
A Novel Competitive ELISA for Specifically Measuring and Differentiating Immune Responses to Classical Swine Fever C-Strain Vaccine in Pigs

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Lines and Media
2.3. Serum Samples
2.4. Expression of C-Strain Erns Protein and Generation of Monoclonal Antibodies
2.5. SDS-PAGE and Western Blotting
2.6. Competitive Enzyme-Linked Immunosorbent Assay (cELISA)
2.7. Reproducibility and Statistical Analysis
3. Results
3.1. Generation of Capture Antigen and Suitable Competitive Monoclonal Antibody
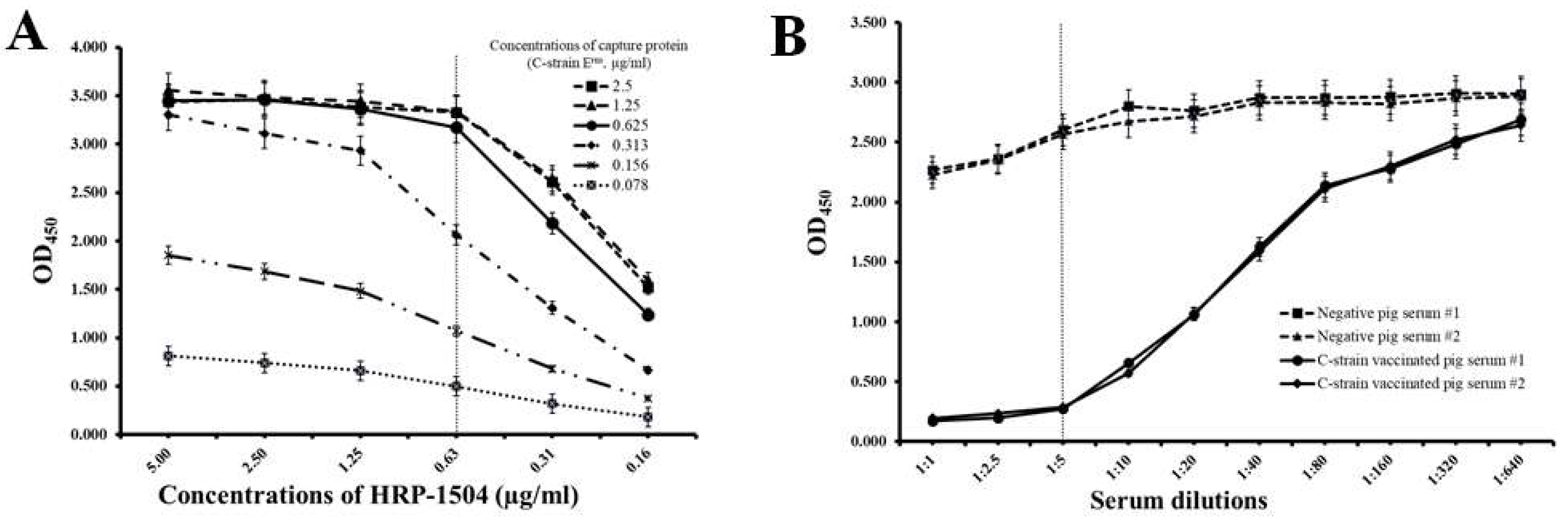
3.2. Establishment of Competitive ELISA System with C-Strain Erns Protein and mAb 1504
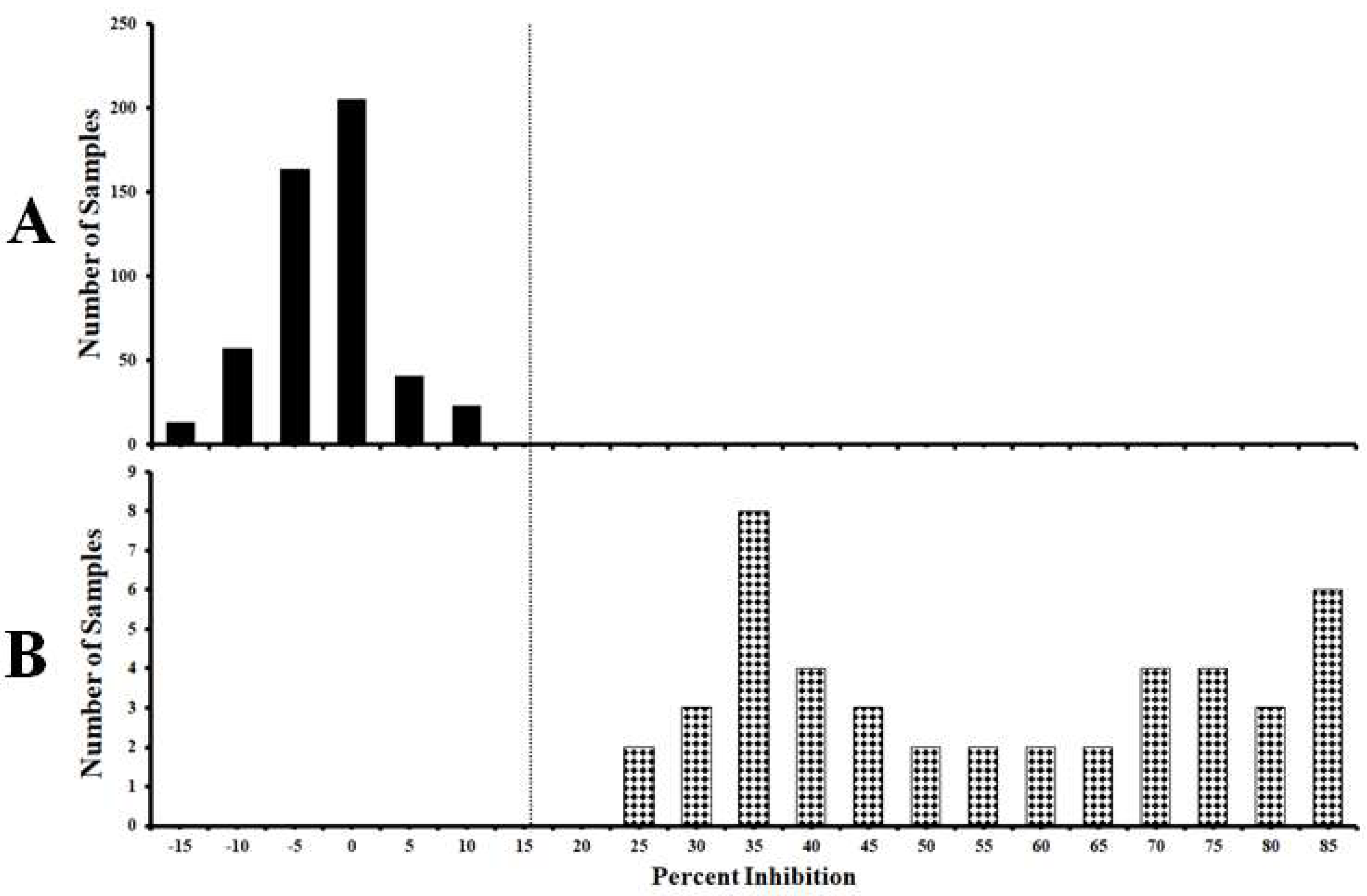
3.3. Standardization of the Cut-Off Value of the Established cELISA
3.4. The Established cELISA Can Efficiently Differentiate C-Strain/C-Strain Erns Immunized Pigs from Wild-Type CSFV and Other Common Swine Virus-Infected Pigs
3.5. Reproducibility of the cELISA
3.6. Kinetics of Erns Antibody Response of C-Strain-Vaccinated Pigs at Different Time Intervals
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moennig, V. Introduction to classical swine fever: Virus, disease and control policy. Vet. Microbiol. 2000, 73, 93–102. [Google Scholar] [CrossRef]
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Madera, R.; Li, Y.; McVey, D.S.; Drolet, B.S.; Shi, J. Recent Advances in the Diagnosis of Classical Swine Fever and Future Perspectives. Pathogens 2020, 9, 658. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, L.; McVey, D.S. Of pigs and men: The best-laid plans for prevention and control of swine fevers. Anim. Front. 2021, 11, 6–13. [Google Scholar] [CrossRef]
- van Oirschot, J.T. Vaccinology of classical swine fever: From lab to field. Vet. Microbiol. 2003, 96, 367–384. [Google Scholar] [CrossRef]
- Luo, Y.; Li, S.; Sun, Y.; Qiu, H.J. Classical swine fever in China: A minireview. Vet. Microbiol. 2014, 172, 1–6. [Google Scholar] [CrossRef]
- Blome, S.; Moß, C.; Reimann, I.; König, P.; Beer, M. Classical swine fever vaccines: State-of-the-art. Vet. Microbiol. 2017, 206, 10–20. [Google Scholar] [CrossRef]
