
Persistent, and Asymptomatic Viral Infections and Whitefly-Transmitted Viruses Impacting Cantaloupe and Watermelon in Georgia, USA

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
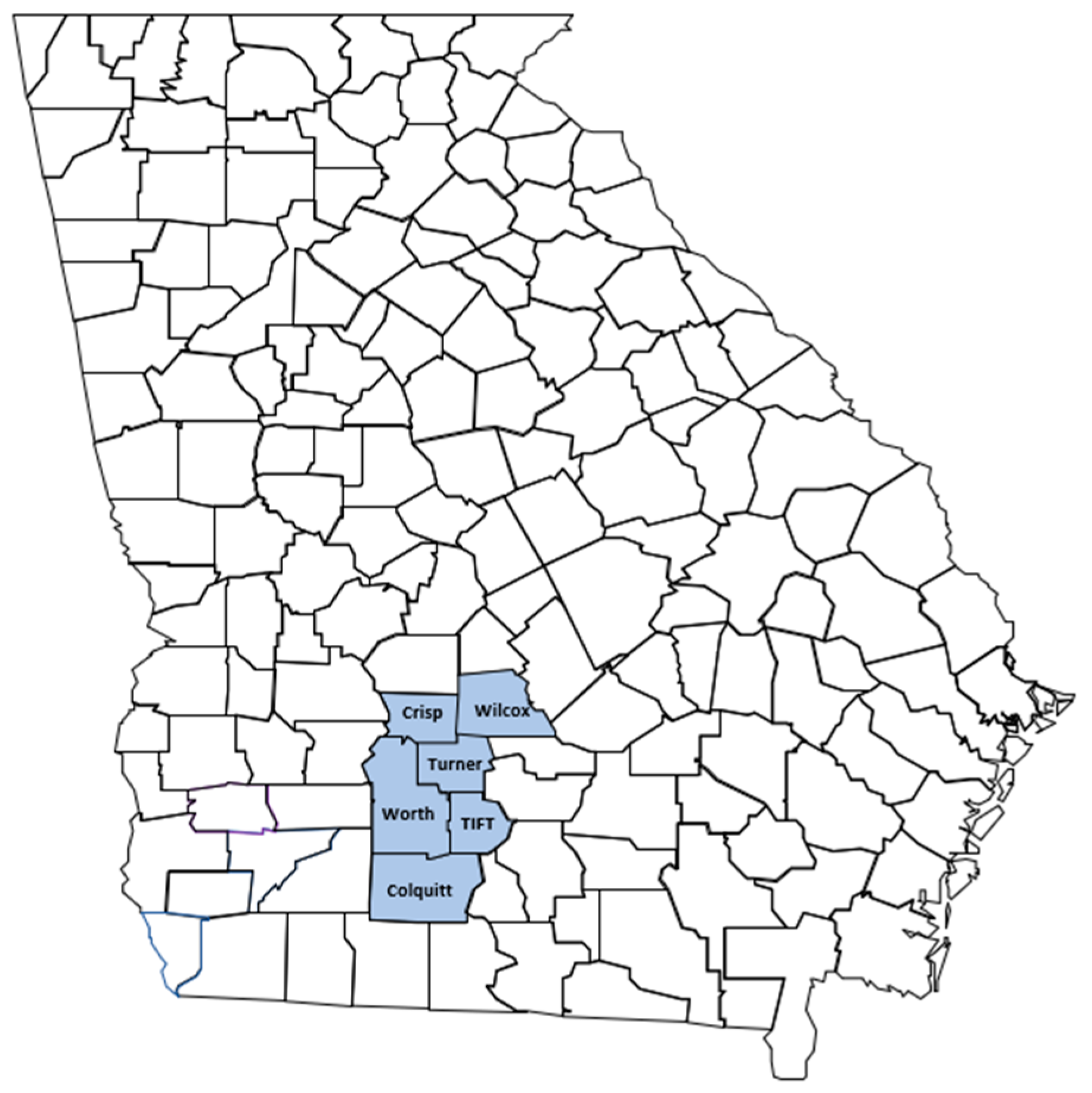
2.1. Survey Route and Sample Collection
2.2. RNA Extraction and HTS of Small RNA
2.3. Analysis of HTS Data
2.4. Total Nucleic Acid Extraction and Virus Detection by PCR and RT-PCR
2.5. Construction of Consensus Viral Genome Sequences
3. Results
3.1. Symptoms
3.2. Viruses Detected by HTS and Their Characteristics
3.3. Prevalence and Distribution of the Viruses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Georgia Farm Gate Value Report 2019. Available online: https://caed.uga.edu/publications/georgia-agricultural-statistics.html (accessed on 24 April 2022).
