
Construction and Evaluation of Recombinant Adenovirus Candidate Vaccines for Chikungunya Virus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Virus, and Animals
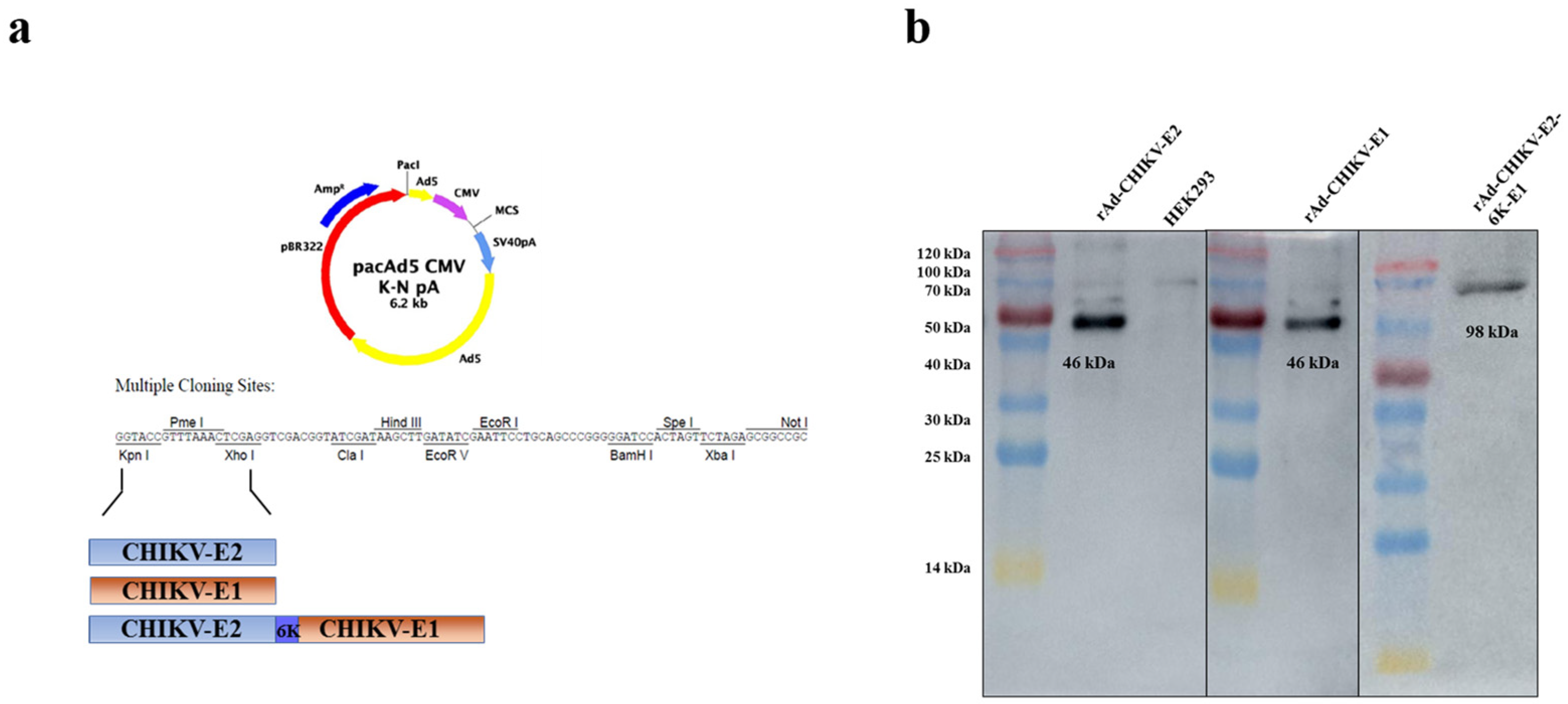
2.2. Construction and Harvest of Recombinant Viruses
2.3. Expression of E2, E1, E2-6K-E1 Glycoprotein of CHIKV
2.4. Immunization Schedule of Recombinant Vaccines in Mice
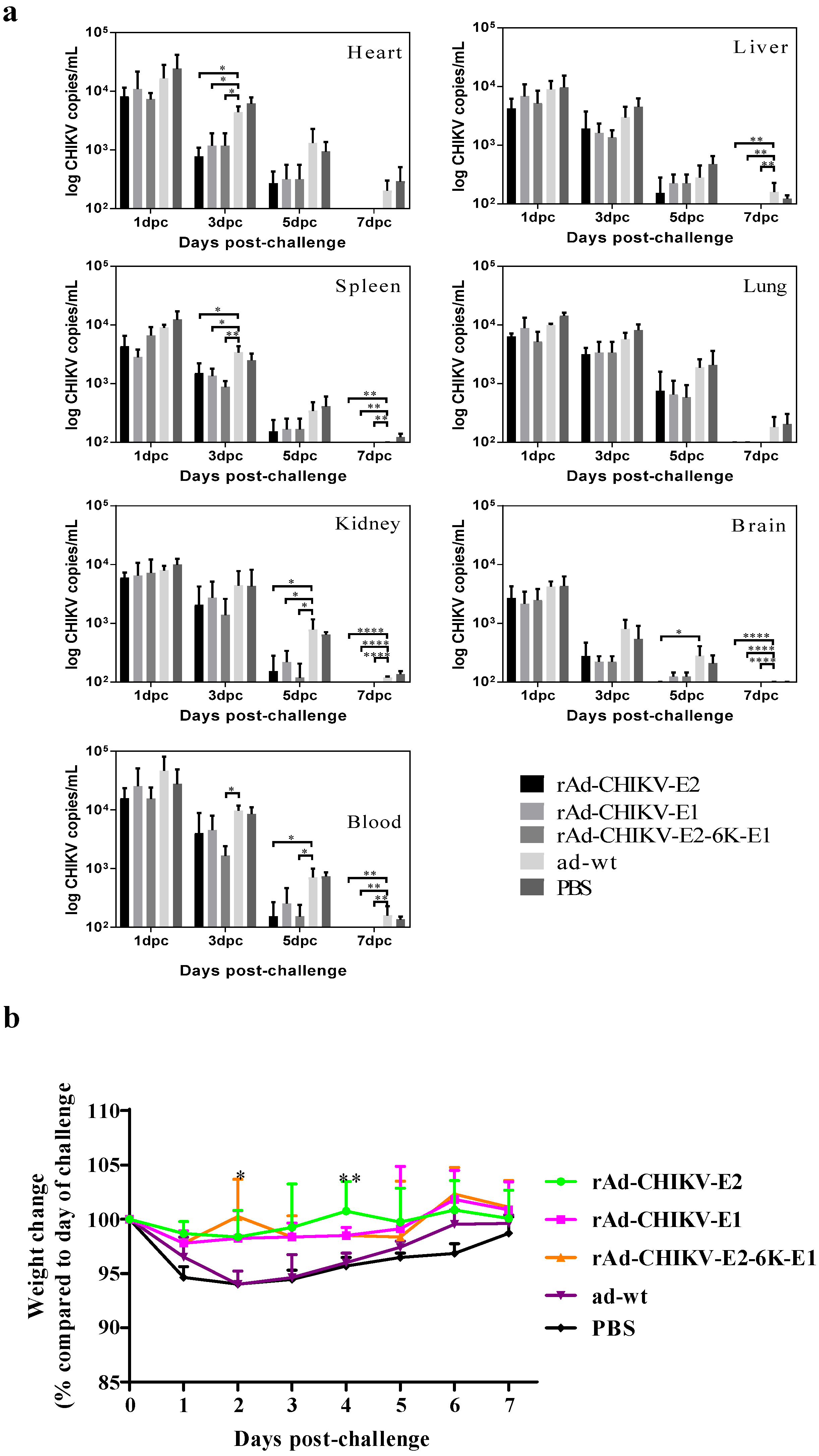
2.5. Challenge Assay of Mice
2.6. Quantitative RT-PCR
2.7. Evaluation of Humoral Immune Responses
2.7.1. Determination of Antibody Titers
2.7.2. Detection of Neutralization Antibodies
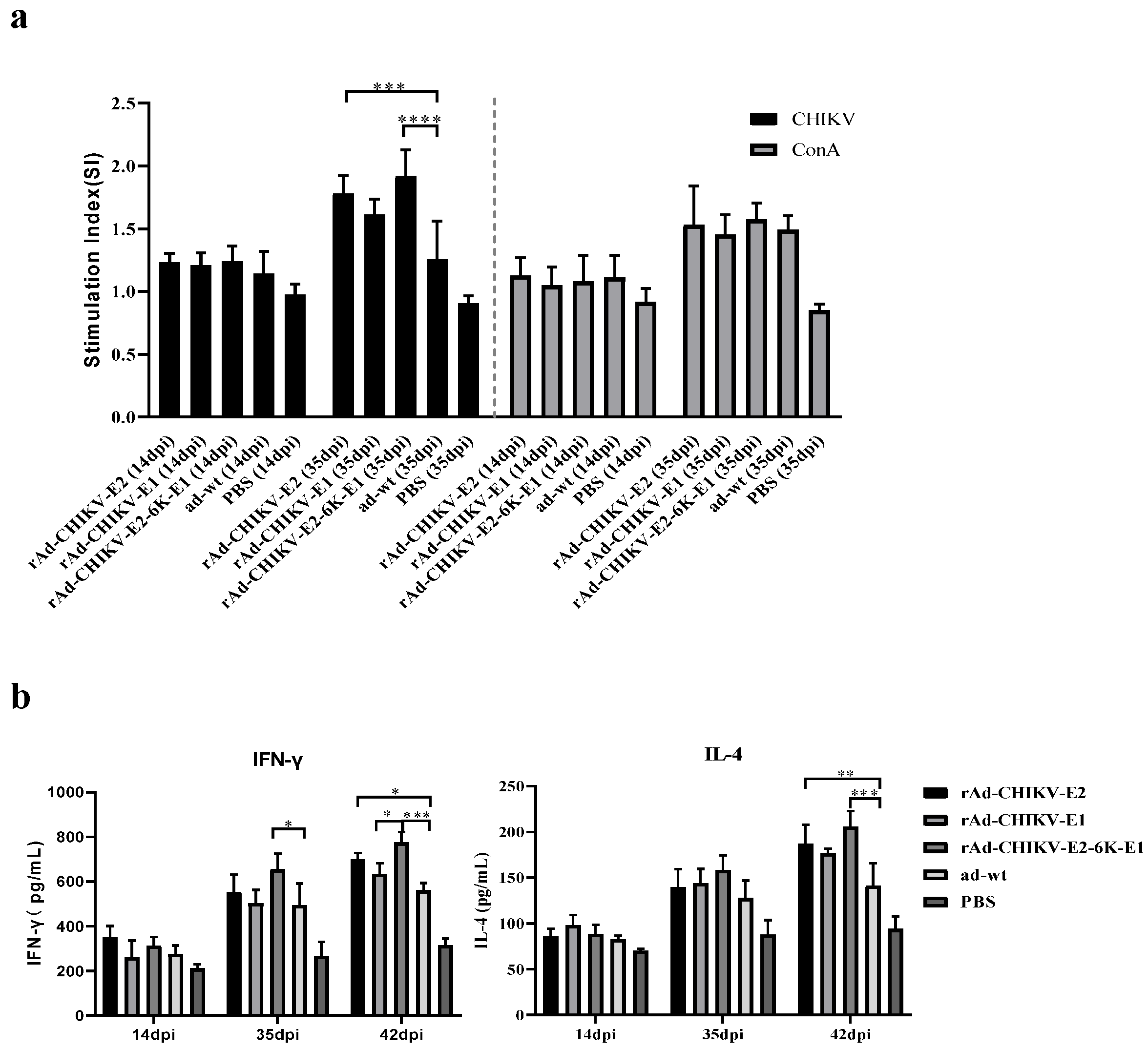
2.8. Assessment of Cellular Immune Responses
2.8.1. Measurement of Cytokines
2.8.2. Lymphocytes Proliferation Assay
2.9. Statistical Analysis
3. Results
3.1. Construction and Identification of Recombinant Viruses
3.2. CHIKV Recombinant Adenovirus Elicits Higher Humoral Immune Responses
3.3. CHIKV Recombinant Adenovirus Vaccines Balances Th1/Th2 Responses
3.4. Protective Efficacy of Recombinant Adenovirus Vaccines against Viral Challenge In Vivo
4. Discussion
5. Conclusions
Authors Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Ross, R.W. The Newala epidemic. III. The virus: Isolation, pathogenic properties and relationship to the epidemic. J. Hyg. 1956, 54, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Staples, J.E.; Breiman, R.F.; Powers, A.M. Chikungunya fever: An epidemiological review of a re-emerging infectious disease. Clin. Infect. Dis. 2009, 49, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, W.H. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952-53. II. General description and epidemiology. Trans. R. Soc. Trop. Med. Hyg. 1955, 49, 33–57. [Google Scholar] [CrossRef]