- Postel, A.; Austermann-Busch, S.; Petrov, A.; Moennig, V.; Becher, P. Epidemiology, diagnosis and control of classical swine fever: Recent developments and future challenges. Transbound. Emerg. Dis. 2018, 65, 248–261. [Google Scholar] [CrossRef]
- Coronado, L.; Perera, C.L.; Rios, L.; Frías, M.T.; Pérez, L.J. A Critical Review about Different Vaccines against Classical Swine Fever Virus and Their Repercussions in Endemic Regions. Vaccines 2021, 9, 154. [Google Scholar] [CrossRef]
- Paton, D.J.; McGoldrick, A.; Greiser-Wilke, I.; Parchariyanon, S.; Song, J.Y.; Liou, P.P.; Stadejek, T.; Lowings, J.P.; Björklund, H.; Belák, S. Genetic typing of classical swine fever virus. Vet. Microbiol. 2000, 73, 137–157. [Google Scholar] [CrossRef]
- Postel, A.; Schmeiser, S.P.; Perera, C.L.; Rodríguez, L.J.; Frias-Lepoureau, M.T.; Becher, P. Classical swine fever virus isolates from Cuba form a new subgenotype 1.4. Vet. Microbiol. 2013, 161, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Lv, L.; Gu, J.; Chen, T.; Xiao, Y.; Liu, S. Genetic diversity and positive selection analysis of classical swine fever virus envelope protein gene E2 in east China under C-strain vaccination. Front. Microbiol. 2016, 7, 85. [Google Scholar] [CrossRef] [PubMed][Green Version]
- König, M.; Lengsfeld, T.; Pauly, T.; Stark, R.; Thiel, H.J. Classical swine fever virus: Independent induction of protective immunity by two structural glycoproteins. J. Virol. 1995, 69, 6479–6486. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Bai, Y.; Song, Y.; Liu, Y.; Yu, W.; Sun, Y.; Wang, L.; Deng, R.; Xing, G.; Zhang, G. Generation and immunogenicity analysis of recombinant classical swine fever virus glycoprotein E2 and Erns expressed in baculovirus expression system. Virol. J. 2021, 18, 44. [Google Scholar] [CrossRef]
- Moormann, R.J.; Bouma, A.; Kramps, J.A.; Terpstra, C.; De Smit, H.J. Development of a classical swine fever subunit marker vaccine and companion diagnostic test. Vet. Microbiol. 2000, 73, 209–219. [Google Scholar] [CrossRef]
