
The Penis, the Vagina and HIV Risk: Key Differences (Aside from the Obvious)

 , ,
, , {kind=link}
{kind=link}
Abstract
1. Introduction
2. Sexual HIV Transmission
- Per-coitus risk of HIV transmission is comparable between sites.
- Both are stratified squamous epithelia.
- Immune activation is associated with increased risk, and quiescence with protection.
- Antimicrobial peptides with anti-HIV activity are associated with increased risk.
- STIs increase inflammation and HIV risk.
- Genital washing increases inflammation and HIV risk.
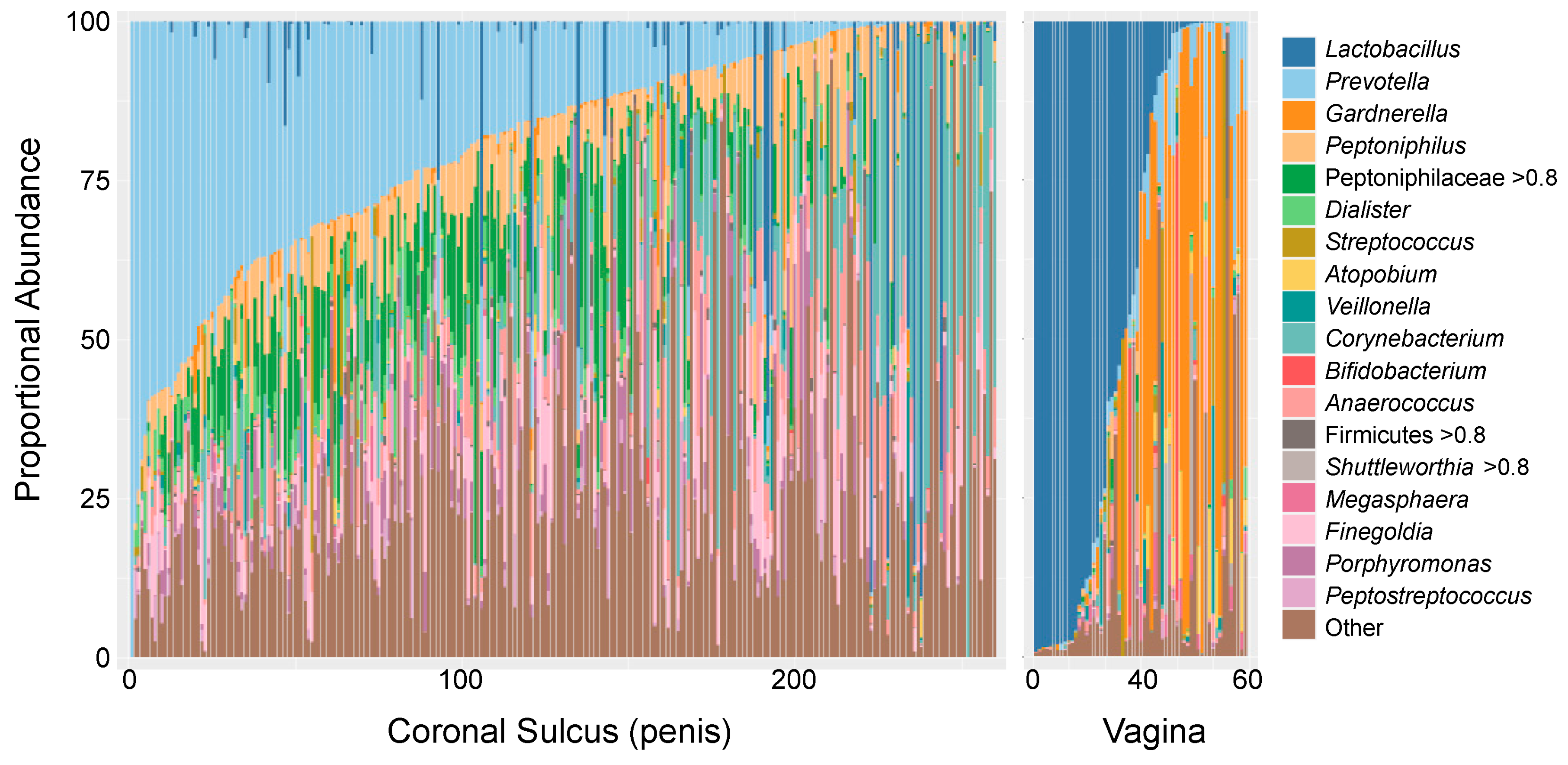
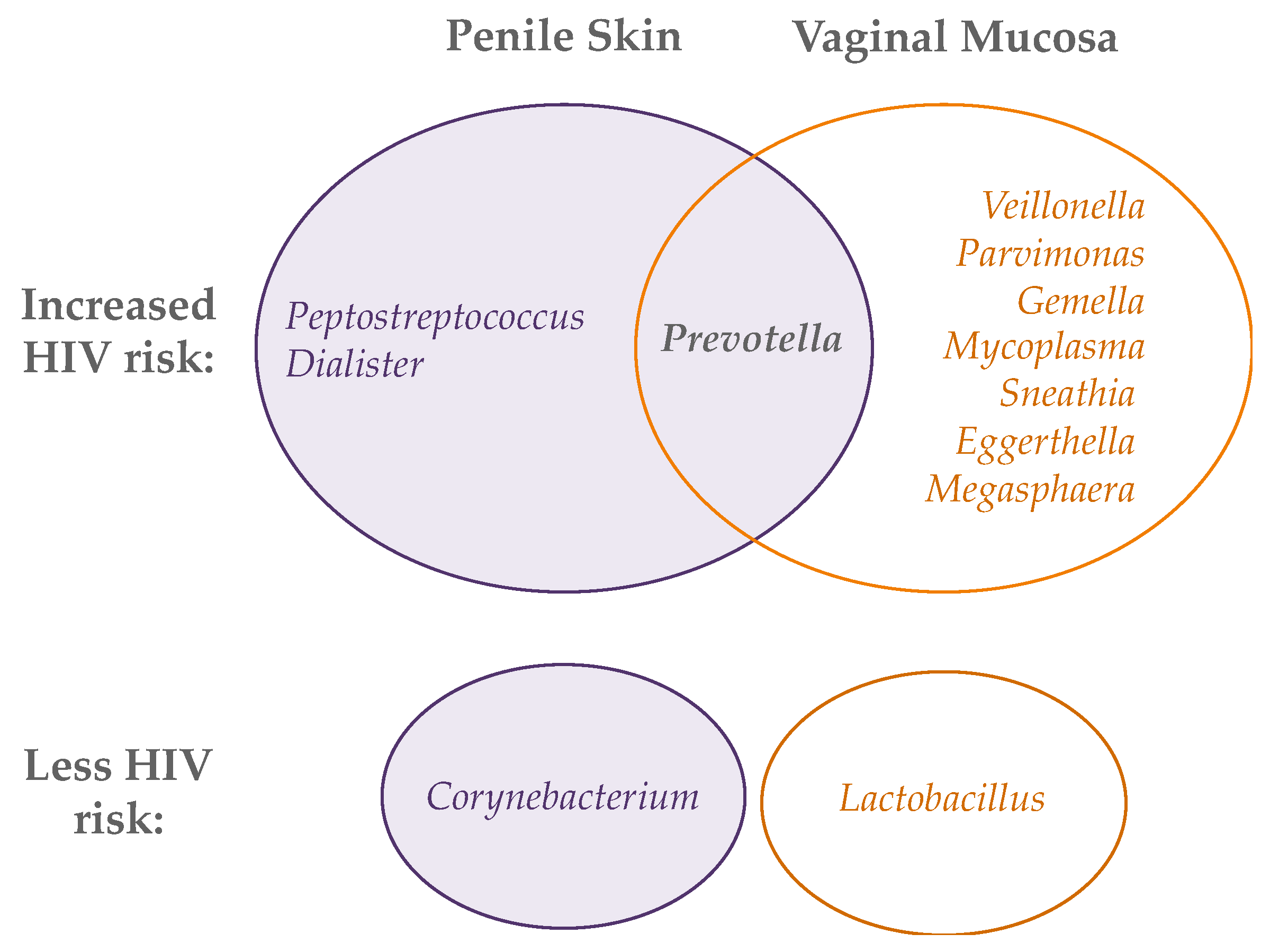
- Abundances of Prevotella spp. correlate with inflammation and HIV risk.
