, C.A.; Christensen, D.; Segalés, J.; Darji, A. Immune Responses to Pandemic H1N1 Influenza Virus Infection in Pigs Vaccinated with a Conserved Hemagglutinin HA1 Peptide Adjuvanted with CAF®01 or CDA/AGalCerMPEG. Vaccines 2021, 9, 751. [Google Scholar] [CrossRef]
- Galindo-Cardiel, I.; Ballester, M.; Solanes, D.; Nofrarías, M.; López-Soria, S.; Argilaguet, J.M.; Lacasta, A.; Accensi, F.; Rodríguez, F.; Segalés, J. Standardization of Pathological Investigations in the Framework of Experimental ASFV Infections. Virus Res. 2013, 173, 180–190. [Google Scholar] [CrossRef]
- Sibila, M.; Aragón, V.; Fraile, L.; Segalés, J. Comparison of Four Lung Scoring Systems for the Assessment of the Pathological Outcomes Derived from Actinobacillus Pleuropneumoniae Experimental Infections. BMC Vet. Res. 2014, 10, 165. [Google Scholar] [CrossRef]
- ImageJ. Available online: https://imagej.nih.gov/ij/ (accessed on 14 January 2022).
- Sisteré-Oró, M.; López-Serrano, S.; Veljkovic, V.; Pina-Pedrero, S.; Vergara-Alert, J.; Córdoba, L.; Pérez-Maillo, M.; Pleguezuelos, P.; Vidal, E.; Segalés, J.; et al. DNA Vaccine Based on Conserved HA-Peptides Induces Strong Immune Response and Rapidly Clears Influenza Virus Infection from Vaccinated Pigs. PLoS ONE 2019, 14, e0222201. [Google Scholar] [CrossRef] [PubMed]
- Sabattini, E.; Bisgaard, K.; Ascani, S.; Poggi, S.; Piccioli, M.; Ceccarelli, C.; Pieri, F.; Fraternali-Orcioni, G.; Pileri, S.A. The EnVision(TM)+ System: A New Immunohistochemical Method for Diagnostics and Research. Critical Comparison with the APAAP, ChemMate(TM), CSA, LABC, and SABC Techniques. J. Clin. Pathol. 1998, 51, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Detmer, S.E.; Gunvaldsen, R.E.; Harding, J.C. Comparison of Influenza a Virus Infection in High- and Low-Birth-Weight Pigs Using Morphometric Analysis. Influenza Other Respir. Viruses 2013, 7, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a Real-Time Reverse Transcriptase PCR Assay for Type A Influenza Virus and the Avian H5 and H7 Hemagglutinin Subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef]
- Zhou, B.; Donnelly, M.E.; Scholes, D.T.; St. George, K.; Hatta, M.; Kawaoka, Y.; Wentworth, D.E. Single-Reaction Genomic Amplification Accelerates Sequencing and Vaccine Production for Classical and Swine Origin Human Influenza A Viruses. J. Virol. 2009, 83, 10309–10313. [Google Scholar] [CrossRef]
- Lycett, S.J.; Baillie, G.; Coulter, E.; Bhatt, S.; Kellam, P.; McCauley, J.W.; Wood, J.L.N.; Brown, I.H.; Pybus, O.G.; Brown Leigh, A.J. Estimating Reassortment Rates in Co-Circulating Eurasian Swine Influenza Viruses. J. Gen. Virol. 2012, 93, 2326–2336. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 September 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. Original Articles SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Mussá, T.; Ballester, M.; Silva-Campa, E.; Baratelli, M.; Busquets, N.; Lecours, M.P.; Dominguez, J.; Amadori, M.; Fraile, L.; Hernández, J.; et al. Swine, Human or Avian Influenza Viruses Differentially Activates Porcine Dendritic Cells Cytokine Profile. Vet. Immunol. Immunopathol. 2013, 154, 25–35. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, 1–4. [Google Scholar] [CrossRef] [PubMed]
- RStudio|Open Source & Professional Software for Data Science Teams-RStudio. Available online: https://www.rstudio.com/ (accessed on 1 September 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Wilm, A.; Aw, P.P.K.; Bertrand, D.; Yeo, G.H.T.; Ong, S.H.; Wong, C.H.; Khor, C.C.; Petric, R.; Hibberd, M.L.; Nagarajan, N. LoFreq: A Sequence-Quality Aware, Ultra-Sensitive Variant Caller for Uncovering Cell-Population Heterogeneity from High-Throughput Sequencing Datasets. Nucleic Acids Res. 2012, 40, 11189. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A Program for Annotating and Predicting the Effects of Single Nucleotide Polymorphisms, SnpEff: SNPs in the Genome of Drosophila Melanogaster Strain W1118; Iso-2; Iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.W.; Moncla, L.H.; Hughes, A.L. SNPGenie: Estimating Evolutionary Parameters to Detect Natural Selection Using Pooled next-Generation Sequencing Data. Bioinformatics 2015, 31, 3709–3711. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Zhu, L.J. TrackViewer: A Bioconductor Package for Interactive and Integrative Visualization of Multi-Omics Data. Nature Methods 2019 16:6 2019, 16, 453–454. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Wang, B.; Zhang, Y.; Zhu, Y.; Song, B.; Xu, H.; Zhai, Y.; Qiao, M.; Sun, F. A Cryo-Electron Microscopy Support Film Formed by 2D Crystals of Hydrophobin HFBI. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Zhu, X.; McBride, R.; Nycholat, C.M.; Yu, W.; Paulson, J.C.; Wilson, I.A. Influenza Virus Neuraminidases with Reduced Enzymatic Activity That Avidly Bind Sialic Acid Receptors. J. Virol. 2012, 86, 13371. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 27 April 2022).