- Langedijk, J.P.; Middel, W.G.; Meloen, R.H.; Kramps, J.A.; de Smit, J.A. Enzyme-linked immunosorbent assay using a virus type-specific peptide based on a subdomain of envelope protein Erns for serologic diagnosis of pestivirus infections in swine. J. Clin. Microbiol. 2001, 39, 906–912. [Google Scholar] [CrossRef]
- Lin, M.; Trottier, E.; Pasick, J. Antibody responses of pigs to defined Erns fragments after infection with classical swine fever virus. Clin. Diagn. Lab. Immunol. 2005, 12, 180–186. [Google Scholar]
- Schroeder, S.; von Rosen, T.; Blome, S.; Loeffen, W.; Haegeman, A.; Koenen, F.; Uttenthal, A. Evaluation of classical swine fever virus antibody detection assays with an emphasis on the differentiation of infected from vaccinated animals. Rev. Sci. Tech. 2012, 31, 997–1010. [Google Scholar] [CrossRef]
- Pannhorst, K.; Fröhlich, A.; Staubach, C.; Meyer, D.; Blome, S.; Becher, P. Evaluation of an Erns-based enzyme-linked immunosorbent assay to distinguish Classical swine fever virus-infected pigs from pigs vaccinated with CP7_E2alf. J. Vet. Diagn. Investig. 2015, 27, 449–460. [Google Scholar] [CrossRef]
- Madera, R.; Gong, W.; Wang, L.; Burakova, Y.; Lleellish, K.; Galliher-Beckley, A.; Nietfeld, J.; Henningson, J.; Jia, K.; Li, P.; et al. Pigs immunized with a novel E2 subunit vaccine are protected from subgenotype heterologous classical swine fever virus challenge. BMC Vet. Res. 2016, 12, 197. [Google Scholar] [CrossRef]
- Wang, L.; Mi, S.; Madera, R.; Ganges, L.; Borca, M.V.; Ren, J.; Cunningham, C.; Cino-Ozuna, A.G.; Li, H.; Tu, C.; et al. A neutralizing monoclonal antibody-based competitive ELISA for classical swine fever C-strain post-vaccination monitoring. BMC Vet. Res. 2020, 16, 14. [Google Scholar]
- Postel, A.; Moennig, V.; Becher, P. Classical swine fever in Europe—The current situation. Berl. Munch. Tierarztl. Wochenschr. 2013, 126, 468–475. [Google Scholar] [PubMed]