- Larsen, R. First report of cucurbit leaf crumple virus in snap bean in Georgia. Phytopathology 2010, 100, S68. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Dutta, B.; Coolong, T.; Sparks, A.N.; Adkins, S.; Srinivasan, R. First report of cucurbit yellow stunting disorder virus in cucurbits in Georgia, United States. Plant Health Prog. 2018, 19, 9–10. [Google Scholar] [CrossRef] [Green Version]
- Kavalappara, S.R.; Milner, H.; Sparks, A.N.; McGregor, C.; Wintermantel, W.M.; Bag, S. First report of cucurbit chlorotic yellows virus in association with other whitefly-transmitted viruses in squash (Cucurbita pepo) in Georgia. Plant Dis. 2021, 105, 1862. [Google Scholar] [CrossRef] [PubMed]
- Kavalappara, S.R.; Milner, H.; Konakalla, N.C.; Morgan, K.; Sparks, A.N.; McGregor, C.; Culbreath, A.K.; Wintermantel, W.M.; Bag, S. High-throughput sequencing-aided survey reveals widespread mixed infections of whitefly-transmitted viruses in cucurbits in Georgia, USA. Viruses 2021, 13, 988. [Google Scholar] [CrossRef] [PubMed]
- Adeleke, I.A.; Kavalappara, S.R.; Torrance, T.; Bennett, J.E.; McGregor, C.; Srinivasan, R.; Bag, S. First report of watermelon crinkle leaf-associated virus 1 naturally infecting watermelon (Citrullus lanatus) in Georgia, USA. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Baker, C.; Webb, S.; Adkins, S. Squash Vein Yellowing Virus, Causal Agent of Watermelon Vine Declines in Florida. Florida Department of Agriculture & Consumer Services. Plant Pathology Circular No. 407; June–July 2008. Available online: https://www.fdacs.gov/content/download/11413/file/%20pp407.pdf (accessed on 21 April 2022).
- Hernandez, R.N.; Isakeit, T.; Al Rwahnih, M.; Villegas, C.; Alabi, O.J. First report of watermelon crinkle leaf-associated virus 1 (WCLaV-1) and WCLaV-2 infecting watermelon (Citrullus lanatus) in the United States. Plant Dis. 2021, 105, 2025. [Google Scholar] [CrossRef]
- Hendricks, K.E.M.; Hernandez, R.N.; Roberts, P.D.; Isakeit, T.; Alabi, O.J. First report of watermelon crinkle leaf-associated virus 1 (WCLaV-1) and WCLaV-2 in watermelon (Citrullus lanatus) plants coinfected with cucurbit chlorotic yellows virus in Florida. Plant Dis. 2022, 106, 339. [Google Scholar] [CrossRef]
- Mlotshwa, S.; Pruss, G.J.; Vance, V. Small RNAs in viral infection and host defense. Trends Plant Sci. 2008, 13, 375–382. [Google Scholar] [CrossRef]
- Boonham, N.; Kreuze, J.; Winter, S.; van der Vlugt, R.; Bergervoet, J.; Tomlinson, J.; Mumford, R. Methods in virus diagnostics: From ELISA to next generation sequencing. Virus Res. 2014, 186, 20–31. [Google Scholar] [CrossRef]
- Bag, S.; Al Rwahnih, M.; Li, A.; Gonzalez, A.; Rowhani, A.; Uyemoto, J.K.; Sudarshana, M.R. Detection of a new luteovirus in imported nectarine trees: A case study to propose adoption of metagenomics in post-entry quarantine. Phytopathology 2015, 105, 840–846. [Google Scholar] [CrossRef] [Green Version]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant virus metagenomics: Advances in virus discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pooggin, M.M. Small RNA-omics for plant virus identification, virome reconstruction, and antiviral defense characterization. Front Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef] [PubMed]