- Epithelial structure: the penis has a cornified outer layer and no goblet cells.
- On the penis, different STIs infect and inflame different sites: HSV-2, HPV, and chancroid affect the skin, while chlamydia, gonorrhea, syphilis and trichomoniasis affect the urethra.
- In the vagina, microbiota dominated by Lactobacillus spp. are associated with lower risk of HIV; on the penis, microbiotas with low anaerobe abundance are associated with lower risk.
3. Genital Immunology and HIV Risk
3.1. Epithelial Structure
3.2. HIV Target Cells
3.3. Genital Cytokines
3.4. Innate Antimicrobial Peptides
3.5. Individuals Who Are HIV-Exposed but Remain SeroNegative (HESN)
4. Clinical Factors That Alter Genital Immunology and Enhance Risk
4.1. Genital Infections
4.2. Genital Hygiene and Products
4.3. Contraception
4.4. Penile Circumcision
5. Penile and Vaginal Microbiome in HIV Risk
6. Implications and Future Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- UNAIDS Global AIDS Update 2021. Confronting Inequalities: Lessons for Pandemic Responses from 40 Years of AIDS. Available online: https://www.unaids.org/sites/default/files/media_asset/2021-global-aids-update_en.pdf (accessed on 26 April 2022).
- Brown, L.B.; Spinelli, M.A.; Gandhi, M. The interplay between HIV and COVID-19: Summary of the data and responses to date. Curr. Opin. HIV AIDS 2021, 16, 63–73. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. UNAIDS Data 2021; Joint United Nations Programme on HIV/AIDS: Geneva, Switzerland, 2021; Volume 2021, p. 468. [Google Scholar]
- Kaul, R.; Prodger, J.; Joag, V.; Shannon, B.; Yegorov, S.; Galiwango, R.; McKinnon, L. Inflammation and HIV Transmission in Sub-Saharan Africa. Curr. HIV/AIDS Rep. 2015, 12, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Kaul, R.; Cohen, C.R.; Chege, D.; Yi, T.J.; Tharao, W.; McKinnon, L.R.; Remis, R.; Anzala, O.; Kimani, J. Biological factors that may contribute to regional and racial disparities in HIV prevalence. Am. J. Reprod. Immunol. 2011, 65, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Cavarelli, M.; Le Grand, R. The importance of semen leukocytes in HIV-1 transmission and the development of prevention strategies. Hum. Vaccin Immunother. 2020, 16, 2018–2032. [Google Scholar] [CrossRef]
- Rodger, A.J.; Cambiano, V.; Bruun, T.; Vernazza, P.; Collins, S.; van Lunzen, J.; Corbelli, G.M.; Estrada, V.; Geretti, A.M.; Beloukas, A.; et al. Sexual Activity Without Condoms and Risk of HIV Transmission in Serodifferent Couples When the HIV-Positive Partner Is Using Suppressive Antiretroviral Therapy. JAMA 2016, 316, 171–181. [Google Scholar] [CrossRef]
- Rodger, A.J.; Cambiano, V.; Bruun, T.; Vernazza, P.; Collins, S.; Degen, O.; Corbelli, G.M.; Estrada, V.; Geretti, A.M.; Beloukas, A.; et al. Risk of HIV transmission through condomless sex in serodifferent gay couples with the HIV-positive partner taking suppressive antiretroviral therapy (PARTNER): Final results of a multicentre, prospective, observational study. Lancet 2019, 393, 2428–2438. [Google Scholar] [CrossRef]
- Bavinton, B.R.; Pinto, A.N.; Phanuphak, N.; Grinsztejn, B.; Prestage, G.P.; Zablotska-Manos, I.B.; Jin, F.; Fairley, C.K.; Moore, R.; Roth, N.; et al. Viral suppression and HIV transmission in serodiscordant male couples: An international, prospective, observational, cohort study. Lancet HIV 2018, 5, e438–e447. [Google Scholar] [CrossRef]
- Patel, P.; Borkowf, C.B.; Brooks, J.T.; Lasry, A.; Lansky, A.; Mermin, J. Estimating per-act HIV transmission risk: A systematic review. AIDS 2014, 28, 1509–1519. [Google Scholar] [CrossRef]
- Stewart, J.; Douglas, G.; O’Rourke, T.; Gammel, C. Promoting safer sex in the context of heterosexual anal intercourse: A scoping review. J. Clin. Nurs. 2021, 30, 2111–2130. [Google Scholar] [CrossRef]
- McKinnon, L.R.; Kaul, R. Quality and quantity: Mucosal CD4+ T cells and HIV susceptibility. Curr. Opin. HIV AIDS 2012, 7, 195–202. [Google Scholar] [CrossRef]
- Stoner, M.C.D.; Nguyen, N.; Kilburn, K.; Gomez-Olive, F.X.; Edwards, J.K.; Selin, A.; Hughes, J.P.; Agyei, Y.; Macphail, C.; Kahn, K.; et al. Age-disparate partnerships and incident HIV infection in adolescent girls and young women in rural South Africa. AIDS 2019, 33, 83–91. [Google Scholar] [CrossRef]
- Yi, T.J.; Shannon, B.; Prodger, J.; McKinnon, L.; Kaul, R. Genital immunology and HIV susceptibility in young women. Am. J. Reprod. Immunol. 2013, 69 (Suppl. 1), 74–79. [Google Scholar] [CrossRef]
- Klot, J.F.; Auerbach, J.D.; Veronese, F.; Brown, G.; Pei, A.; Wira, C.R.; Hope, T.J.; M’Boup, S. Greentree Meeting on Sexual Violence and HIV (Kaul, Greentree white paper: Sexual violence, genitoanal injury, and HIV: Priorities for research, policy, and practice. AIDS Res. Hum. Retrovir. 2012, 28, 1379–1388. [Google Scholar] [CrossRef]
- Weiss, H.A.; Buve, A.; Robinson, N.J.; Van Dyck, E.; Kahindo, M.; Anagonou, S.; Musonda, R.; Zekeng, L.; Morison, L.; Carael, M.; et al. The epidemiology of HSV-2 infection and its association with HIV infection in four urban African populations. Aids 2001, 15 (Suppl. 4), S97–S108. [Google Scholar] [CrossRef]
- Prodger, J.L.; Kaul, R. The biology of how circumcision reduces HIV susceptibility: Broader implications for the prevention field. AIDS Res. Ther. 2017, 14, 49. [Google Scholar] [CrossRef]
- Gray, R.H.; Wawer, M.J.; Brookmeyer, R.; Sewankambo, N.K.; Serwadda, D.; Wabwire-Mangen, F.; Lutalo, T.; Li, X.; vanCott, T.; Quinn, T.C. Probability of HIV-1 transmission per coital act in monogamous, heterosexual, HIV-1-discordant couples in Rakai, Uganda. Lancet 2001, 357, 1149–1153. [Google Scholar] [CrossRef]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2009, 214, 516–559. [Google Scholar] [CrossRef]
- Menon, G.K.; Cleary, G.W.; Lane, M.E. The structure and function of the stratum corneum. Int. J. Pharm. 2012, 435, 3–9. [Google Scholar] [CrossRef]
- Anderson, D.J.; Marathe, J.; Pudney, J. The structure of the human vaginal stratum corneum and its role in immune defense. Am. J. Reprod. Immunol. 2014, 71, 618–623. [Google Scholar] [CrossRef]
- Houben, E.; Hachem, J.P.; De Paepe, K.; Rogiers, V. Epidermal ceramidase activity regulates epidermal desquamation via stratum corneum acidification. Skin Pharmacol. Physiol. 2008, 21, 111–118. [Google Scholar] [CrossRef]
