
Isolation and Characterization of a Novel Cyanophage Encoding Multiple Auxiliary Metabolic Genes

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cyanophage Isolation, Purification, and Enrichment
2.2. Specificity Test
2.3. Transmission Electron Microscopic Analysis
2.4. DNA Extraction
2.5. Genomic Analysis
2.6. Mass Spectrometry-Based Proteomics
2.7. NanoLC-ESI-MS/MS Analysis
3. Results and Discussion
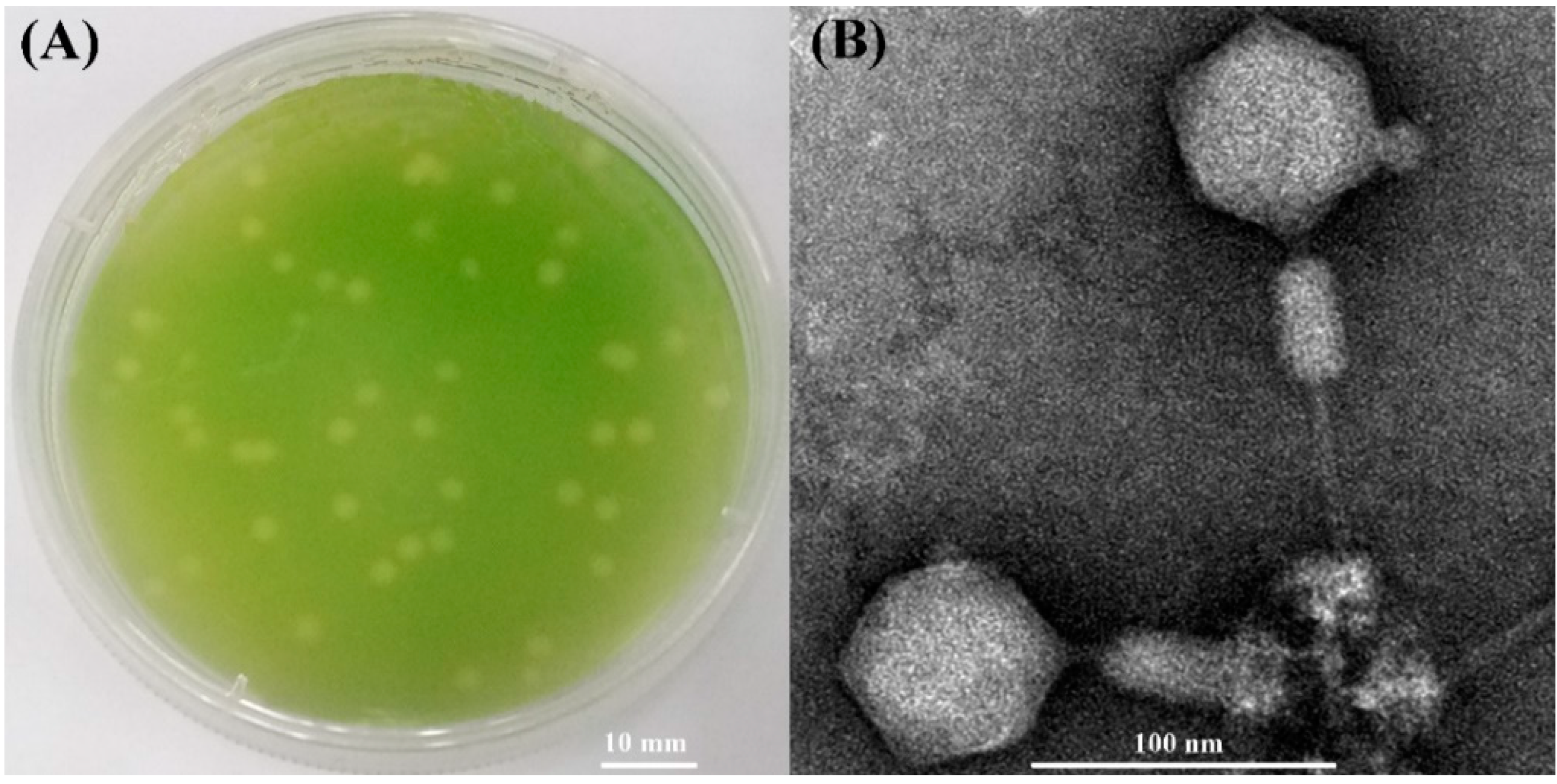
3.1. Morphology and Host Range of S-SZBM1
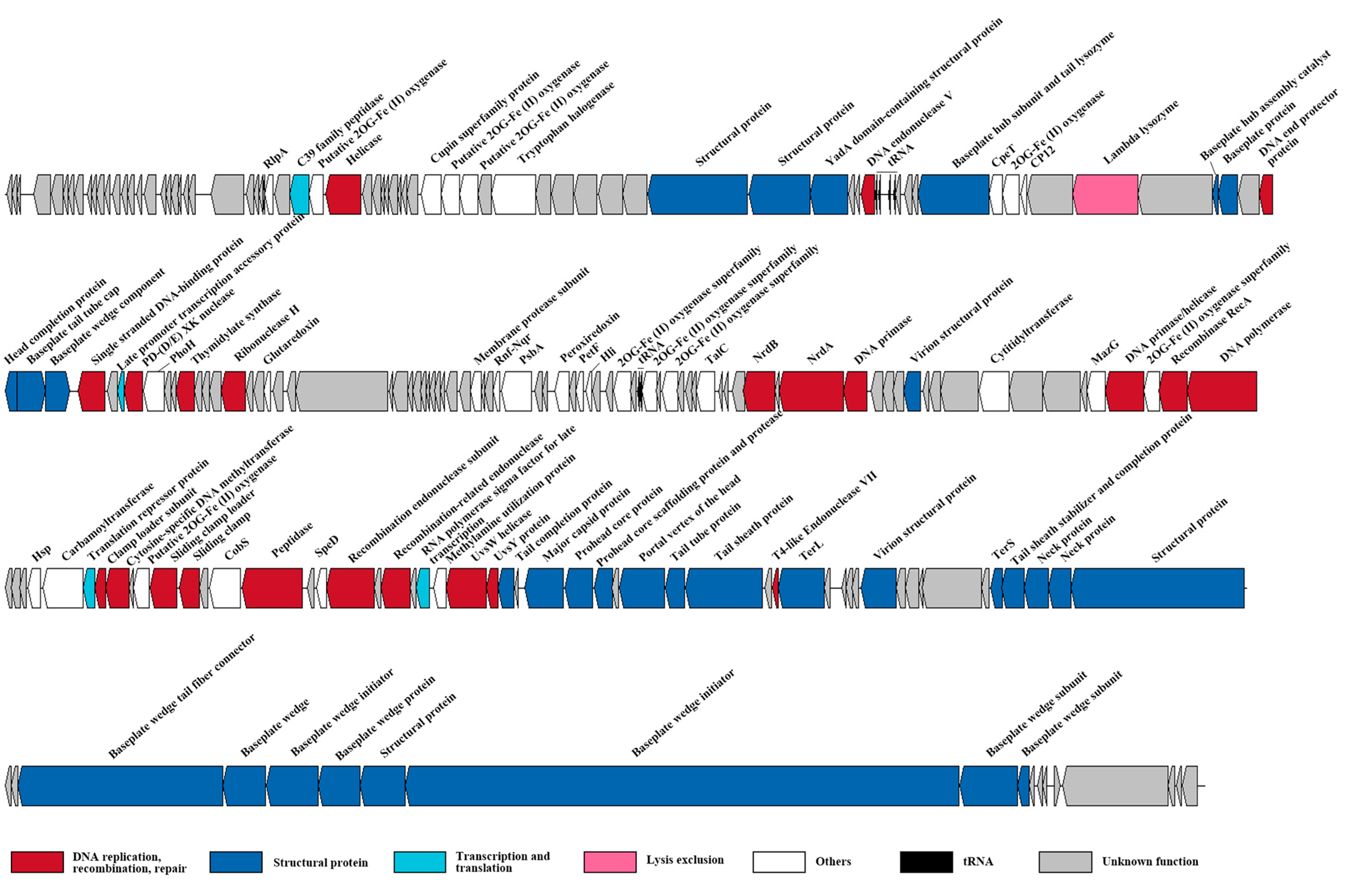
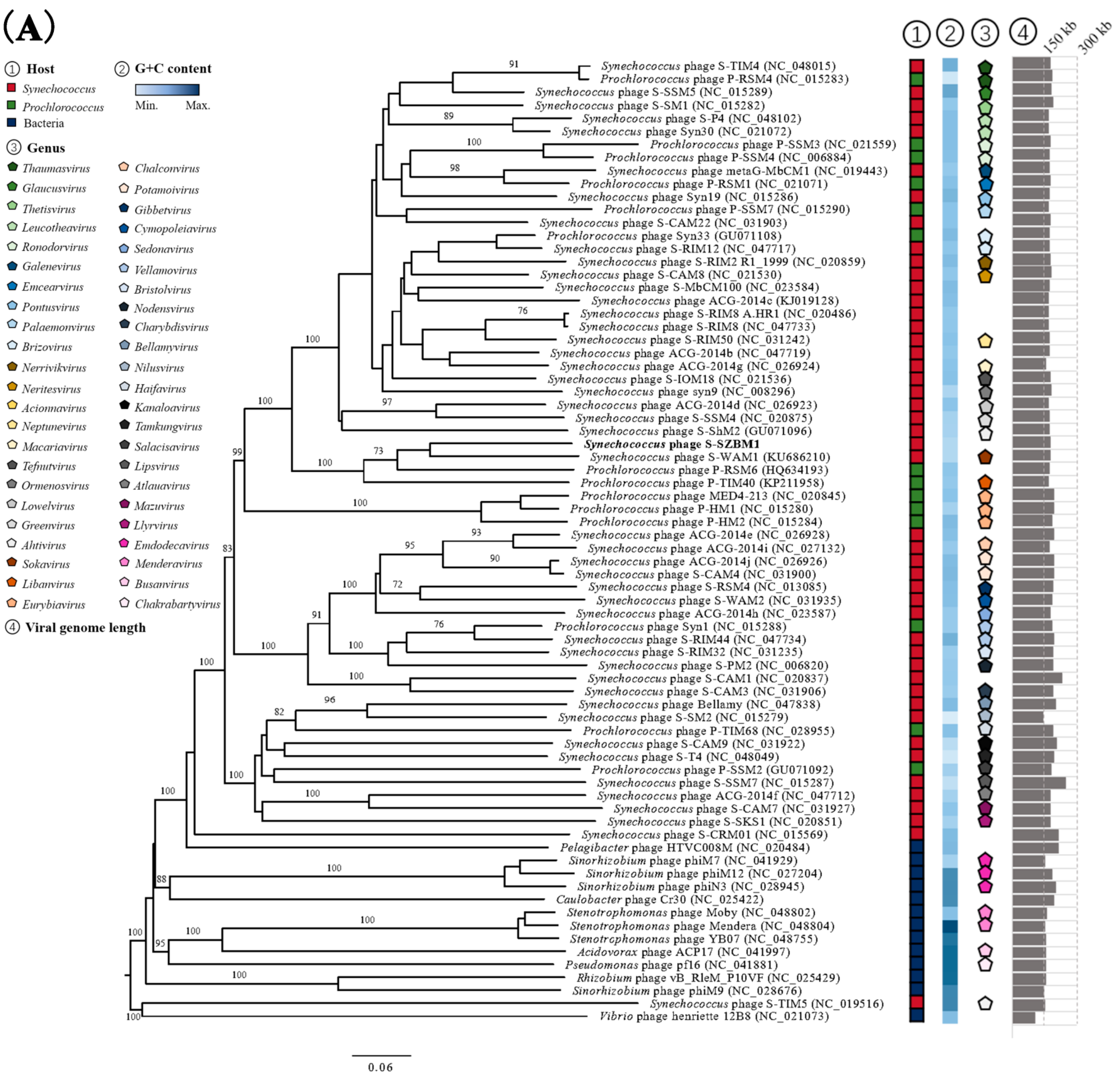
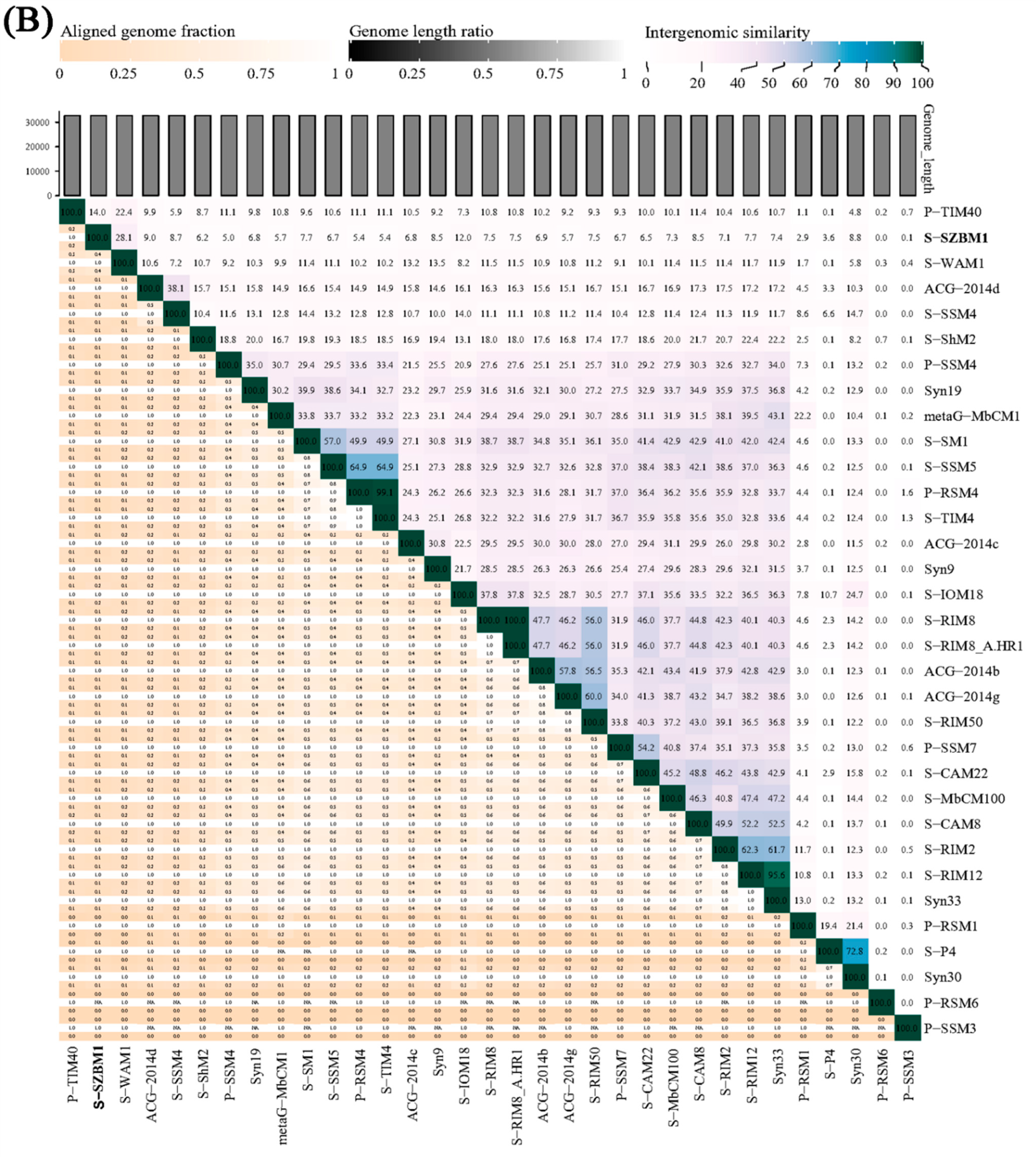
3.2. Genome Features and Taxonomy of S-SZBM1
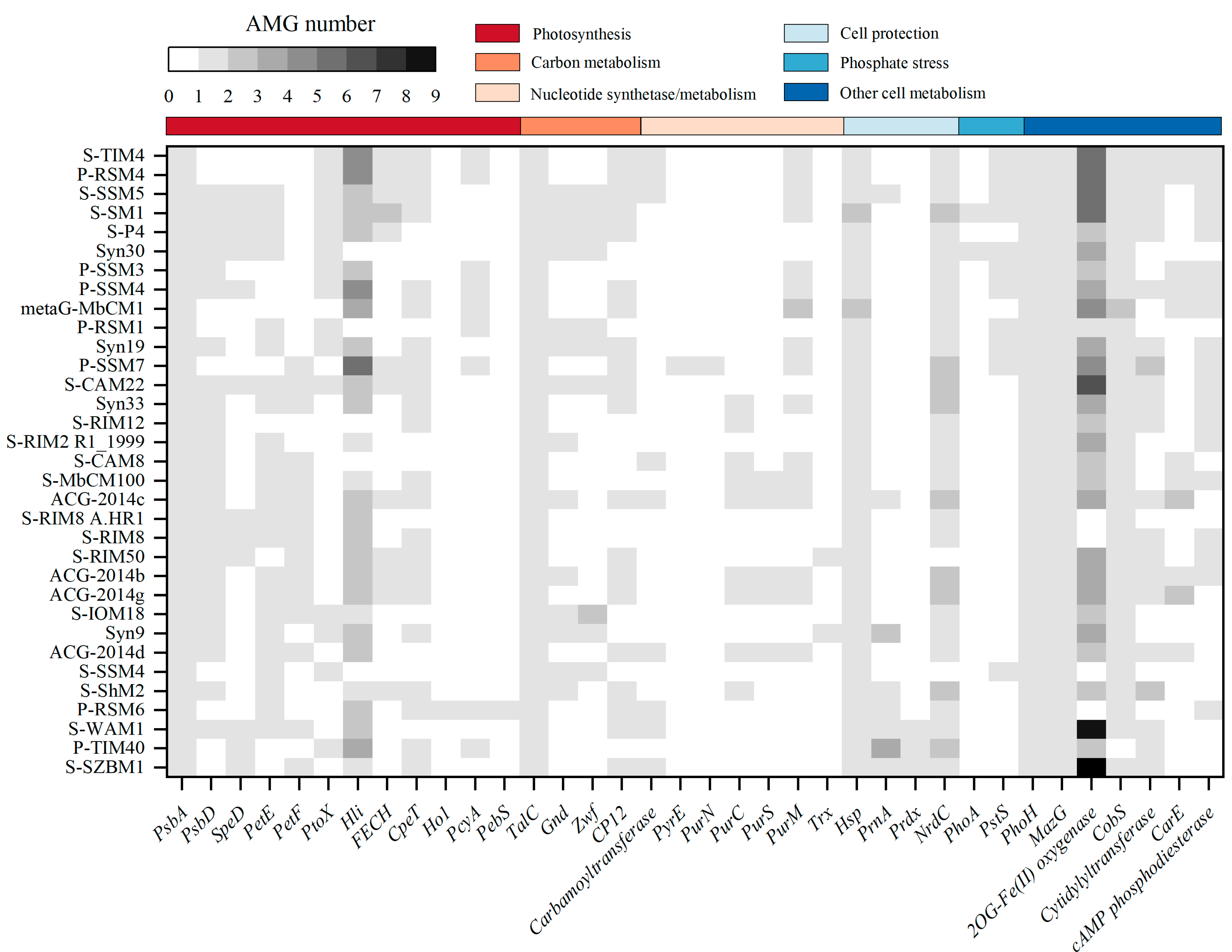
3.3. AMGs Encoded by S-SZBM1 Re-Program Phage–Host Interactions
3.4. Cyanophages Structural Proteome
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scanlan, D.J.; Ostrowski, M.; Mazard, S.; Dufresne, A.; Garczarek, L.; Hess, W.R.; Post, A.F.; Hagemann, M.; Paulsen, I.; Partensky, F. Ecological genomics of marine picocyanobacteria. Microbiol. Mol. Biol. Rev. 2009, 73, 249–299. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Milo, R. The biomass composition of the oceans: A blueprint of our blue planet. Cell 2019, 179, 1451–1454. [Google Scholar] [CrossRef]