- Massart, S.; Chiumenti, M.; De Jonghe, K.; Glover, R.; Haegeman, A.; Koloniuk, I.; Kominek, P.; Kreuze, J.; Kutnjak, D.; Lotos, L.; et al. Virus detection by high-throughput sequencing of small RNAs: Large-scale performance testing of sequence analysis strategies. Phytopathology 2019, 109, 488–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenele, R.S.; Salywon, A.M.; Majure, L.C.; Cobb, I.N.; Bhaskara, A.; Avalos-Calleros, J.A.; Argüello-Astorga, G.R.; Schmidlin, K.; Khalifeh, A.; Smith, K.; et al. A novel divergent geminivirus identified in asymptomatic new world cactaceae plants. Viruses 2020, 12, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowkes, A.R.; McGreig, S.; Pufal, H.; Duffy, S.; Howard, B.; Adams, I.P.; Macarthur, R.; Weekes, R.; Fox, A. Integrating high throughput sequencing into survey design reveals turnip yellows virus and soybean dwarf virus in pea (Pisum sativum) in the United Kingdom. Viruses 2021, 13, 2530. [Google Scholar] [CrossRef]
- Kwibuka, Y.; Bisimwa, E.; Blouin, A.G.; Bragard, C.; Candresse, T.; Faure, C.; Filloux, D.; Lett, J.-M.; Maclot, F.; Marais, A.; et al. Ampeloviruses infecting cassava in central Africa and the south-west Indian Ocean Islands. Viruses 2021, 13, 1030. [Google Scholar] [CrossRef]
- Pecman, A.; Kutnjak, D.; Gutiérrez-Aguirre, I.; Adams, I.; Fox, A.; Boonham, N.; Ravnikar, M. Next generation sequencing for detection and discovery of plant viruses and viroids: Comparison of two approaches. Front Microbiol. 2017, 8, 1998. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Zhan, B.; Cao, M.; Wang, K.; Wang, X.; Zhou, X. Detection and characterization of cucumis melo cryptic virus, Cucumis melo amalgavirus-1, and melon necrotic spot virus in Cucumis melo. Viruses 2019, 11, 81. [Google Scholar] [CrossRef] [Green Version]
- Sabanadzovic, S.; Wintermantel, W.M.; Valverde, R.A.; McCreight, J.D.; Aboughanem-Sabanadzovic, N. Cucumis melo endornavirus: Genome organization, host range and co-divergence with the host. Virus Res. 2016, 214, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Jenkins Hladky, L.L.; Fashing, P.L.; McCreight, J.D.; Turini, T.A.; Wintermantel, W.M. First report of cucurbit yellow stunting disorder virus and cucurbit chlorotic yellows virus in melon in the Central Valley of California. Plant Dis. 2021, 105, 3768. [Google Scholar] [CrossRef]
- Tomasechova, J.; Hancinsky, R.; Predajna, L.; Kraic, J.; Mihalik, D.; Soltys, K.; Vavrova, S.; Bohmer, M.; Sabanadzovic, S.; Glasa, M. High-throughput sequencing reveals bell pepper endornavirus infection in pepper (Capsicum annum) in Slovakia and enables its further molecular characterization. Plants 2019, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Villamor, D.E.V.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E. High throughput sequencing for plant virus detection and discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef]
- Charon, J.; Marcelino, V.R.; Wetherbee, R.; Verbruggen, H.; Holmes, E.C. Metatranscriptomic identification of diverse and divergent RNA viruses in green and chlorarachniophyte algae cultures. Viruses 2020, 12, 1180. [Google Scholar] [CrossRef]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Mumford, R.; Jackeviciene, E.; Navalinskiene, M.; Samuitiene, M.; Boonham, N. Next-generation sequencing and metagenomic analysis: A universal diagnostic tool in plant virology. Mol. Plant Pathol. 2009, 10, 537–545. [Google Scholar] [CrossRef]