- Enserink, M. Infectious diseases. Chikungunya: No longer a third world disease. Science 2007, 318, 1860–1861. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.H.; Morita, K.; Parquet Md Mdel, C.; Hasebe, F.; Mathenge, E.G.; Igarashi, A. Complete nucleotide sequence of chikungunya virus and evidence for an internal polyadenylation site. J. Gen. Virol. 2002, 83 Pt 12, 3075–3084. [Google Scholar] [CrossRef] [PubMed]
- Goh, L.Y.; Hobson-Peters, J.; Prow, N.A.; Baker, K.; Piyasena, T.B.; Taylor, C.T.; Rana, A.; Hastie, M.L.; Gorman, J.J.; Hall, R.A. The Chikungunya Virus Capsid Protein Contains Linear B Cell Epitopes in the N- and C-Terminal Regions that are Dependent on an Intact C-Terminus for Antibody Recognition. Viruses 2015, 7, 2943–2964. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jose, J.; Xiang, Y.; Kuhn, R.J.; Rossmann, M.G. Structural changes of envelope proteins during alphavirus fusion. Nature 2010, 468, 705–708. [Google Scholar] [CrossRef]
- Weber, C.; Buchner, S.M.; Schnierle, B.S. A small antigenic determinant of the Chikungunya virus E2 protein is sufficient to induce neutralizing antibodies which are partially protective in mice. PLoS Negl. Trop. Dis. 2015, 9, e0003684. [Google Scholar] [CrossRef]
- Sanchez-San Martin, C.; Nanda, S.; Zheng, Y.; Fields, W.; Kielian, M. Cross-inhibition of chikungunya virus fusion and infection by alphavirus E1 domain III proteins. J. Virol. 2013, 87, 7680–7687. [Google Scholar] [CrossRef]
- Chung, N.H.; Chen, Y.C.; Yang, S.J.; Lin, Y.C.; Dou, H.Y.; Hui-Ching Wang, L.; Liao, C.L.; Chow, Y.H. Induction of Th1 and Th2 in the protection against SARS-CoV-2 through mucosal delivery of an adenovirus vaccine expressing an engineered spike protein. Vaccine 2022, 40, 574–586. [Google Scholar] [CrossRef]