- Proctor, L.M.; Fuhrman, J.A. Viral mortality of marine bacteria and cyanobacteria. Nature 1990, 343, 60–62. [Google Scholar] [CrossRef]
- Suttle, C.A. Marine viruses-major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef]
- Puxty, R.J.; Evans, D.J.; Millard, A.D.; Scanlan, D.J. Energy limitation of cyanophage development: Implications for marine carbon cycling. ISME J. 2018, 12, 1273–1286. [Google Scholar] [CrossRef]
- Buckling, A.; Rainey, P.B. Antagonistic coevolution between a bacterium and a bacteriophage. Proc. Biol. Sci. 2002, 269, 931–936. [Google Scholar] [CrossRef]
- Sabehi, G.; Shaulov, L.; Silver, D.H.; Yanai, I.; Harel, A.; Lindell, D. A novel lineage of myoviruses infecting cyanobacteria is widespread in the oceans. Proc. Natl. Acad. Sci. USA 2012, 109, 2037–2042. [Google Scholar] [CrossRef]
- Marston, M.F.; Pierciey, F.J., Jr.; Shepard, A.; Gearin, G.; Qi, J.; Yandava, C.; Schuster, S.C.; Henn, M.R.; Martiny, J.B. Rapid diversification of coevolving marine Synechococcus and a virus. Proc. Natl. Acad. Sci. USA 2012, 109, 4544–4549. [Google Scholar] [CrossRef]
- Brum, J.R.; Ignacio-Espinoza, J.C.; Roux, S.; Doulcier, G.; Acinas, S.G.; Alberti, A.; Chaffron, S.; Cruaud, C.; Vargas, C.D.; Gaso, J.M.; et al. Patterns and ecological drivers of ocean viral communities. Science 2015, 348, 1261498. [Google Scholar] [CrossRef] [PubMed]