- Gupta, R.; Ramnani, P. Microbial keratinases and their prospective applications: An overview. Appl. Microbiol. Biotechnol. 2006, 70, 21–33. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Dinh, M.H.; Hirbod, T.; Kigozi, G.; Okocha, E.A.; Cianci, G.C.; Kong, X.; Prodger, J.L.; Broliden, K.; Kaul, R.; Serwadda, D.; et al. No difference in keratin thickness between inner and outer foreskins from elective male circumcisions in Rakai, Uganda. PLoS ONE 2012, 7, e41271. [Google Scholar] [CrossRef]
- Carias, A.M.; Hope, T.J. Barriers of Mucosal Entry of HIV/SIV. Curr. Immunol. Rev. 2019, 15, 4–13. [Google Scholar] [CrossRef]
- Masson, L.; Passmore, J.A.; Liebenberg, L.J.; Werner, L.; Baxter, C.; Arnold, K.B.; Williamson, C.; Little, F.; Mansoor, L.E.; Naranbhai, V.; et al. Genital inflammation and the risk of HIV acquisition in women. Clin. Infect. Dis. 2015, 61, 260–269. [Google Scholar] [CrossRef]
- McKinnon, L.R.; Liebenberg, L.J.; Yende-Zuma, N.; Archary, D.; Ngcapu, S.; Sivro, A.; Nagelkerke, N.; Garcia Lerma, J.G.; Kashuba, A.D.; Masson, L.; et al. Genital inflammation undermines the effectiveness of tenofovir gel in preventing HIV acquisition in women. Nat. Med. 2018, 24, 491–496. [Google Scholar] [CrossRef]
- Prodger, J.L.; Gray, R.H.; Shannon, B.; Shahabi, K.; Kong, X.; Grabowski, K.; Kigozi, G.; Nalugoda, F.; Serwadda, D.; Wawer, M.J.; et al. Chemokine Levels in the Penile Coronal Sulcus Correlate with HIV-1 Acquisition and Are Reduced by Male Circumcision in Rakai, Uganda. PLoS Pathog. 2016, 12, e1006025. [Google Scholar] [CrossRef]
- Carnathan, D.G.; Wetzel, K.S.; Yu, J.; Lee, S.T.; Johnson, B.A.; Paiardini, M.; Yan, J.; Morrow, M.P.; Sardesai, N.Y.; Weiner, D.B.; et al. Activated CD4+CCR5+ T cells in the rectum predict increased SIV acquisition in SIVGag/Tat-vaccinated rhesus macaques. Proc. Natl. Acad. Sci. USA 2015, 112, 518–523. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Wietgrefe, S.W.; Li, Q.; Shore, M.D.; Duan, L.; Reilly, C.; Lifson, J.D.; Haase, A.T. Roles of substrate availability and infection of resting and activated CD4+ T cells in transmission and acute simian immunodeficiency virus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 5640–5645. [Google Scholar] [CrossRef]
- Saba, E.; Grivel, J.C.; Vanpouille, C.; Brichacek, B.; Fitzgerald, W.; Margolis, L.; Lisco, A. HIV-1 sexual transmission: Early events of HIV-1 infection of human cervico-vaginal tissue in an optimized ex vivo model. Mucosal Immunol. 2010, 3, 280–290. [Google Scholar] [CrossRef]
- Meditz, A.L.; Haas, M.K.; Folkvord, J.M.; Melander, K.; Young, R.; McCarter, M.; Mawhinney, S.; Campbell, T.B.; Lie, Y.; Coakley, E.; et al. HLA-DR+ CD38+ CD4+ T lymphocytes have elevated CCR5 expression and produce the majority of R5-tropic HIV-1 RNA in vivo. J. Virol. 2011, 85, 10189–10200. [Google Scholar] [CrossRef] [PubMed]
- Joag, V.R.; McKinnon, L.R.; Liu, J.; Kidane, S.T.; Yudin, M.H.; Nyanga, B.; Kimwaki, S.; Besel, K.E.; Obila, J.O.; Huibner, S.; et al. Identification of preferential CD4+ T-cell targets for HIV infection in the cervix. Mucosal Immunol. 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Zhang, Y.; Monteiro, P.; Goulet, J.P.; Gosselin, A.; Grandvaux, N.; Hope, T.J.; Fassati, A.; Routy, J.P.; Ancuta, P. HIV-1 selectively targets gut-homing CCR6+CD4+ T cells via mTOR-dependent mechanisms. JCI Insight 2017, 2, e93230. [Google Scholar] [CrossRef] [PubMed]
- Prodger, J.L.; Ssemaganda, A.; Ssetaala, A.; Kitandwe, P.K.; Muyanja, E.; Mpendo, J.; Nanvubya, A.; Wambuzi, M.; Nielsen, L.; Kiwanuka, N.; et al. Schistosoma mansoni Infection in Ugandan Men Is Associated with Increased Abundance and Function of HIV Target Cells in Blood, but Not the Foreskin: A Cross-sectional Study. PLoS Negl. Trop. Dis. 2015, 9, e0004067. [Google Scholar] [CrossRef]
- Stieh, D.J.; Matias, E.; Xu, H.; Fought, A.J.; Blanchard, J.L.; Marx, P.A.; Veazey, R.S.; Hope, T.J. Th17 Cells Are Preferentially Infected Very Early after Vaginal Transmission of SIV in Macaques. Cell Host Microbe 2016, 19, 529–540. [Google Scholar] [CrossRef]
- Ryan, E.S.; Micci, L.; Fromentin, R.; Paganini, S.; McGary, C.S.; Easley, K.; Chomont, N.; Paiardini, M. Loss of Function of Intestinal IL-17 and IL-22 Producing Cells Contributes to Inflammation and Viral Persistence in SIV-Infected Rhesus Macaques. PLoS Pathog. 2016, 12, e1005412. [Google Scholar] [CrossRef]
- Kim, C.J.; Nazli, A.; Rojas, O.L.; Chege, D.; Alidina, Z.; Huibner, S.; Mujib, S.; Benko, E.; Kovacs, C.; Shin, L.Y.; et al. A role for mucosal IL-22 production and Th22 cells in HIV-associated mucosal immunopathogenesis. Mucosal Immunol. 2012, 5, 670–680. [Google Scholar] [CrossRef]
- Duhen, T.; Geiger, R.; Jarrossay, D.; Lanzavecchia, A.; Sallusto, F. Production of interleukin 22 but not interleukin 17 by a subset of human skin-homing memory T cells. Nat. Immunol. 2009, 10, 857–863. [Google Scholar] [CrossRef]
- McKinnon, L.R.; Nyanga, B.; Kim, C.J.; Izulla, P.; Kwatampora, J.; Kimani, M.; Shahabi, K.; Mugo, N.; Smith, J.S.; Anzala, A.O.; et al. Early HIV-1 infection is associated with reduced frequencies of cervical Th17 cells. J. Acquir. Immune Defic. Syndr. 2015, 68, 6–12. [Google Scholar] [CrossRef]
- Martinelli, E.; Veglia, F.; Goode, D.; Guerra-Perez, N.; Aravantinou, M.; Arthos, J.; Piatak, M., Jr.; Lifson, J.D.; Blanchard, J.; Gettie, A.; et al. The frequency of alpha(4)beta(7)(high) memory CD4(+) T cells correlates with susceptibility to rectal simian immunodeficiency virus infection. J. Acquir. Immune Defic. Syndr. 2013, 64, 325–331. [Google Scholar] [CrossRef]
- Sivro, A.; Schuetz, A.; Sheward, D.; Joag, V.; Yegorov, S.; Liebenberg, L.J.; Yende-Zuma, N.; Stalker, A.; Mwatelah, R.S.; Selhorst, P.; et al. Integrin alpha4beta7 expression on peripheral blood CD4+ T cells predicts HIV acquisition and disease progression outcomes. Sci. Transl. Med. 2018, 10, eaam6354. [Google Scholar] [CrossRef]
- de Jong, M.A.; de Witte, L.; Oudhoff, M.J.; Gringhuis, S.I.; Gallay, P.; Geijtenbeek, T.B. TNF-alpha and TLR agonists increase susceptibility to HIV-1 transmission by human Langerhans cells ex vivo. J. Clin. Investig. 2008, 118, 3440–3452. [Google Scholar] [CrossRef]