- Li, R.; Gao, S.; Hernandez, A.G.; Wechter, W.P.; Fei, Z.; Ling, K.S. Deep sequencing of small RNAs in tomato for virus and viroid identification and strain differentiation. PLoS ONE 2012, 7, e37127. [Google Scholar] [CrossRef]
- Xin, M.; Cao, M.; Liu, W.; Ren, Y.; Lu, C.; Wang, X. The genomic and biological characterization of Citrullus lanatus cryptic virus infecting watermelon in China. Virus Res. 2017, 232, 106–112. [Google Scholar] [CrossRef]
- Zhang, S.; Tian, X.; Navarro, B.; Di Serio, F.; Cao, M. Watermelon crinkle leaf-associated virus 1 and watermelon crinkle leaf-associated virus 2 have a bipartite genome with molecular signatures typical of the members of the genus Coguvirus (family Phenuiviridae). Arch. Virol. 2021, 166, 2829–2834. [Google Scholar] [CrossRef]
- Maeda, M.H.K.; Koyama, L.H.H.; Campos, R.N.S.; Kauffmann, C.M.; Souza, J.O.; Gilbertson, R.; Inoue-Nagata, A.K.; Freitas, D.M.S.; Nogueira, D.R.S.; Melo, F.L.; et al. First report of watermelon crinkle leaf-associated virus 1 and 2 infecting watermelon (Citrullus lanatus) plants in Brazil. Plant Dis. 2022, 106, 773. [Google Scholar] [CrossRef]
- Roossinck, M.J. Lifestyles of plant viruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1899–1905. [Google Scholar] [CrossRef] [Green Version]
- Fauquet, C.; Stanley, J. Revising the way we conceive and name viruses below the species level: A review of geminivirus taxonomy calls for new standardized isolate descriptors. Arch. Virol. 2005, 150, 2151–2179. [Google Scholar] [CrossRef]
- Boccardo, G.; Lisa, V.; Luisoni, E.; Milne, R.G. Cryptic plant viruses. Adv. Virus Res. 1987, 32, 171–214. [Google Scholar] [CrossRef]
- Fukuhara, T. Endornaviruses: Persistent dsRNA viruses with symbiotic properties in diverse eukaryotes. Virus Genes 2019, 55, 165–173. [Google Scholar] [CrossRef]
- Fukuhara, T.; Moriyama, H. Endornaviruses. In Encyclopedia of Virology, 3rd ed.; Mahy, B.W.J., van Regenmortel, M.H.V., Eds.; Academic Press: Cambridge, MA, USA, 2008; pp. 109–116. [Google Scholar] [CrossRef]
- Fukuhara, T.; Koga, R.; Aoki, N.; Yuki, C.; Yamamoto, N.; Oyama, N.; Udagawa, T.; Horiuchi, H.; Miyazaki, S.; Higashi, Y.; et al. The wide distribution of endornaviruses, large double-stranded RNA replicons with plasmid-like properties. Arch. Virol. 2006, 151, 995–1002. [Google Scholar] [CrossRef]
- Valverde, R.A.; Khalifa, M.E.; Okada, R.; Fukuhara, T.; Sabanadzovic, S. ICTV virus taxonomy profile: Endornaviridae. J. Gen. Virol. 2019, 100, 1204–1205. [Google Scholar] [CrossRef]
- Bernardo, P.; Charles-Dominique, T.; Barakat, M.; Ortet, P.; Fernandez, E.; Filloux, D.; Hartnady, P.; Rebelo, T.A.; Cousins, S.R.; Mesleard, F.; et al. Geometagenomics illuminates the impact of agriculture on the distribution and prevalence of plant viruses at the ecosystem scale. ISME J. 2018, 12, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Park, D.; Goh, C.J.; Kim, H.; Hahn, Y. Identification of two novel amalgaviruses in the common eelgrass (Zostera marina) and in-silico analysis of the Amalgavirus +1 programmed ribosomal frameshifting sites. Plant Pathol. J 2018, 34, 150–156. [Google Scholar] [CrossRef]
- Osaki, H.; Sasaki, A. A novel alphapartitivirus detected in Japanese pear. Virus Genes 2018, 54, 149–154. [Google Scholar] [CrossRef]
- Susi, H.; Filloux, D.; Frilander, M.J.; Roumagnac, P.; Laine, A.L. Diverse and variable virus communities in wild plant populations revealed by metagenomic tools. PeerJ 2019, 7, e6140. [Google Scholar] [CrossRef]