- Kyriakis, C.S.; Rose, N.; Foni, E.; Maldonado, J.; Loeffen, W.L.A.; Madec, F.; Simon, G.; van Reeth, K. Influenza A Virus Infection Dynamics in Swine Farms in Belgium, France, Italy and Spain, 2006-2008. Vet. Microbiol. 2013, 162, 543–550. [Google Scholar] [CrossRef]
- Torremorell, M.; Allerson, M.; Corzo, C.; Diaz, A.; Gramer, M. Transmission of Influenza A Virus in Pigs. Transbound. Emerg. Dis. 2012, 59, 68–84. [Google Scholar] [CrossRef]
- Simon-Grifé, M.; Martín-Valls, G.E.; Vilar, M.J.; García-Bocanegra, I.; Mora, M.; Martín, M.; Mateu, E.; Casal, J. Seroprevalence and Risk Factors of Swine Influenza in Spain. Vet. Microbiol. 2011, 149, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Salvesen, H.A.; Whitelaw, C.B.A. Current and Prospective Control Strategies of Influenza A Virus in Swine. Porc. Health Manag. 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; de Hert, K.; van Reeth, K. Cross-Protection against European Swine Influenza Viruses in the Context of Infection Immunity against the 2009 Pandemic H1N1 Virus: Studies in the Pig Model of Influenza. Vet. Res. 2015, 46. [Google Scholar] [CrossRef] [PubMed]
- Mancera Gracia, J.C.; Pearce, D.S.; Masic, A.; Balasch, M. Influenza A Virus in Swine: Epidemiology, Challenges and Vaccination Strategies. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Davis, A.S.; Jagger, B.W.; Schwartzman, L.M.; Dunham, E.J.; Kash, J.C.; Taubenberger, J.K. Analysis by Single-Gene Reassortment Demonstrates That the 1918 Influenza Virus Is Functionally Compatible with a Low-Pathogenicity Avian Influenza Virus in Mice. J. Virol. 2012, 86, 9211–9220. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, Q.; Shi, J.; Deng, G.; Guo, J.; Zeng, X.; He, X.; Kong, H.; Gu, C.; Li, X.; Liu, J.; et al. H7N9 Influenza Viruses Are Transmissible in Ferrets by Respiratory Droplet. Science 2013, 341, 410–414. [Google Scholar] [CrossRef]
- Sobel Leonard, A.; McClain, M.T.; Smith, G.J.D.; Wentworth, D.E.; Halpin, R.A.; Lin, X.; Ransier, A.; Stockwell, T.B.; Das, S.R.; Gilbert, A.S.; et al. Deep Sequencing of Influenza A Virus from a Human Challenge Study Reveals a Selective Bottleneck and Only Limited Intrahost Genetic Diversification. J. Virol. 2016, 90, 11247–11258. [Google Scholar] [CrossRef]
- Domingo, E.; García-Crespo, C.; Lobo-Vega, R.; Perales, C. Mutation Rates, Mutation Frequencies, and Proofreading-Repair Activities in RNA Virus Genetics. Viruses 2021, 13, 1882. [Google Scholar] [CrossRef]
- Martín-Benito, J.; Ortín, J. Influenza Virus Transcription and Replication. Adv. Virus. Res. 2013, 87, 113–137. [Google Scholar] [CrossRef]
- Poole, E.; Elton, D.; Medcalf, L.; Digard, P. Functional Domains of the Influenza A Virus PB2 Protein: Identification of NP- and PB1-Binding Sites. Virology 2004, 321, 120–133. [Google Scholar] [CrossRef]