- Fahrbach, K.M.; Barry, S.M.; Ayehunie, S.; Lamore, S.; Klausner, M.; Hope, T.J. Activated CD34-derived Langerhans cells mediate transinfection with human immunodeficiency virus. J. Virol. 2007, 81, 6858–6868. [Google Scholar] [CrossRef]
- Shannon, B.; Gajer, P.; Yi, T.J.; Ma, B.; Humphrys, M.S.; Thomas-Pavanel, J.; Chieza, L.; Janakiram, P.; Saunders, M.; Tharao, W.; et al. Distinct Effects of the Cervicovaginal Microbiota and Herpes Simplex Type 2 Infection on Female Genital Tract Immunology. J. Infect. Dis. 2017, 215, 1366–1375. [Google Scholar] [CrossRef]
- Arnold, K.B.; Burgener, A.; Birse, K.; Romas, L.; Dunphy, L.J.; Shahabi, K.; Abou, M.; Westmacott, G.R.; McCorrister, S.; Kwatampora, J.; et al. Increased levels of inflammatory cytokines in the female reproductive tract are associated with altered expression of proteases, mucosal barrier proteins, and an influx of HIV-susceptible target cells. Mucosal Immunol. 2016, 9, 194–205. [Google Scholar] [CrossRef]
- Kaul, R.; Rebbapragada, A.; Hirbod, T.; Wachihi, C.; Ball, T.B.; Plummer, F.A.; Kimani, J.; Jaoko, W. Genital levels of soluble immune factors with anti-HIV activity may correlate with increased HIV susceptibility. Aids 2008, 22, 2049–2051. [Google Scholar] [CrossRef]
- Gray, R.H.; Kigozi, G.; Serwadda, D.; Makumbi, F.; Watya, S.; Nalugoda, F.; Kiwanuka, N.; Moulton, L.H.; Chaudhary, M.A.; Chen, M.Z.; et al. Male circumcision for HIV prevention in men in Rakai, Uganda: A randomised trial. Lancet 2007, 369, 657–666. [Google Scholar] [CrossRef]
- Abdool Karim, Q.; Abdool Karim, S.S.; Frohlich, J.A.; Grobler, A.C.; Baxter, C.; Mansoor, L.E.; Kharsany, A.B.; Sibeko, S.; Mlisana, K.P.; Omar, Z.; et al. Effectiveness and safety of tenofovir gel, an antiretroviral microbicide, for the prevention of HIV infection in women. Science 2010, 329, 1168–1174. [Google Scholar] [CrossRef]
- Brogden, N.K.; Mehalick, L.; Fischer, C.L.; Wertz, P.W.; Brogden, K.A. The emerging role of peptides and lipids as antimicrobial epidermal barriers and modulators of local inflammation. Skin Pharmacol. Physiol. 2012, 25, 167–181. [Google Scholar] [CrossRef]
- Nakashima, H.; Yamamoto, N.; Masuda, M.; Fujii, N. Defensins inhibit HIV replication in vitro. Aids 1993, 7, 1129. [Google Scholar] [CrossRef]
- Wu, Z.; Cocchi, F.; Gentles, D.; Ericksen, B.; Lubkowski, J.; Devico, A.; Lehrer, R.I.; Lu, W. Human neutrophil alpha-defensin 4 inhibits HIV-1 infection in vitro. FEBS Lett. 2005, 579, 162–166. [Google Scholar] [CrossRef]
- Quinones-Mateu, M.E.; Lederman, M.M.; Feng, Z.; Chakraborty, B.; Weber, J.; Rangel, H.R.; Marotta, M.L.; Mirza, M.; Jiang, B.; Kiser, P.; et al. Human epithelial beta-defensins 2 and 3 inhibit HIV-1 replication. Aids 2003, 17, F39–F48. [Google Scholar] [CrossRef]
- Sun, L.; Finnegan, C.M.; Kish-Catalone, T.; Blumenthal, R.; Garzino-Demo, P.; La Terra Maggiore, G.M.; Berrone, S.; Kleinman, C.; Wu, Z.; Abdelwahab, S.; et al. Human beta-defensins suppress human immunodeficiency virus infection: Potential role in mucosal protection. J. Virol. 2005, 79, 14318–14329. [Google Scholar] [CrossRef]
- Tugizov, S.M.; Herrera, R.; Veluppillai, P.; Greenspan, D.; Soros, V.; Greene, W.C.; Levy, J.A.; Palefsky, J.M. HIV is inactivated after transepithelial migration via adult oral epithelial cells but not fetal epithelial cells. Virology 2011, 409, 211–222. [Google Scholar] [CrossRef]
- Bergman, P.; Walter-Jallow, L.; Broliden, K.; Agerberth, B.; Soderlund, J. The antimicrobial peptide LL-37 inhibits HIV-1 replication. Curr. HIV Res. 2007, 5, 410–415. [Google Scholar] [CrossRef]
- Wong, J.H.; Legowska, A.; Rolka, K.; Ng, T.B.; Hui, M.; Cho, C.H.; Lam, W.W.; Au, S.W.; Gu, O.W.; Wan, D.C. Effects of cathelicidin and its fragments on three key enzymes of HIV-1. Peptides 2011, 32, 1117–1122. [Google Scholar] [CrossRef]
- Steinstraesser, L.; Tippler, B.; Mertens, J.; Lamme, E.; Homann, H.H.; Lehnhardt, M.; Wildner, O.; Steinau, H.U.; Uberla, K. Inhibition of early steps in the lentiviral replication cycle by cathelicidin host defense peptides. Retrovirology 2005, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Demirkhanyan, L.H.; Marin, M.; Padilla-Parra, S.; Zhan, C.; Miyauchi, K.; Jean-Baptiste, M.; Novitskiy, G.; Lu, W.; Melikyan, G.B. Multifaceted mechanisms of HIV-1 entry inhibition by human alpha-defensin. J. Biol. Chem. 2012, 287, 28821–28838. [Google Scholar] [CrossRef] [PubMed]
- Levinson, P.; Choi, R.Y.; Cole, A.L.; Hirbod, T.; Rhedin, S.; Payne, B.; Guthrie, B.L.; Bosire, R.; Cole, A.M.; Farquhar, C.; et al. HIV-neutralizing activity of cationic polypeptides in cervicovaginal secretions of women in HIV-serodiscordant relationships. PLoS ONE 2012, 7, e31996. [Google Scholar] [CrossRef]
- Ghosh, M.; Fahey, J.V.; Shen, Z.; Lahey, T.; Cu-Uvin, S.; Wu, Z.; Mayer, K.; Wright, P.F.; Kappes, J.C.; Ochsenbauer, C.; et al. Anti-HIV activity in cervical-vaginal secretions from HIV-positive and -negative women correlate with innate antimicrobial levels and IgG antibodies. PLoS ONE 2010, 5, e11366. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, N.; Cole, A.L.; Svoboda, P.; Pohl, J.; Cole, A.M. Cationic polypeptides are required for anti-HIV-1 activity of human vaginal fluid. J. Immunol. 2005, 175, 7560–7567. [Google Scholar] [CrossRef]
- Drannik, A.G.; Nag, K.; Yao, X.D.; Henrick, B.M.; Ball, T.B.; Plummer, F.A.; Wachihi, C.; Kimani, J.; Rosenthal, K.L. Anti-HIV-1 activity of elafin depends on its nuclear localization and altered innate immune activation in female genital epithelial cells. PLoS ONE 2012, 7, e52738. [Google Scholar] [CrossRef]
- Drannik, A.G.; Nag, K.; Yao, X.D.; Henrick, B.M.; Jain, S.; Ball, T.B.; Plummer, F.A.; Wachihi, C.; Kimani, J.; Rosenthal, K.L. Anti-HIV-1 activity of elafin is more potent than its precursor’s, trappin-2, in genital epithelial cells. J. Virol. 2012, 86, 4599–4610. [Google Scholar] [CrossRef]
- Ghosh, M.; Shen, Z.; Fahey, J.V.; Cu-Uvin, S.; Mayer, K.; Wira, C.R. Trappin-2/Elafin: A novel innate anti-human immunodeficiency virus-1 molecule of the human female reproductive tract. Immunology 2010, 129, 207–219. [Google Scholar] [CrossRef]