- Dora, E.G.; Rossi, S.L.; Weaver, S.C.; Tucker, S.N.; Mateo, R. An adjuvanted adenovirus 5-based vaccine elicits neutralizing antibodies and protects mice against chikungunya virus-induced footpad swelling. Vaccine 2019, 37, 3146–3150. [Google Scholar] [CrossRef]
- Zhang, H.; Nan, F.; Li, Z.; Zhao, G.; Xie, C.; Ha, Z.; Zhang, J.; Han, J.; Xiao, P.; Zhuang, X.; et al. Construction and immunological evaluation of recombinant Newcastle disease virus vaccines expressing highly pathogenic porcine reproductive and respiratory syndrome virus GP3/GP5 proteins in pigs. Vet. Microbiol. 2019, 239, 108490. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Dash, P.K.; Santhosh, S.R.; Shukla, J.; Parida, M.; Rao, P.V. Development of a quantitative competitive reverse transcription polymerase chain reaction (QC-RT-PCR) for detection and quantitation of Chikungunya virus. Mol. Biotechnol. 2010, 45, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Weger-Lucarelli, J.; Chu, H.; Aliota, M.T.; Partidos, C.D.; Osorio, J.E. A novel MVA vectored Chikungunya virus vaccine elicits protective immunity in mice. PLoS Negl. Trop. Dis. 2014, 8, e2970. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Krause, K.K.; Azouz, F.; Nakano, E.; Nerurkar, V.R. A guinea pig model of Zika virus infection. Virol. J. 2017, 14, 75. [Google Scholar] [CrossRef] [PubMed]
- Edelman, R.; Tacket, C.O.; Wasserman, S.S.; Bodison, S.A.; Perry, J.G.; Mangiafico, J.A. Phase II safety and immunogenicity study of live chikungunya virus vaccine TSI-GSD-218. Am. J. Trop. Med. Hyg. 2000, 62, 681–685. [Google Scholar] [CrossRef]
- Muthumani, K.; Lankaraman, K.M.; Laddy, D.J.; Sundaram, S.G.; Chung, C.W.; Sako, E.; Wu, L.; Khan, A.; Sardesai, N.; Kim, J.J.; et al. Immunogenicity of novel consensus-based DNA vaccines against Chikungunya virus. Vaccine 2008, 26, 5128–5134. [Google Scholar] [CrossRef]