- Xiao, C.; Ma, W.; Sun, N.; Huang, L.; Li, Y.; Zeng, Z.; Wen, Y.; Zhang, Z.; Li, H.; Li, Q.; et al. PB2-588 V Promotes the Mammalian Adaptation of H10N8, H7N9 and H9N2 Avian Influenza Viruses. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Lina, L.; Saijuan, C.; Chengyu, W.; Yuefeng, L.; Shishan, D.; Ligong, C.; Kangkang, G.; Zhendong, G.; Jiakai, L.; Jianhui, Z.; et al. Adaptive Amino Acid Substitutions Enable Transmission of an H9N2 Avian Influenza Virus in Guinea Pigs. Sci. Rep. 2019, 9, 19734. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lee, H.H.Y.; Yang, Z.F.; Mok, C.K.P.; Zhang, Z. PB2-Q591K Mutation Determines the Pathogenicity of Avian H9N2 Influenza Viruses for Mammalian Species. PLoS ONE 2016, 11, e0162163. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Zeng, W.; Hao, X.; Huang, J.; Cai, M.; Zhou, P.; Zhang, G. Continuous Evolution of Influenza A Viruses of Swine from 2013 to 2015 in Guangdong, China. PLoS ONE 2019, 14, e0217607. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xu, Q.; Zhong, Y.; Yu, H.; Shu, J.; Ma, T.; Li, Z. Genetic Variation and Co-Evolutionary Relationship of RNA Polymerase Complex Segments in Influenza A Viruses. Virology 2017, 511, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Rolling, T.; Koerner, I.; Zimmermann, P.; Holz, K.; Haller, O.; Staeheli, P.; Kochs, G. Adaptive Mutations Resulting in Enhanced Polymerase Activity Contribute to High Virulence of Influenza A Virus in Mice. J. Virol. 2009, 83, 6673–6680. [Google Scholar] [CrossRef] [PubMed]
- Slaine, P.D.; MacRae, C.; Kleer, M.; Lamoureux, E.; McAlpine, S.; Warhuus, M.; Comeau, A.M.; McCormick, C.; Hatchette, T.; Khaperskyy, D.A. Adaptive Mutations in Influenza A/California/07/2009 Enhance Polymerase Activity and Infectious Virion Production. Viruses 2018, 10, 272. [Google Scholar] [CrossRef]
- Mitra, S.; Kumar, D.; Hu, L.; Sankaran, B.; Moosa, M.M.; Rice, A.P.; Ferreon, J.C.; Ferreon, A.C.M.; Prasad, B.V.V. Influenza A Virus Protein NS1 Exhibits Strain-Independent Conformational Plasticity. J. Virol. 2019, 93, 917–936. [Google Scholar] [CrossRef]
- Thepmalee, C.; Sanguansermsri, P.; Suwanankhon, N.; Chamnanpood, C.; Chamnanpood, P.; Pongcharoen, S.; Niumsap, P.R.; Surangkul, D.; Sanguansermsri, D. Changes in the NS1 Gene of Avian Influenza Viruses Isolated in Thailand Affect Expression of Type I Interferon in Primary Chicken Embryonic Fibroblast Cells. Indian J. Virol. 2013, 24, 365. [Google Scholar] [CrossRef][Green Version]
- Jia, D.; Rahbar, R.; Chan, R.W.Y.; Lee, S.M.Y.; Chan, M.C.W.; Wang, B.X.; Baker, D.P.; Sun, B.; Malik Peiris, J.S.; Nicholls, J.M.; et al. Influenza Virus Non-Structural Protein 1 (NS1) Disrupts Interferon Signaling. PLoS ONE 2010, 5, 13927. [Google Scholar] [CrossRef]
- Koliopoulos, M.G.; Lethier, M.; van der Veen, A.G.; Haubrich, K.; Hennig, J.; Kowalinski, E.; Stevens, R.V.; Martin, S.R.; Reis E Sousa, C.; Cusack, S.; et al. Molecular Mechanism of Influenza A NS1-Mediated TRIM25 Recognition and Inhibition. Nat. Commun. 2018, 9, 1820. [Google Scholar] [CrossRef]