- McNeely, T.B.; Dealy, M.; Dripps, D.J.; Orenstein, J.M.; Eisenberg, S.P.; Wahl, S.M. Secretory leukocyte protease inhibitor: A human saliva protein exhibiting anti-human immunodeficiency virus 1 activity in vitro. J. Clin. Investig. 1995, 96, 456–464. [Google Scholar] [CrossRef]
- McNeely, T.B.; Shugars, D.C.; Rosendahl, M.; Tucker, C.; Eisenberg, S.P.; Wahl, S.M. Inhibition of human immunodeficiency virus type 1 infectivity by secretory leukocyte protease inhibitor occurs prior to viral reverse transcription. Blood 1997, 90, 1141–1149. [Google Scholar] [CrossRef]
- Wahl, S.M.; McNeely, T.B.; Janoff, E.N.; Shugars, D.; Worley, P.; Tucker, C.; Orenstein, J.M. Secretory leukocyte protease inhibitor (SLPI) in mucosal fluids inhibits HIV-I. Oral Dis. 1997, 3 (Suppl. 1), S64–S69. [Google Scholar] [CrossRef]
- Levinson, P.; Kaul, R.; Kimani, J.; Ngugi, E.; Moses, S.; MacDonald, K.S.; Broliden, K.; Hirbod, T. Levels of innate immune factors in genital fluids: Association of alpha defensins and LL-37 with genital infections and increased HIV acquisition. Aids 2009, 23, 309–317. [Google Scholar] [CrossRef]
- Hirbod, T.; Kong, X.; Kigozi, G.; Ndyanabo, A.; Serwadda, D.; Prodger, J.L.; Tobian, A.A.; Nalugoda, F.; Wawer, M.J.; Shahabi, K.; et al. HIV acquisition is associated with increased antimicrobial peptides and reduced HIV neutralizing IgA in the foreskin prepuce of uncircumcised men. PLoS Pathog. 2014, 10, e1004416. [Google Scholar] [CrossRef]
- Wilkinson, T.S.; Roghanian, A.; Simpson, A.J.; Sallenave, J.M. WAP domain proteins as modulators of mucosal immunity. Biochem. Soc. Trans. 2011, 39, 1409–1415. [Google Scholar] [CrossRef]
- Yao, X.D.; Omange, R.W.; Henrick, B.M.; Lester, R.T.; Kimani, J.; Ball, T.B.; Plummer, F.A.; Rosenthal, K.L. Acting locally: Innate mucosal immunity in resistance to HIV-1 infection in Kenyan commercial sex workers. Mucosal Immunol. 2013, 7, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Chege, D.; Chai, Y.; Huibner, S.; Kain, T.; Wachihi, C.; Kimani, M.; Barasa, S.; McKinnon, L.R.; Muriuki, F.K.; Kariri, A.; et al. Blunted IL17/IL22 and Pro-Inflammatory Cytokine Responses in the Genital Tract and Blood of HIV-Exposed, Seronegative Female Sex Workers in Kenya. PLoS ONE 2012, 7, e43670. [Google Scholar] [CrossRef] [PubMed]
- Omollo, K.; Lajoie, J.; Oyugi, J.; Wessels, J.M.; Mwaengo, D.; Kimani, J.; Kaushic, C.; Fowke, K.R. Differential Elevation of Inflammation and CD4(+) T Cell Activation in Kenyan Female Sex Workers and Non-Sex Workers Using Depot-Medroxyprogesterone Acetate. Front. Immunol. 2020, 11, 598307. [Google Scholar] [CrossRef] [PubMed]
- Prodger, J.L.; Hirbod, T.; Kigozi, G.; Nalugoda, F.; Reynolds, S.J.; Galiwango, R.; Shahabi, K.; Serwadda, D.; Wawer, M.J.; Gray, R.H.; et al. Immune correlates of HIV exposure without infection in foreskins of men from Rakai, Uganda. Mucosal Immunol. 2014, 7, 634–644. [Google Scholar] [CrossRef]
- Levine, W.C.; Pope, V.; Bhoomkar, A.; Tambe, P.; Lewis, J.S.; Zaidi, A.A.; Farshy, C.E.; Mitchell, S.; Talkington, D.F. Increase in endocervical CD4 lymphocytes among women with nonulcerative sexually transmitted diseases. J. Infect. Dis. 1998, 177, 167–174. [Google Scholar] [CrossRef]
- Masson, L.; Salkinder, A.L.; Olivier, A.J.; McKinnon, L.R.; Gamieldien, H.; Mlisana, K.; Scriba, T.J.; Lewis, D.A.; Little, F.; Jaspan, H.B.; et al. Relationship between female genital tract infections, mucosal interleukin-17 production and local T helper type 17 cells. Immunology 2015, 146, 557–567. [Google Scholar] [CrossRef]
- Dabee, S.; Barnabas, S.L.; Lennard, K.S.; Jaumdally, S.Z.; Gamieldien, H.; Balle, C.; Happel, A.U.; Murugan, B.D.; Williamson, A.L.; Mkhize, N.; et al. Defining characteristics of genital health in South African adolescent girls and young women at high risk for HIV infection. PLoS ONE 2019, 14, e0213975. [Google Scholar] [CrossRef]
- Zhu, J.; Hladik, F.; Woodward, A.; Klock, A.; Peng, T.; Johnston, C.; Remington, M.; Magaret, A.; Koelle, D.M.; Wald, A.; et al. Persistence of HIV-1 receptor-positive cells after HSV-2 reactivation is a potential mechanism for increased HIV-1 acquisition. Nat. Med. 2009, 15, 886–892. [Google Scholar] [CrossRef]
- Shannon, B.; Yi, T.J.; Thomas-Pavanel, J.; Chieza, L.; Janakiram, P.; Saunders, M.; Tharao, W.; Huibner, S.; Remis, R.; Rebbapragada, A.; et al. Impact of asymptomatic herpes simplex virus type 2 infection on mucosal homing and immune cell subsets in the blood and female genital tract. J. Immunol. 2014, 192, 5074–5082. [Google Scholar] [CrossRef]
- Posavad, C.M.; Zhao, L.; Dong, L.; Jin, L.; Stevens, C.E.; Magaret, A.S.; Johnston, C.; Wald, A.; Zhu, J.; Corey, L.; et al. Enrichment of herpes simplex virus type 2 (HSV-2) reactive mucosal T cells in the human female genital tract. Mucosal Immunol. 2017, 10, 1259–1269. [Google Scholar] [CrossRef]
- Cohen, M.S.; Council, O.D.; Chen, J.S. Sexually transmitted infections and HIV in the era of antiretroviral treatment and prevention: The biologic basis for epidemiologic synergy. J. Int. AIDS Soc. 2019, 22 (Suppl. 6), e25355. [Google Scholar] [CrossRef]
- Forman, M.S.; Vaidya, D.; Bolorunduro, O.; Diener-West, M.; Pass, R.F.; Arav-Boger, R. Cytomegalovirus Kinetics Following Primary Infection in Healthy Women. J. Infect. Dis. 2017, 215, 1523–1526. [Google Scholar] [CrossRef]
- Begnel, E.R.; Drake, A.L.; Kinuthia, J.; Matemo, D.; Huang, M.L.; Asbjornsdottir, K.H.; Chohan, V.; Beima-Sofie, K.; John-Stewart, G.; Lehman, D.; et al. Cervical cytomegalovirus reactivation, cytokines and spontaneous preterm birth in Kenyan women. Clin. Exp. Immunol. 2021, 203, 472–479. [Google Scholar] [CrossRef]
- Shannon, B.; Yi, T.J.; Perusini, S.; Gajer, P.; Ma, B.; Humphrys, M.S.; Thomas-Pavanel, J.; Chieza, L.; Janakiram, P.; Saunders, M.; et al. Association of HPV infection and clearance with cervicovaginal immunology and the vaginal microbiota. Mucosal Immunol. 2017, 10, 1310–1319. [Google Scholar] [CrossRef]
- Liebenberg, L.J.P.; McKinnon, L.R.; Yende-Zuma, N.; Garrett, N.; Baxter, C.; Kharsany, A.B.M.; Archary, D.; Rositch, A.; Samsunder, N.; Mansoor, L.E.; et al. HPV infection and the genital cytokine milieu in women at high risk of HIV acquisition. Nat. Commun. 2019, 10, 5227. [Google Scholar] [CrossRef]