- Bejerman, N.; Roumagnac, P.; Nemchinov, L.G. High-throughput sequencing for deciphering the virome of alfalfa (Medicago sativa L.). Front. Microbiol. 2020, 11, 553109. [Google Scholar] [CrossRef]
- Ma, Y.; Marais, A.; Lefebvre, M.; Faure, C.; Candresse, T. Metagenomic analysis of virome cross-talk between cultivated Solanum lycopersicum and wild Solanum nigrum. Virology 2020, 540, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Galipienso, L.; Elvira-González, L.; Velasco, L.; Herrera-Vásquez, J.Á.; Rubio, L. Detection of persistent viruses by high-throughput sequencing in tomato and pepper from panama: Phylogenetic and evolutionary studies. Plants 2021, 10, 2295. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, T.; Abe, J. High-throughput sequencing indicates novel varicosavirus, emaravirus, and deltapartitivirus infections in vitis coignetiae. Viruses 2021, 13, 827. [Google Scholar] [CrossRef]
- Valverde, R.A.; Gutierrez, D.L. Transmission of a dsRNA in bell pepper and evidence that it consists of the genome of an endornavirus. Virus Genes 2007, 35, 399–403. [Google Scholar] [CrossRef]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M.; Ictv Report, C. ICTV Virus Taxonomy Profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Lusk, R.W. Diverse and widespread contamination evident in the unmapped depths of high throughput sequencing data. PLoS ONE 2014, 9, e110808. [Google Scholar] [CrossRef] [Green Version]
- Merchant, S.; Wood, D.E.; Salzberg, S.L. Unexpected cross-species contamination in genome sequencing projects. PeerJ 2014, 2, e675. [Google Scholar] [CrossRef]
- Tosar, J.P.; Rovira, C.; Naya, H.; Cayota, A. Mining of public sequencing databases supports a non-dietary origin for putative foreign miRNAs: Underestimated effects of contamination in NGS. RNA 2014, 20, 754–757. [Google Scholar] [CrossRef] [Green Version]
- Ballenghien, M.; Faivre, N.; Galtier, N. Patterns of cross-contamination in a multispecies population genomic project: Detection, quantification, impact, and solutions. BMC Biol. 2017, 15, 25. [Google Scholar] [CrossRef] [Green Version]
- Kavalappara, S.R.; Milner, H.; Riley, D.G.; Bag, S. First report of turnip yellows virus infecting cabbage (Brassica oleracea var. capitata) in the USA. Plant Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Barman, A.K.; Roberts, P.M.; Prostko, E.P.; Toews, M.D. Seasonal occurrence and reproductive suitability of weed hosts for sweetpotato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae), in South Georgia. J. Entomol. Sci. 2021, 57, 1–11. [Google Scholar] [CrossRef]
- Kavalappara, S.R.; Riley, D.G.; Cremonez, P.S.G.; Perier, J.D.; Bag, S. Wild radish (Raphanus raphanistrum L.) is a potential reservoir host of cucurbit chlorotic yellows virus. Viruses 2022, 14, 593. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences 5′–3′ | Tm (°C) | Amplicon Size | References |
|---|---|---|---|---|
| CmAV1-2459F | AACCTCCCACATTCTGGA | 55 | 740 | [21] |
| CmAV1-3189R | TCCAGTCAGCATAGGTCTCC | |||
| CmEV-1F | ACCCCACTATTAGATATGCTAAGGTC | 55 | 680 | [22] |
| CmEV-1R | CTCCAGGAGTAAGATATAATGTAACCG | |||
| CmCV-109F | ACTGAAGGATGAGTTCGCA | 55 | 640 | [21] |
| CmCV-751R | CCATCGGCATTCAGAACT | |||
| CCYV_RDRP_1515 | CTCCGAGTAGATCATCCCAAATC | 62 | 953 | [4] |
| CCYV_RDRP_1515 | TCACCAGAAACTCCACAATCTC | |||