- Roux, S.; Brum, J.R.; Dutilh, B.E.; Sunagawa, S.; Duhaime, M.B.; Loy, A.; Poulos, B.T.; Solonenko, N.; Lara, E.; Poulain, J.; et al. Ecogenomics and potential biogeochemical impacts of globally abundant ocean viruses. Nature 2016, 537, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.; Sullivan, M.B.; Johnson, Z.I.; Tolonen, A.C.; Rohwer, F.; Chisholm, S.W. Transfer of photosynthesis genes to and from Prochlorococcus viruses. Proc. Natl. Acad. Sci. USA 2004, 101, 11013–11018. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.B.; Coleman, M.L.; Weigele, P.; Rohwer, F.; Chisholm, S.W. Three Prochlorococcus cyanophage genomes: Signature features and ecological interpretations. PLOS Biol. 2005, 3, e144. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M.; Bonnain, C.; Malki, K.; Sawaya, N.A. Phage puppet masters of the marine microbial realm. Nat. Microbiol. 2018, 3, 754–766. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Chisholm, S.W. Marine viruses exploit their host’s two-component regulatory system in response to resource limitation. Curr. Biol. 2012, 22, 124–128. [Google Scholar] [CrossRef]
- Thompson, L.R.; Zeng, Q.; Kelly, L.; Huang, K.H.; Singer, A.U.; Stubbe, J.; Chisholm, S.W. Phage auxiliary metabolic genes and the redirection of cyanobacterial host carbon metabolism. Proc. Natl. Acad. Sci. USA 2011, 108, E757–E764. [Google Scholar] [CrossRef]
- Crummett, L.T.; Puxty, R.J.; Weihe, C.; Marston, M.F.; Martiny, J.B. The genomic content and context of auxiliary metabolic genes in marine cyanomyoviruses. Virology 2016, 499, 219–229. [Google Scholar] [CrossRef]
- Hurwitz, B.L.; U’Ren, J.M. Viral metabolic reprogramming in marine ecosystems. Curr. Opin. Microbiol. 2016, 31, 161–168. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, R.; Wang, N.; Cai, L.; Tong, Y.; Sun, Q.; Chen, F.; Jiao, N. Novel phage-host interactions and evolution as revealed by a cyanomyovirus isolated from an estuarine environment. Environ. Microbiol. 2018, 20, 2974–2989. [Google Scholar] [CrossRef]
- Lindell, D.; Jaffe, J.D.; Coleman, M.L.; Futschik, M.E.; Axmann, I.M.; Rector, T.; Kettler, G.; Sullivan, M.B.; Steen, R.; Hess, W.R.; et al. Genome-wide expression dynamics of a marine virus and host reveal features of co-evolution. Nature 2007, 449, 83–86. [Google Scholar] [CrossRef]