- van de Wijgert, J.H.; Morrison, C.S.; Cornelisse, P.G.; Munjoma, M.; Moncada, J.; Awio, P.; Wang, J.; Van der Pol, B.; Chipato, T.; Salata, R.A.; et al. Bacterial vaginosis and vaginal yeast, but not vaginal cleansing, increase HIV-1 acquisition in African women. J. Acquir. Immune Defic. Syndr. 2008, 48, 203–210. [Google Scholar] [CrossRef]
- Hester, R.A.; Kennedy, S.B. Candida infection as a risk factor for HIV transmission. J. Womens Health 2003, 12, 487–494. [Google Scholar] [CrossRef]
- Perez-Soto, E.; Fernandez-Martinez, E.; Oros-Pantoja, R.; Medel-Flores, O.; Miranda-Covarrubias, J.C.; Sanchez-Monroy, V. Proinflammatory and Oxidative Stress States Induced by Human Papillomavirus and Chlamydia trachomatis Coinfection Affect Sperm Quality in Asymptomatic Infertile Men. Medicina 2021, 57, 862. [Google Scholar] [CrossRef]
- Gray, C.M.; O’Hagan, K.L.; Lorenzo-Redondo, R.; Olivier, A.J.; Amu, S.; Chigorimbo-Murefu, N.; Harryparsad, R.; Sebaa, S.; Maziya, L.; Dietrich, J.; et al. Impact of chemokine C-C ligand 27, foreskin anatomy and sexually transmitted infections on HIV-1 target cell availability in adolescent South African males. Mucosal Immunol. 2020, 13, 118–127. [Google Scholar] [CrossRef]
- Gianella, S.; Strain, M.C.; Rought, S.E.; Vargas, M.V.; Little, S.J.; Richman, D.D.; Spina, C.A.; Smith, D.M. Associations between virologic and immunologic dynamics in blood and in the male genital tract. J. Virol. 2012, 86, 1307–1315. [Google Scholar] [CrossRef]
- Sheth, P.M.; Yi, T.J.; Kovacs, C.; Kemal, K.S.; Jones, R.B.; Osborne, B.; Pilon, R.; la Porte, C.; Ostrowski, M.; Mazzulli, T.; et al. Mucosal correlates of isolated HIV semen shedding during effective antiretroviral therapy. Mucosal Immunol. 2012, 5, 248–257. [Google Scholar] [CrossRef]
- Johnson, K.E.; Redd, A.D.; Quinn, T.C.; Collinson-Streng, A.N.; Cornish, T.; Kong, X.; Sharma, R.; Tobian, A.A.; Tsai, B.; Sherman, M.E.; et al. Effects of HIV-1 and herpes simplex virus type 2 infection on lymphocyte and dendritic cell density in adult foreskins from Rakai, Uganda. J. Infect. Dis. 2011, 203, 602–609. [Google Scholar] [CrossRef]
- Prodger, J.L.; Gray, R.; Kigozi, G.; Nalugoda, F.; Galiwango, R.; Nehemiah, K.; Kakanga, M.; Hirbod, T.; Wawer, M.J.; Sewankambo, N.; et al. Impact of asymptomatic Herpes simplex virus-2 infection on T cell phenotype and function in the foreskin. AIDS 2012, 26, 1319–1322. [Google Scholar] [CrossRef]
- Tobian, A.A.; Grabowski, M.K.; Kigozi, G.; Redd, A.D.; Eaton, K.P.; Serwadda, D.; Cornish, T.C.; Nalugoda, F.; Watya, S.; Buwembo, D.; et al. Human Papillomavirus Clearance Among Males Is Associated With HIV Acquisition and Increased Dendritic Cell Density in the Foreskin. J. Infect. Dis. 2013, 207, 1713–1722. [Google Scholar] [CrossRef] [PubMed]
- Low, N.; Chersich, M.F.; Schmidlin, K.; Egger, M.; Francis, S.C.; van de Wijgert, J.H.; Hayes, R.J.; Baeten, J.M.; Brown, J.; Delany-Moretlwe, S.; et al. Intravaginal practices, bacterial vaginosis, and HIV infection in women: Individual participant data meta-analysis. PLoS Med. 2011, 8, e1000416. [Google Scholar] [CrossRef]
- Makumbi, F.E.; Ssempijja, V.; Sekasanvu, J.; Ssekubugu, R.; Kigozi, G.; Watya, S.; Nakigozi, G.; Serwadda, D.; Wawer, M.; Gray, R.H. Postcoital penile washing and the risk of HIV acquisition in uncircumcised men. AIDS 2016, 30, 1669–1673. [Google Scholar] [CrossRef]
- Van Damme, L.; Ramjee, G.; Alary, M.; Vuylsteke, B.; Chandeying, V.; Rees, H.; Sirivongrangson, P.; Mukenge-Tshibaka, L.; Ettiegne-Traore, V.; Uaheowitchai, C.; et al. Effectiveness of COL-1492, a nonoxynol-9 vaginal gel, on HIV-1 transmission in female sex workers: A randomised controlled trial. Lancet 2002, 360, 971–977. [Google Scholar] [CrossRef]
- Fichorova, R.N.; Bajpai, M.; Chandra, N.; Hsiu, J.G.; Spangler, M.; Ratnam, V.; Doncel, G.F. Interleukin (IL)-1, IL-6, and IL-8 predict mucosal toxicity of vaginal microbicidal contraceptives. Biol. Reprod. 2004, 71, 761–769. [Google Scholar] [CrossRef]
- Fichorova, R.N.; Tucker, L.D.; Anderson, D.J. The molecular basis of nonoxynol-9-induced vaginal inflammation and its possible relevance to human immunodeficiency virus type 1 transmission. J. Infect. Dis. 2001, 184, 418–428. [Google Scholar] [CrossRef]
- Adriaens, E.; Remon, J.P. Mucosal irritation potential of personal lubricants relates to product osmolality as detected by the slug mucosal irritation assay. Sex. Transm. Dis. 2008, 35, 512–516. [Google Scholar] [CrossRef]
- Dezzutti, C.S.; Brown, E.R.; Moncla, B.; Russo, J.; Cost, M.; Wang, L.; Uranker, K.; Kunjara Na Ayudhya, R.P.; Pryke, K.; Pickett, J.; et al. Is wetter better? An evaluation of over-the-counter personal lubricants for safety and anti-HIV-1 activity. PLoS ONE 2012, 7, e48328. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Shahabi, K.; Spitzer, R.; Farrugia, M.; Kaul, R.; Yudin, M. Cervico-vaginal inflammatory cytokine alterations after intrauterine contraceptive device insertion: A pilot study. PLoS ONE 2018, 13, e0207266. [Google Scholar] [CrossRef] [PubMed]
- Achilles, S.L.; Meyn, L.A.; Mhlanga, F.G.; Matubu, A.T.; Stoner, K.A.; Beamer, M.A.; Chirenje, Z.M.; Hillier, S.L. Zim CHIC: A cohort study of immune changes in the female genital tract associated with initiation and use of contraceptives. Am. J. Reprod. Immunol. 2020, 84, e13287. [Google Scholar] [CrossRef]
- Ortiz, M.E.; Croxatto, H.B. Copper-T intrauterine device and levonorgestrel intrauterine system: Biological bases of their mechanism of action. Contraception 2007, 75, S16–S30. [Google Scholar] [CrossRef]
- Achilles, S.L.; Austin, M.N.; Meyn, L.A.; Mhlanga, F.; Chirenje, Z.M.; Hillier, S.L. Impact of contraceptive initiation on vaginal microbiota. Am. J. Obstet. Gynecol. 2018, 218, 622.e1–622.e10. [Google Scholar] [CrossRef]
- Curtis, K.M.; Hannaford, P.C.; Rodriguez, M.I.; Chipato, T.; Steyn, P.S.; Kiarie, J.N. Hormonal contraception and HIV acquisition among women: An updated systematic review. BMJ Sex. Reprod. Health 2020, 46, 8–16. [Google Scholar] [CrossRef]
- Morrison, C.S.; Chen, P.L.; Kwok, C.; Baeten, J.M.; Brown, J.; Crook, A.M.; Van Damme, L.; Delany-Moretlwe, S.; Francis, S.C.; Friedland, B.A.; et al. Hormonal contraception and the risk of HIV acquisition: An individual participant data meta-analysis. PLoS Med. 2015, 12, e1001778. [Google Scholar] [CrossRef]