- Wang, D.; Suhrbier, A.; Penn-Nicholson, A.; Woraratanadharm, J.; Gardner, J.; Luo, M.; Le, T.T.; Anraku, I.; Sakalian, M.; Einfeld, D.; et al. A complex adenovirus vaccine against chikungunya virus provides complete protection against viraemia and arthritis. Vaccine 2011, 29, 2803–2809. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Wang, E.; Seymour, R.; Weaver, S.C.; Rose, J.K. A chimeric vesiculo/alphavirus is an effective alphavirus vaccine. J. Virol. 2013, 87, 395–402. [Google Scholar] [CrossRef]
- Chang, L.J.; Dowd, K.A.; Mendoza, F.H.; Saunders, J.G.; Sitar, S.; Plummer, S.H.; Yamshchikov, G.; Sarwar, U.N.; Hu, Z.; Enama, M.E.; et al. Safety and tolerability of chikungunya virus-like particle vaccine in healthy adults: A phase 1 dose-escalation trial. Lancet 2014, 384, 2046–2052. [Google Scholar] [CrossRef]
- Voss, J.E.; Vaney, M.C.; Duquerroy, S.; Vonrhein, C.; Girard-Blanc, C.; Crublet, E.; Thompson, A.; Bricogne, G.; Rey, F.A. Glycoprotein organization of Chikungunya virus particles revealed by X-ray crystallography. Nature 2010, 468, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Powers, A.M. Vaccine and Therapeutic Options to Control Chikungunya Virus. Clin. Microbiol. Rev. 2018, 31, e00104-16. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, I.S.B.; Tanabe, E.L.L.; Santos, E.C.; Martins, W.V.; Araujo, I.; Cavalcante, M.C.A.; Lima, A.R.V.; Camara, N.O.S.; Anderson, L.; Yunusov, D.; et al. Cellular and Molecular Immune Response to Chikungunya Virus Infection. Front. Cell Infect. Microbiol. 2018, 8, 345. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Liss, N.M.; Chen, D.H.; Liao, M.; Fox, J.M.; Shimak, R.M.; Fong, R.H.; Chafets, D.; Bakkour, S.; Keating, S.; et al. Neutralizing Monoclonal Antibodies Block Chikungunya Virus Entry and Release by Targeting an Epitope Critical to Viral Pathogenesis. Cell Rep. 2015, 13, 2553–2564. [Google Scholar] [CrossRef] [PubMed]