- Wang, K.; Wommack, K.E.; Chen, F. Abundance and distribution of Synechococcus spp. and cyanophages in the Chesapeake Bay. Appl. Environ. Microbiol. 2011, 77, 7459–7468. [Google Scholar] [CrossRef]
- Marsan, D.; Wommack, K.E.; Ravel, J.; Chen, F. Draft genome sequence of Synechococcus sp. strain CB0101, isolated from the Chesapeake Bay estuary. Genome Announc. 2014, 2, e01111–e01113. [Google Scholar] [CrossRef]
- Marsan, D.; Place, A.; Fucich, D.; Chen, F. Toxin-antitoxin systems in estuarine Synechococcus strain CB0101 and their transcriptomic responses to environmental stressors. Front. Microbiol. 2017, 8, 1213. [Google Scholar] [CrossRef]
- Chen, F.; Wang, K.; Kan, J.; Bachoon, D.S.; Lu, J.; Lau, S.; Campbell, L. Phylogenetic diversity of Synechococcus in the Chesapeake Bay revealed by ribulose-1, 5-bisphosphate carboxylase-oxygenase (RuBisCO) large subunit gene (rbcL) sequences. Aquat. Microb. Ecol. 2004, 36, 153–164. [Google Scholar] [CrossRef][Green Version]
- Zhao, Z.; Gonsior, M.; Luek, J.; Timko, S.; Ianiri, H.; Hertkorn, N.; Schmitt-Kopplin, P.; Fang, X.; Zeng, Q.; Jiao, N.; et al. Picocyanobacteria and deep-ocean fluorescent dissolved organic matter share similar optical properties. Nat. Commun. 2017, 8, 15284. [Google Scholar] [CrossRef]