- Pereira, C.F.; Wise, H.M.; Kurian, D.; Pinto, R.M.; Amorim, M.J.; Gill, A.C.; Digard, P. Effects of Mutations in the Effector Domain of Influenza A Virus NS1 Protein. BMC Res. Notes 2018, 11, 673. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Domingues, P.; Zell, R.; Hale, B.G. Structure-Guided Functional Annotation of the Influenza A Virus NS1 Protein Reveals Dynamic Evolution of the P85β-Binding Site during Circulation in Humans. J. Virol. 2017, 91, 1081–1098. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.-K.; Li, Y.; Liu, Q.; Anderson, D.H.; Babiuk, L.A.; Zhou, Y. SH3 Binding Motif 1 in Influenza A Virus NS1 Protein Is Essential for PI3K/Akt Signaling Pathway Activation. J. Virol. 2007, 81, 12730–12739. [Google Scholar] [CrossRef]
- Rossman, J.S.; Lamb, R.A. Influenza Virus Assembly and Budding. Virology 2011, 411, 229. [Google Scholar] [CrossRef] [PubMed]
- Elleman, C.J.; Barclay, W.S. The M1 Matrix Protein Controls the Filamentous Phenotype of Influenza A Virus. Virology 2004, 321, 144–153. [Google Scholar] [CrossRef]
- Biswas, S.K.; Boutz, P.L.; Nayak, D.P. Influenza Virus Nucleoprotein Interacts with Influenza Virus Polymerase Proteins. J. Virol. 1998, 72, 5493–5501. [Google Scholar] [CrossRef]
- Li, Z.; Watanabe, T.; Hatta, M.; Watanabe, S.; Nanbo, A.; Ozawa, M.; Kakugawa, S.; Shimojima, M.; Yamada, S.; Neumann, G.; et al. Mutational Analysis of Conserved Amino Acids in the Influenza A Virus Nucleoprotein. J. Virol. 2009, 83, 4153–4162. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.L.; Fan, H.; Bauer, D.L.V.; Grimes, J.M.; Fodor, E.; Keown, J.R. Structure of an H3N2 Influenza Virus Nucleoprotein. Acta Crystallogr. F Struct. Biol. Commun. 2021, 77, 208. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sneyd, H.; Dekant, R.; Wang, J. Influenza A Virus Nucleoprotein: A Highly Conserved Multi-Functional Viral Protein as a Hot Antiviral Drug Target. Curr. Top. Med. Chem. 2017, 17, 2271–2285. [Google Scholar] [CrossRef]
- Sun, W.; Luo, T.; Liu, W.; Li, J. Progress in the Development of Universal Influenza Vaccines. Viruses 2020, 12, 1033. [Google Scholar] [CrossRef]
- Pleguezuelos, O.; James, E.; Fernandez, A.; Lopes, V.; Rosas, L.A.; Cervantes-Medina, A.; Cleath, J.; Edwards, K.; Neitzey, D.; Gu, W.; et al. Efficacy of FLU-v, a Broad-Spectrum Influenza Vaccine, in a Randomized Phase IIb Human Influenza Challenge Study. NPJ Vaccines 2020, 5, 22. [Google Scholar] [CrossRef]
- Zhong, J.; Liang, L.; Huang, P.; Zhu, X.; Zou, L.; Yu, S.; Zhang, X.; Zhang, Y.; Ni, H.; Yan, J. Genetic Mutations in Influenza H3N2 Viruses from a 2012 Epidemic in Southern China. Virol. J. 2013, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Murcia, P.R.; Hughes, J.; Battista, P.; Lloyd, L.; Baillie, G.J.; Ramirez-Gonzalez, R.H.; Ormond, D.; Oliver, K.; Elton, D.; Mumford, J.A.; et al. Evolution of an Eurasian Avian-like Influenza Virus in Naïve and Vaccinated Pigs. PLoS Pathog. 2012, 8, e1002730. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.S.; Kelly, A.; Runstadler, J.A. Prevalence and Diversity of Avian Influenza Viruses in Environmental Reservoirs. J. Gen. Virol. 2008, 89, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Kalhoro, D.H.; Liang, S.; Kalhoro, M.S.; Pirzado, S.A.; Rajput, N.; Naeem, M.; Parveen, F.; Liu, Y. Identification and Genetic Evolution Analysis of One Strain of H3N2 Canine Influenza Virus Isolated from Nanjing, China. Pak. J. Zool. 2018, 50, 817–824. [Google Scholar] [CrossRef]