- Evidence for Contraceptive Options; HIV Outcomes (ECHO) Trial Consortium. HIV incidence among women using intramuscular depot medroxyprogesterone acetate, a copper intrauterine device, or a levonorgestrel implant for contraception: A randomised, multicentre, open-label trial. Lancet 2019, 394, 303–313. [Google Scholar] [CrossRef]
- Byrne, E.H.; Anahtar, M.N.; Cohen, K.E.; Moodley, A.; Padavattan, N.; Ismail, N.; Bowman, B.A.; Olson, G.S.; Mabhula, A.; Leslie, A.; et al. Association between injectable progestin-only contraceptives and HIV acquisition and HIV target cell frequency in the female genital tract in South African women: A prospective cohort study. Lancet Infect. Dis. 2016, 16, 441–448. [Google Scholar] [CrossRef]
- Tobian, A.A.; Serwadda, D.; Quinn, T.C.; Kigozi, G.; Gravitt, P.E.; Laeyendecker, O.; Charvat, B.; Ssempijja, V.; Riedesel, M.; Oliver, A.E.; et al. Male circumcision for the prevention of HSV-2 and HPV infections and syphilis. N. Engl. J. Med. 2009, 360, 1298–1309. [Google Scholar] [CrossRef]
- Tobian, A.A.; Gray, R.H. The medical benefits of male circumcision. JAMA 2011, 306, 1479–1480. [Google Scholar] [CrossRef][Green Version]
- Wawer, M.J.; Tobian, A.A.; Kigozi, G.; Kong, X.; Gravitt, P.E.; Serwadda, D.; Nalugoda, F.; Makumbi, F.; Ssempiija, V.; Sewankambo, N.; et al. Effect of circumcision of HIV-negative men on transmission of human papillomavirus to HIV-negative women: A randomised trial in Rakai, Uganda. Lancet 2011, 377, 209–218. [Google Scholar] [CrossRef]
- Gray, R.H.; Serwadda, D.; Kong, X.; Makumbi, F.; Kigozi, G.; Gravitt, P.E.; Watya, S.; Nalugoda, F.; Ssempijja, V.; Tobian, A.A.; et al. Male circumcision decreases acquisition and increases clearance of high-risk human papillomavirus in HIV-negative men: A randomized trial in Rakai, Uganda. J. Infect. Dis. 2010, 201, 1455–1462. [Google Scholar] [CrossRef]
- Galiwango, R.M.; Park, D.E.; Huibner, S.; Onos, A.; Aziz, M.; Roach, K.; Anok, A.; Nnamutete, J.; Isabirye, Y.; Wasswa, J.B.; et al. Immune milieu and microbiome of the distal urethra in Ugandan men: Impact of penile circumcision and implications for HIV susceptibility. Microbiome 2022, 10, 7. [Google Scholar] [CrossRef]
- McClelland, R.S.; Lingappa, J.R.; Srinivasan, S.; Kinuthia, J.; John-Stewart, G.C.; Jaoko, W.; Richardson, B.A.; Yuhas, K.; Fiedler, T.L.; Mandaliya, K.N.; et al. Evaluation of the association between the concentrations of key vaginal bacteria and the increased risk of HIV acquisition in African women from five cohorts: A nested case-control study. Lancet Infect. Dis. 2018, 18, 554–564. [Google Scholar] [CrossRef]
- Gosmann, C.; Anahtar, M.N.; Handley, S.A.; Farcasanu, M.; Abu-Ali, G.; Bowman, B.A.; Padavattan, N.; Desai, C.; Droit, L.; Moodley, A.; et al. Lactobacillus-Deficient Cervicovaginal Bacterial Communities Are Associated with Increased HIV Acquisition in Young South African Women. Immunity 2017, 46, 29–37. [Google Scholar] [CrossRef]
- Prodger, J.L.; Abraham, A.G.; Tobian, A.A.; Park, D.E.; Aziz, M.; Roach, K.; Gray, R.H.; Buchanan, L.; Kigozi, G.; Galiwango, R.M.; et al. Penile bacteria associated with HIV seroconversion, inflammation, and immune cells. JCI Insight 2021, 6, e147363. [Google Scholar] [CrossRef]
- Liu, C.M.; Prodger, J.L.; Tobian, A.A.R.; Abraham, A.G.; Kigozi, G.; Hungate, B.A.; Aziz, M.; Nalugoda, F.; Sariya, S.; Serwadda, D.; et al. Penile Anaerobic Dysbiosis as a Risk Factor for HIV Infection. MBio 2017, 8, e00996-17. [Google Scholar] [CrossRef]
- Valore, E.V.; Park, C.H.; Igreti, S.L.; Ganz, T. Antimicrobial components of vaginal fluid. Am. J. Obstet. Gynecol. 2002, 187, 561–568. [Google Scholar] [CrossRef]
- Mirmonsef, P.; Hotton, A.L.; Gilbert, D.; Gioia, C.J.; Maric, D.; Hope, T.J.; Landay, A.L.; Spear, G.T. Glycogen Levels in Undiluted Genital Fluid and Their Relationship to Vaginal pH, Estrogen, and Progesterone. PLoS ONE 2016, 11, e0153553. [Google Scholar] [CrossRef]
- Mirmonsef, P.; Spear, G.T. The barrier to HIV transmission provided by genital tract Lactobacillus colonization. Am. J. Reprod. Immunol. 2014, 71, 531–536. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. In vaginal fluid, bacteria associated with bacterial vaginosis can be suppressed with lactic acid but not hydrogen peroxide. BMC Infect. Dis. 2011, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal pH and microbicidal lactic acid when lactobacilli dominate the microbiota. PLoS ONE 2013, 8, e80074. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Luna, Y.; Yu, P.; Fan, H. Lactobacilli inactivate Chlamydia trachomatis through lactic acid but not H2O2. PLoS ONE 2014, 9, e107758. [Google Scholar] [CrossRef] [PubMed]
- Nardini, P.; Nahui Palomino, R.A.; Parolin, C.; Laghi, L.; Foschi, C.; Cevenini, R.; Vitali, B.; Marangoni, A. Lactobacillus crispatus inhibits the infectivity of Chlamydia trachomatis elementary bodies, in vitro study. Sci. Rep. 2016, 6, 29024. [Google Scholar] [CrossRef]
- Graver, M.A.; Wade, J.J. The role of acidification in the inhibition of Neisseria gonorrhoeae by vaginal lactobacilli during anaerobic growth. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 8. [Google Scholar] [CrossRef]
- Breshears, L.M.; Edwards, V.L.; Ravel, J.; Peterson, M.L. Lactobacillus crispatus inhibits growth of Gardnerella vaginalis and Neisseria gonorrhoeae on a porcine vaginal mucosa model. BMC Microbiol. 2015, 15, 276. [Google Scholar] [CrossRef]
- Edwards, V.L.; Smith, S.B.; McComb, E.J.; Tamarelle, J.; Ma, B.; Humphrys, M.S.; Gajer, P.; Gwilliam, K.; Schaefer, A.M.; Lai, S.K.; et al. The Cervicovaginal Microbiota-Host Interaction Modulates Chlamydia trachomatis Infection. mBio 2019, 10, e01548-19. [Google Scholar] [CrossRef]
- McKinnon, L.R.; Achilles, S.L.; Bradshaw, C.S.; Burgener, A.; Crucitti, T.; Fredricks, D.N.; Jaspan, H.B.; Kaul, R.; Kaushic, C.; Klatt, N.; et al. The Evolving Facets of Bacterial Vaginosis: Implications for HIV Transmission. AIDS Res. Hum. Retrovir. 2019, 35, 219–228. [Google Scholar] [CrossRef]
- Turpin, R.; Slopen, N.; Borgogna, J.C.; Yeoman, C.J.; He, X.; Miller, R.S.; Klebanoff, M.A.; Ravel, J.; Brotman, R.M. Perceived Stress and Molecular-BV in the NIH Longitudinal Study of Vaginal Flora. Am. J. Epidemiol. 2021, 190, 2374–2383. [Google Scholar] [CrossRef]