| CuLCrV CP 259 F | TCAAAGGTTTCCCGCTCTGC | 58 | 588 | [5] |
| CuLCrV CP 846 R | TCCTGCTTCCTGGTGGTTGTAG | |||
| CYSDV_RDRP_1542 | TTTCGGCTCCCAGAGTTAATG | 58 | 492 | [23] |
| CYSDV_RDRP_1542 | CGATCTCCGTGGTGTGATAAG | |||
| WCLaV-1F | GGTGAGTTAGTGTGTCTGAAGG | 55 | 881 | [8] |
| WCLaV-1R | GAGGTTGCCTGAGGTGATAAG |
| Virus Detected | Sample ID & Location | Genome Size of Refseq (nt) | Total sRNA Reads | Reads Matching to Virus | Coverage (%) | Nucleotide Identity (%) |
|---|---|---|---|---|---|---|
| CmAV1 | WM3 | 3424 | 17,110,156 | 54,623 (0.31) | 3395 (99.1) | 99.18 |
| Wilcox | ||||||
| WM4 | 3424 | 19,040,070 | 189,757 (0.99) | 3397 (99.2) | 99.24 | |
| Wilcox | ||||||
| CA | 3424 | 18,645,801 | 27,819 (0.14) | 3419 (99.8) | 99.85 | |
| Turner | ||||||
| WM2 | 3424 | 28,380,401 | 16,477 (0.05) | 3398 (99.2) | 99.24 | |
| Wilcox | ||||||
| WM5 | 3424 | 21,552,412 | 30,070 (0.13) | 3398 (99.2) | 99.27 | |
| Wilcox | ||||||
| CmCV | WM2 | 1592 RNA 1 1715 RNA 2 | 28,380,401 | 144,683 (0.5) | 1587 RNA 1 (99.9) | 99.94 RNA 1 |
| Wilcox | 1714 RNA 2 (99.6) | 99.60 RNA 2 | ||||
| CmEV | WM 1 | 15078 | 18,645,801 | 273,320 (1.46) | 14,577 (96.6) | 96.68 |
| Tift | ||||||
| WCLaV-1 | WM2 | 6645 RNA 1 2682 RNA 2 | 28,380,401 | 557,592 (1.96) | 6599 RNA 1 (99.3) | 99.71(RNA 1) |
| Wilcox | 2678 RNA 2 (99.8) | 99.30 (RNA 2) | ||||
| CA | 6645 RNA 1 2682 RNA 2 | 18,645,801 | 239,859 (1.28) | 6601 RNA 1 (99.3) | 99.73 (RNA 1) | |
| Turner | 2678 RNA 2 (99.8) | 99.30 (RNA 2) |
| Virus | 2021 | 2019 | Virus Detected in Number of Samples | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Watermelon | Cantaloupe | Watermelon | ||||||||
| Colquitt | Crisp | Worth | Wilcox | Turner | Tift | Turner | Tift | Colquitt | ||
| WCLaV-1 | 2(8) | - | 1(5) | 17(28) | 33(83) | 39 (98) | - | - | - | 92 |
| CmAV1 | - | 47 (78) | 1(5) | 37(62) | 27(68) | 20(50) | 1(3) | 6(15) | - | 139 |
| CmEV | - | - | - | 8(20) | - | 40 (100) | 40 (100) | - | 88 | |
| CCYV | - | - | - | - | - | - | - | - | 34 (79) | 34 |
| CYSDV | - | - | - | - | - | - | - | - | 1 (2) | 1 |
| CuLCrV | - | - | - | - | - | - | - | - | 35 (81) | 35 |
| CmCV | - | - | - | - | - | - | - | - | - | - |
| Total sample tested | 25 | 60 | 20 | 60 | 40 | 40 | 40 | 40 | 43 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeleke, I.A.; Kavalappara, S.R.; McGregor, C.; Srinivasan, R.; Bag, S. Persistent, and Asymptomatic Viral Infections and Whitefly-Transmitted Viruses Impacting Cantaloupe and Watermelon in Georgia, USA. Viruses 2022, 14, 1310. https://doi.org/10.3390/v14061310
Adeleke IA, Kavalappara SR, McGregor C, Srinivasan R, Bag S. Persistent, and Asymptomatic Viral Infections and Whitefly-Transmitted Viruses Impacting Cantaloupe and Watermelon in Georgia, USA. Viruses. 2022; 14(6):1310. https://doi.org/10.3390/v14061310
Chicago/Turabian StyleAdeleke, Ismaila Adeyemi, Saritha Raman Kavalappara, Cecilia McGregor, Rajagopalbabu Srinivasan, and Sudeep Bag. 2022. "Persistent, and Asymptomatic Viral Infections and Whitefly-Transmitted Viruses Impacting Cantaloupe and Watermelon in Georgia, USA" Viruses 14, no. 6: 1310. https://doi.org/10.3390/v14061310
APA StyleAdeleke, I. A., Kavalappara, S. R., McGregor, C., Srinivasan, R., & Bag, S. (2022). Persistent, and Asymptomatic Viral Infections and Whitefly-Transmitted Viruses Impacting Cantaloupe and Watermelon in Georgia, USA. Viruses, 14(6), 1310. https://doi.org/10.3390/v14061310