- Kirkpatrick, E.; Qiu, X.; Wilson, P.C.; Bahl, J.; Krammer, F. The Influenza Virus Hemagglutinin Head Evolves Faster than the Stalk Domain. Sci. Rep. 2018, 8, 10432. [Google Scholar] [CrossRef]
- Hussain, S.; Daniels, R.S.; Wharton, S.A.; Howell, S.; Halai, C.; Kunzelmann, S.; Whittaker, L.; McCauley, J.W. Reduced Sialidase Activity of Influenza A(H3N2) Neuraminidase Associated with Positively Charged Amino Acid Substitutions. J. Gen. Virol. 2021, 102, 001648. [Google Scholar] [CrossRef]
- Ge, J.; Lin, X.; Guo, J.; Liu, L.; Li, Z.; Lan, Y.; Liu, L.; Guo, J.; Lu, J.; Huang, W.; et al. The Antibody Response Against Neuraminidase in Human Influenza A (H3N2) Virus Infections During 2018/2019 Flu Season: Focusing on the Epitopes of 329-N-Glycosylation and E344 in N2. Front. Microbiol. 2022, 13, 590. [Google Scholar] [CrossRef]
- Korsun, N.; Trifonova, I.; Voleva, S.; Grigorova, I.; Angelova, S. Genetic Characterisation of the Influenza Viruses Circulating in Bulgaria during the 2019–2020 Winter Season. Virus Genes 2021, 57, 401–412. [Google Scholar] [CrossRef]
- Li, Z.; Zhong, L.; He, J.; Huang, Y.; Zhao, Y. Development and Application of Reverse Genetic Technology for the Influenza Virus. Virus Genes 2021, 57, 151. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HI Antibody Titer Against Challenge Strain in Sera | |||||
|---|---|---|---|---|---|
| Group | Pig ID | 42 dpv | Euthanasia Day | ||
| 2 dpi | 5 dpi | 9 dpi | |||
| Vaccinated animals | 1 | 10 | 40 | ||
| 2 | 40 | 320 | |||
| 3 | 20 | 160 | |||
| 4 | 80 | 80 | |||
| 5 | 80 | 160 | |||
| 6 | 20 | 80 | |||
| 7 | 40 | 160 | |||
| 8 | 40 | 80 | |||
| mean | 41.3 | 173.3 | 106.7 | 120 | |
| Nonvaccinated animals | 9 | 0 | 0 | ||
| 10 | 0 | 0 | |||
| 11 | 0 | 0 | |||
| 12 | 0 | 10 | |||
| 13 | 0 | 20 | |||
| 14 | 0 | 0 | |||
| 15 | 0 | 640 | |||
| 16 | 0 | 320 | |||
| mean | 0 | 0 | 10 | 480 | |
| Group | Pig ID | Nasal Swab Samples | Euthanized Day | Tissues Samples | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 dpi | 2 dpi | 3 dpi | 4 dpi | 5 dpi | 6 dpi | 7 dpi | 8 dpi | 9 dpi | LUNG | NT | BALF | |||
| Vaccinated animals | 1 | Neg. | Neg. | 2 dpi | 32.96 | Neg. | 34.45 | |||||||
| 2 | Neg. | Neg. | 34.48 | Neg. | Neg. | |||||||||
| 3 | Neg. | Neg. | 39.36 | Neg. | Neg. | |||||||||
| 4 | Neg. | Neg. | 33.45 | 32.56 | 37 | 5 dpi | Neg. | Neg. | 39.58 | |||||
| 5 | Neg. | Neg. | Neg. | Neg. | 37.42 | Neg. | Neg. | Neg. | ||||||
| 6 | Neg. | Neg. | Neg. | Neg. | Neg. | Neg. | Neg. | Neg. | ||||||
| 7 | Neg. | Neg. | 38,56 | Neg. | Neg. | Neg. | Neg. | Neg. | Neg. | 9 dpi | Neg. | Neg. | 38.09 | |
| 8 | Neg. | Neg. | Neg. | Neg. | Neg, | Neg, | Neg. | Neg. | 37.23 | Neg. | Neg. | Neg. | ||