- Postel, A.; Nishi, T.; Kameyama, K.I.; Meyer, D.; Suckstorff, O.; Fukai, K.; Becher, P. Reemergence of Classical Swine Fever, Japan, 2018. Emerg. Infect. Dis. 2019, 25, 1228–1231. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Villamandos, J.C.; Carrasco, L.; Bautista, M.J.; Sierra, M.A.; Quezada, M.; Hervas, J.; Chacón, M.; Ruiz-Villamor, E.; Salguero, F.J.; Sónchez-Cordón, P.J.; et al. African swine fever and classical swine fever: A review of the pathogenesis. Dtsch. Tierarztl. Wochenschr. 2003, 110, 165–169. [Google Scholar]
- Mi, S.; Wang, L.; Li, H.; Fei, B.; Madera, R.; Shi, X.; Zhang, L.; Mao, Y.; Yan, X.; Xia, X.; et al. Characterization of monoclonal antibodies that specifically differentiate field isolates from vaccine strains of classical swine fever virus. Front. Immunol. 2022; accepted. [Google Scholar]
- Blome, S.; Wernike, K.; Reimann, I.; König, P.; Moß, C.; Beer, M. A decade of research into classical swine fever marker vaccine CP7_E2alf (Suvaxyn® CSF Marker): A review of vaccine properties. Vet. Res. 2017, 48, 51. [Google Scholar] [CrossRef] [PubMed]
- Jelsma, T.; Post, J.; Born, E.; Segers, R.; Kortekaas, J. Assessing the Protective Dose of a Candidate DIVA Vaccine against Classical Swine Fever. Vaccines 2021, 9, 483. [Google Scholar] [CrossRef]
- Widjojoatmodjo, M.N.; van Gennip, H.G.; Bouma, A.; van Rijn, P.A.; Moormann, R.J. Classical swine fever virus Erns deletion mutants: Trans-complementation and potential use as nontransmissible, modified, live-attenuated marker vaccines. J. Virol. 2000, 74, 2973–2980. [Google Scholar] [CrossRef]
- van Gennip, H.G.; Bouma, A.; van Rijn, P.A.; Widjojoatmodjo, M.N.; Moormann, R.J. Experimental non-transmissible marker vaccines for classical swine fever (CSF) by trans-complementation of Erns or E2 of CSFV. Vaccine 2002, 20, 1544–1556. [Google Scholar] [CrossRef]
- Greiser-Wilke, I.; Blome, S.; Moennig, V. Diagnostic methods for detection of Classical swine fever virus-status quo and new developments. Vaccine 2007, 25, 5524–5530. [Google Scholar] [CrossRef]
- OIE Terrestrial Manual Chapter 2.2.1. Available online: https://www.oie.int/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access (accessed on 11 February 2022).