- France, M.T.; Ma, B.; Gajer, P.; Brown, S.; Humphrys, M.S.; Holm, J.B.; Waetjen, L.E.; Brotman, R.M.; Ravel, J. VALENCIA: A nearest centroid classification method for vaginal microbial communities based on composition. Microbiome 2020, 8, 166. [Google Scholar] [CrossRef]
- France, M.T.; Rutt, L.; Narina, S.; Arbaugh, S.; McComb, E.; Humphrys, M.S.; Ma, B.; Hayward, M.R.; Costello, E.K.; Relman, D.A.; et al. Complete Genome Sequences of Six Lactobacillus iners Strains Isolated from the Human Vagina. Microbiol. Resour. Announc. 2020, 9, e00234-20. [Google Scholar] [CrossRef]
- Ravel, J.; Brotman, R.M.; Gajer, P.; Ma, B.; Nandy, M.; Fadrosh, D.W.; Sakamoto, J.; Koenig, S.S.; Fu, L.; Zhou, X.; et al. Daily temporal dynamics of vaginal microbiota before, during and after episodes of bacterial vaginosis. Microbiome 2013, 1, 29. [Google Scholar] [CrossRef]
- O’Hanlon, D.E.; Gajer, P.; Brotman, R.M.; Ravel, J. Asymptomatic Bacterial Vaginosis Is Associated With Depletion of Mature Superficial Cells Shed From the Vaginal Epithelium. Front. Cell Infect. Microbiol. 2020, 10, 106. [Google Scholar] [CrossRef]
- Zevin, A.S.; Xie, I.Y.; Birse, K.; Arnold, K.; Romas, L.; Westmacott, G.; Novak, R.M.; McCorrister, S.; McKinnon, L.R.; Cohen, C.R.; et al. Microbiome Composition and Function Drives Wound-Healing Impairment in the Female Genital Tract. PLoS Pathog. 2016, 12, e1005889. [Google Scholar] [CrossRef]
- Tamarelle, J.; Thiebaut, A.C.M.; de Barbeyrac, B.; Bebear, C.; Ravel, J.; Delarocque-Astagneau, E. The vaginal microbiota and its association with human papillomavirus, Chlamydia trachomatis, Neisseria gonorrhoeae and Mycoplasma genitalium infections: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2019, 25, 35–47. [Google Scholar] [CrossRef]
- Joag, V.; Obila, O.; Gajer, P.; Scott, M.C.; Dizzell, S.; Humphrys, M.; Shahabi, K.; Huibner, S.; Shannon, B.; Tharao, W.; et al. Impact of Standard Bacterial Vaginosis Treatment on the Genital Microbiota, Immune Milieu, and Ex Vivo Human Immunodeficiency Virus Susceptibility. Clin. Infect. Dis. 2019, 68, 1675–1683. [Google Scholar] [CrossRef]
- Wiesenfeld, H.C.; Hillier, S.L.; Krohn, M.A.; Landers, D.V.; Sweet, R.L. Bacterial vaginosis is a strong predictor of Neisseria gonorrhoeae and Chlamydia trachomatis infection. Clin. Infect. Dis. 2003, 36, 663–668. [Google Scholar] [CrossRef]
- Brotman, R.M.; Klebanoff, M.A.; Nansel, T.R.; Yu, K.F.; Andrews, W.W.; Zhang, J.; Schwebke, J.R. Bacterial vaginosis assessed by gram stain and diminished colonization resistance to incident gonococcal, chlamydial, and trichomonal genital infection. J. Infect. Dis. 2010, 202, 1907–1915. [Google Scholar] [CrossRef]
- Leitich, H.; Bodner-Adler, B.; Brunbauer, M.; Kaider, A.; Egarter, C.; Husslein, P. Bacterial vaginosis as a risk factor for preterm delivery: A meta-analysis. Am. J. Obstet. Gynecol. 2003, 189, 139–147. [Google Scholar] [CrossRef]
- Eschenbach, D.A.; Hillier, S.; Critchlow, C.; Stevens, C.; DeRouen, T.; Holmes, K.K. Diagnosis and clinical manifestations of bacterial vaginosis. Am. J. Obstet. Gynecol. 1988, 158, 819–828. [Google Scholar] [CrossRef]
- Redelinghuys, M.J.; Geldenhuys, J.; Jung, H.; Kock, M.M. Bacterial Vaginosis: Current Diagnostic Avenues and Future Opportunities. Front. Cell Infect. Microbiol. 2020, 10, 354. [Google Scholar] [CrossRef]
- Schwebke, J.R.; Hillier, S.L.; Sobel, J.D.; McGregor, J.A.; Sweet, R.L. Validity of the vaginal gram stain for the diagnosis of bacterial vaginosis. Obstet. Gynecol. 1996, 88, 573–576. [Google Scholar] [CrossRef]
- Anahtar, M.N.; Byrne, E.H.; Doherty, K.E.; Bowman, B.A.; Yamamoto, H.S.; Soumillon, M.; Padavattan, N.; Ismail, N.; Moodley, A.; Sabatini, M.E.; et al. Cervicovaginal bacteria are a major modulator of host inflammatory responses in the female genital tract. Immunity 2015, 42, 965–976. [Google Scholar] [CrossRef]
- Liu, C.M.; Hungate, B.A.; Tobian, A.A.; Ravel, J.; Prodger, J.L.; Serwadda, D.; Kigozi, G.; Galiwango, R.M.; Nalugoda, F.; Keim, P.; et al. Penile Microbiota and Female Partner Bacterial Vaginosis in Rakai, Uganda. MBio 2015, 6, e00589. [Google Scholar] [CrossRef]
- Liu, C.M.; Hungate, B.A.; Tobian, A.A.; Serwadda, D.; Ravel, J.; Lester, R.; Kigozi, G.; Aziz, M.; Galiwango, R.M.; Nalugoda, F.; et al. Male circumcision significantly reduces prevalence and load of genital anaerobic bacteria. MBio 2013, 4, e00076. [Google Scholar] [CrossRef]
- Bailey, R.; Moses, S.; Parker, C.; Agot, K.; Maclean, I.; Krieger, J.; Williams, C.; Campbell, R.; Ndinyaachola, J. Male circumcision for HIV prevention in young men in Kisumu, Kenya: A randomised controlled trial. Lancet 2007, 369, 643–656. [Google Scholar] [CrossRef]
- Auvert, B.; Taljaard, D.; Lagarde, E.; Sobngwi-Tambekou, J.; Sitta, R.; Puren, A. Randomized, controlled intervention trial of male circumcision for reduction of HIV infection risk: The ANRS 1265 Trial. PLoS Med. 2005, 2, e298. [Google Scholar] [CrossRef]
- Armstrong, E.; Hemmerling, A.; Miller, S.; Burke, K.E.; Newmann, S.J.; Morris, S.R.; Reno, H.; Huibner, S.; Kulikova, M.; Liu, R.; et al. Metronidazole treatment rapidly reduces genital inflammation through effects on bacterial vaginosis-associated bacteria rather than lactobacilli. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaul, R.; Liu, C.M.; Park, D.E.; Galiwango, R.M.; Tobian, A.A.R.; Prodger, J.L. The Penis, the Vagina and HIV Risk: Key Differences (Aside from the Obvious). Viruses 2022, 14, 1164. https://doi.org/10.3390/v14061164
Kaul R, Liu CM, Park DE, Galiwango RM, Tobian AAR, Prodger JL. The Penis, the Vagina and HIV Risk: Key Differences (Aside from the Obvious). Viruses. 2022; 14(6):1164. https://doi.org/10.3390/v14061164
Chicago/Turabian StyleKaul, Rupert, Cindy M. Liu, Daniel E. Park, Ronald M. Galiwango, Aaron A. R. Tobian, and Jessica L. Prodger. 2022. "The Penis, the Vagina and HIV Risk: Key Differences (Aside from the Obvious)" Viruses 14, no. 6: 1164. https://doi.org/10.3390/v14061164
APA StyleKaul, R., Liu, C. M., Park, D. E., Galiwango, R. M., Tobian, A. A. R., & Prodger, J. L. (2022). The Penis, the Vagina and HIV Risk: Key Differences (Aside from the Obvious). Viruses, 14(6), 1164. https://doi.org/10.3390/v14061164