| Nonvaccinated animals | 9 | Neg. | 31.21 | 2 dpi | 26.49 | Neg. | 35.98 | |||||||
| 10 | Neg. | 33.42 | 25.27 | 31.68 | 34.88 | |||||||||
| 11 | 39.48 | 33.39 | 33.34 | Neg. | 34.14 | |||||||||
| 12 | Neg. | 38.28 | 30.98 | Neg. | Neg. | 5 dpi | 34.41 | Neg. | Neg. | |||||
| 13 | 37.28 | 35 | 30.02 | 33.6 | 39.48 | 34.36 | 39.5 | 37.14 | ||||||
| 14 | Neg. | 38.75 | 36.96 | Neg. | Neg. | 35.38 | Neg. | Neg. | ||||||
| 15 | Neg. | 37.13 | 32.45 | 34.93 | Neg. | Neg. | Neg. | Neg. | Neg. | 39.78 | Neg. | Neg. | ||
| 16 | Neg. | Neg. | 32.8 | 30.17 | Neg. | 34.64 | Neg. | Neg. | Neg. | 9 dpi | 39.54 | Neg. | Neg. | |
| Group | Pig ID | Euthanasia Day | Lung Affected Area (%) | Histopathological Scoring | Immunohistochemistry for SIV |
|---|---|---|---|---|---|
| Vaccinated animals | 1 | 2 dpi | 1.44 | 2.5 | - |
| 2 | 0.32 | 2 | - | ||
| 3 | 0.32 | 2 | - | ||
| 4 | 5 dpi | 0.18 | 1.5 | - | |
| 5 | 0 | 0.5 | - | ||
| 6 | 10 | 2.5 | - | ||
| 7 | 9 dpi | 1.25 | 0.5 | - | |
| 8 | 0 | 0 | - | ||
| mean | 1.69 | 1.44 | |||
| Nonvaccinated animals | 9 | 2 dpi | 21.4 | 3 | ++ |
| 10 | 20.38 | 3 | +++ | ||
| 11 | 1.54 | 3 | - | ||
| 12 | 5 dpi | 7.3 | 2.5 | - | |
| 13 | 2.07 | 2 | + | ||
| 14 | 4.98 | 3 | - | ||
| 15 | 9 dpi | 1.11 | 1 | - | |
| 16 | 0.69 | 1.5 | - | ||
| mean | 7.43 | 2.38 |
| Gene | Depth of Read | Nucleotide Change | Alt. Base Count | Allele Frequency | Aminoacidic Change | |||
|---|---|---|---|---|---|---|---|---|
| Position | Ref. → Alt. | Ref. → Alt. | Position | |||||
| Inoculum | PA | 503 | G → A | 163 | 15 | 2.99 | D → N | 55 |
| 147 | A → C | 1850 | 28 | 19.05 | E → A | 617 | ||
| 160 | G → A | 1852 | 28 | 17.5 | V → I | 618 | ||
| 1375 | C → A | 2047 | 70 | 5.09 | L → I | 683 | ||
| PB2 | 863 | C → A | 217 | 20 | 2.32 | Q → K | 73 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Valiñas, Á.; Baioni, L.; Córdoba, L.; Darji, A.; Chiapponi, C.; Segalés, J.; Ganges, L.; Núñez, J.I. Evolution of Swine Influenza Virus H3N2 in Vaccinated and Nonvaccinated Pigs after Previous Natural H1N1 Infection. Viruses 2022, 14, 2008. https://doi.org/10.3390/v14092008
López-Valiñas Á, Baioni L, Córdoba L, Darji A, Chiapponi C, Segalés J, Ganges L, Núñez JI. Evolution of Swine Influenza Virus H3N2 in Vaccinated and Nonvaccinated Pigs after Previous Natural H1N1 Infection. Viruses. 2022; 14(9):2008. https://doi.org/10.3390/v14092008
Chicago/Turabian StyleLópez-Valiñas, Álvaro, Laura Baioni, Lorena Córdoba, Ayub Darji, Chiara Chiapponi, Joaquim Segalés, Llilianne Ganges, and José I. Núñez. 2022. "Evolution of Swine Influenza Virus H3N2 in Vaccinated and Nonvaccinated Pigs after Previous Natural H1N1 Infection" Viruses 14, no. 9: 2008. https://doi.org/10.3390/v14092008
APA StyleLópez-Valiñas, Á., Baioni, L., Córdoba, L., Darji, A., Chiapponi, C., Segalés, J., Ganges, L., & Núñez, J. I. (2022). Evolution of Swine Influenza Virus H3N2 in Vaccinated and Nonvaccinated Pigs after Previous Natural H1N1 Infection. Viruses, 14(9), 2008. https://doi.org/10.3390/v14092008