- Li, Y.; Zhao, J.J.; Li, N.; Shi, Z.; Cheng, D.; Zhu, Q.H.; Tu, C.; Tong, G.Z.; Qiu, H.J. A multiplex nested RT-PCR for the detection and differentiation of wild-type viruses from C-strain vaccine of classical swine fever virus. J. Virol. Methods 2007, 143, 16–22. [Google Scholar] [CrossRef]
- Leifer, I.; Depner, K.; Blome, S.; Le Potier, M.F.; Le Dimna, M.; Beer, M.; Hoffmann, B. Differentiation of C-strain “Riems” or CP7_E2alf vaccinated animals from animals infected by classical swine fever virus field strains using real-time RT-PCR. J. Virol. Methods 2009, 158, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xia, H.; Everett, H.; Sosan, O.; Crooke, H.; Meindl-Böhmer, A.; Qiu, H.J.; Moennig, V.; Belák, S.; Widén, F. A generic real-time TaqMan assay for specific detection of lapinized Chinese vaccines against classical swine fever. J. Virol. Methods 2011, 175, 170–174. [Google Scholar] [CrossRef]
- Zhang, X.J.; Han, Q.Y.; Sun, Y.; Zhang, X.; Qiu, H.J. Development of a triplex TaqMan real-time RT-PCR assay for differential detection of wild-type and HCLV vaccine strains of classical swine fever virus and bovine viral diarrhea virus 1. Res. Vet. Sci. 2012, 92, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Everett, H.E.; Crudgington, B.S.; Sosan-Soulé, O.; Crooke, H.R. Differential detection of classical swine fever virus challenge strains in C-strain vaccinated pigs. BMC Vet. Res. 2014, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Widén, F.; Everett, H.; Blome, S.; Fernandez Pinero, J.; Uttenthal, A.; Cortey, M.; von Rosen, T.; Tignon, M.; Liu, L. Comparison of two real-time RT-PCR assays for differentiation of C-strain vaccinated from classical swine fever infected pigs and wild boars. Res. Vet. Sci. 2014, 97, 455–457. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibition Range (%) | No. of Serum Tested | CV Range (%) | |
|---|---|---|---|
| Intra-Assay | Inter-Assay | ||
| 65–80 | 20 | 0.05–1.37 | 0.86–8.50 |
| <8 | 20 | 0.10–0.45 | 0.56–5.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Mi, S.; Madera, R.; Li, Y.; Gong, W.; Tu, C.; Shi, J. A Novel Competitive ELISA for Specifically Measuring and Differentiating Immune Responses to Classical Swine Fever C-Strain Vaccine in Pigs. Viruses 2022, 14, 1544. https://doi.org/10.3390/v14071544
Wang L, Mi S, Madera R, Li Y, Gong W, Tu C, Shi J. A Novel Competitive ELISA for Specifically Measuring and Differentiating Immune Responses to Classical Swine Fever C-Strain Vaccine in Pigs. Viruses. 2022; 14(7):1544. https://doi.org/10.3390/v14071544
Chicago/Turabian StyleWang, Lihua, Shijiang Mi, Rachel Madera, Yuzhen Li, Wenjie Gong, Changchun Tu, and Jishu Shi. 2022. "A Novel Competitive ELISA for Specifically Measuring and Differentiating Immune Responses to Classical Swine Fever C-Strain Vaccine in Pigs" Viruses 14, no. 7: 1544. https://doi.org/10.3390/v14071544
APA StyleWang, L., Mi, S., Madera, R., Li, Y., Gong, W., Tu, C., & Shi, J. (2022). A Novel Competitive ELISA for Specifically Measuring and Differentiating Immune Responses to Classical Swine Fever C-Strain Vaccine in Pigs. Viruses, 14(7), 1544. https://doi.org/10.3390/v14071544