- Hallengard, D.; Kakoulidou, M.; Lulla, A.; Kummerer, B.M.; Johansson, D.X.; Mutso, M.; Lulla, V.; Fazakerley, J.K.; Roques, P.; Le Grand, R.; et al. Novel attenuated Chikungunya vaccine candidates elicit protective immunity in C57BL/6 mice. J. Virol. 2014, 88, 2858–2866. [Google Scholar] [CrossRef]
- McConnell, M.J.; Imperiale, M.J. Biology of adenovirus and its use as a vector for gene therapy. Hum. Gene Ther. 2004, 15, 1022–1033. [Google Scholar] [CrossRef]
- Garcia-Arriaza, J.; Cepeda, V.; Hallengard, D.; Sorzano, C.O.; Kummerer, B.M.; Liljestrom, P.; Esteban, M. A novel poxvirus-based vaccine, MVA-CHIKV, is highly immunogenic and protects mice against chikungunya infection. J. Virol. 2014, 88, 3527–3547. [Google Scholar] [CrossRef]
- Khan, M.; Dhanwani, R.; Rao, P.V.; Parida, M. Subunit vaccine formulations based on recombinant envelope proteins of Chikungunya virus elicit balanced Th1/Th2 response and virus-neutralizing antibodies in mice. Virus Res. 2012, 167, 236–246. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Shukarev, G.; Callendret, B.; Luhn, K.; Douoguih, M.; Consortium, E. A two-dose heterologous prime-boost vaccine regimen eliciting sustained immune responses to Ebola Zaire could support a preventive strategy for future outbreaks. Hum. Vaccines Immunother. 2017, 13, 266–270. [Google Scholar] [CrossRef]
- Baden, L.R.; Karita, E.; Mutua, G.; Bekker, L.G.; Gray, G.; Page-Shipp, L.; Walsh, S.R.; Nyombayire, J.; Anzala, O.; Roux, S.; et al. Assessment of the Safety and Immunogenicity of 2 Novel Vaccine Platforms for HIV-1 Prevention: A Randomized Trial. Ann. Intern. Med. 2016, 164, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Sudeep, A.B.; Arankalle, V.A. Evaluation of recombinant E2 protein-based and whole-virus inactivated candidate vaccines against chikungunya virus. Vaccine 2012, 30, 6142–6149. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Injection | Injection Route | Injection Dose | Challenge | Challenge Route | Challenge Dose |
|---|---|---|---|---|---|---|
| I | rAd-CHIKV-E2 | i.m. | 109 pfu/100 μL | CHIKV | i.v. | 104 pfu/100 μL |
| II | rAd-CHIKV-E1 | i.m. | 109 pfu/100 μL | CHIKV | i.v. | 104 pfu/100 μL |
| III | rAd-CHIKV-E2-6K-E1 | i.m. | 109 pfu/100 μL | CHIKV | i.v. | 104 pfu/100 μL |
| IV | ad-wt | i.m. | 109 pfu/100 μL | CHIKV | i.v. | 104 pfu/100 μL |
| V | PBS | i.m. | 100 μL | CHIKV | i.v. | 104 pfu/100 μL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.; Wang, W.; Sun, W.; Zhang, J.; Han, J.; Xie, C.; Ha, Z.; Xie, Y.; Zhang, H.; Jin, N.; et al. Construction and Evaluation of Recombinant Adenovirus Candidate Vaccines for Chikungunya Virus. Viruses 2022, 14, 1779. https://doi.org/10.3390/v14081779
Cao L, Wang W, Sun W, Zhang J, Han J, Xie C, Ha Z, Xie Y, Zhang H, Jin N, et al. Construction and Evaluation of Recombinant Adenovirus Candidate Vaccines for Chikungunya Virus. Viruses. 2022; 14(8):1779. https://doi.org/10.3390/v14081779
Chicago/Turabian StyleCao, Liang, Wei Wang, Wenchao Sun, Jinyong Zhang, Jicheng Han, Changzhan Xie, Zhuo Ha, Yubiao Xie, He Zhang, Ningyi Jin, and et al. 2022. "Construction and Evaluation of Recombinant Adenovirus Candidate Vaccines for Chikungunya Virus" Viruses 14, no. 8: 1779. https://doi.org/10.3390/v14081779
APA StyleCao, L., Wang, W., Sun, W., Zhang, J., Han, J., Xie, C., Ha, Z., Xie, Y., Zhang, H., Jin, N., & Lu, H. (2022). Construction and Evaluation of Recombinant Adenovirus Candidate Vaccines for Chikungunya Virus. Viruses, 14(8), 1779. https://doi.org/10.3390/v14081779