- Wilson, W.H.; Joint, I.R.; Carr, N.G.; Mann, N.H. Isolation and molecular characterization of five marine cyanophages propagated on Synechococcus sp. strain WH7803. Appl. Environ. Microbiol. 1993, 59, 3736–3743. [Google Scholar] [CrossRef]
- Lawrence, J.E.; Steward, G.F. Purification of viruses by centrifugation. In Manual of Aquatic Viral Ecology; Wilhelm, S.W., Weinbauer, M.G., Suttle, C.A., Eds.; American Society of Limnology and Oceanography: Waco, TX, USA, 2010; pp. 166–181. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Anantharaman, K.; Duhaime, M.B.; Breier, J.A.; Wendt, K.A.; Toner, B.M.; Dick, G.J. Sulfur oxidation genes in diverse deep-sea viruses. Science 2014, 344, 757–760. [Google Scholar] [CrossRef]
- Albertsen, M.; Hugenholtz, P.; Skarshewski, A.; Nielsen, K.L.; Tyson, G.W.; Nielsen, P.H. Genome sequences of rare, uncultured bacteria obtained by differential coverage binning of multiple metagenomes. Nat. Biotechnol. 2013, 31, 533–538. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.; Li, S.; An, X.; Pei, G.; Huang, Y.; Fan, H.; Mi, Z.; Zhang, Z.; Wang, W.; et al. A novel termini analysis theory using HTS data alone for the identification of Enterococcus phage EF4-like genome termini. BMC Genom. 2015, 16, 414. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Bin Jang, H.; Bolduc, B.; Zablocki, O.; Kuhn, J.H.; Roux, S.; Adriaenssens, E.M.; Brister, J.R.; Kropinski, A.M.; Krupovic, M.; Lavigne, R.; et al. Taxonomic assignment of uncultivated prokaryotic virus genomes is enabled by gene-sharing networks. Nat. Biotechnol. 2019, 37, 632–639. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Goker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC-A novel tool to calculate the intergenomic similarities of prokaryote-infecting viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Jiang, D.; Lang, X.; Tian, Z.; Wang, T. Considerable model—Data mismatch in temperature over China during the mid-holocene: Results of PMIP simulations. J. Clim. 2012, 25, 4135–4153. [Google Scholar] [CrossRef]
- Jiang, Z.; Kumar, M.; Padula, M.P.; Pernice, M.; Kahlke, T.; Kim, M.; Ralph, P.J. Development of an efficient protein extraction method compatible with LC-MS/MS for proteome mapping in two Australian seagrasses Zostera muelleri and Posidonia australis. Front. Plant Sci. 2017, 8, 1416. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Y.; Arellano, S.M.; Xiao, K.; Qian, P.Y. Comparative proteome and phosphoproteome analyses during cyprid development of the barnacle Balanus (=Amphibalanus) amphitrite. J. Proteome Res. 2010, 9, 3146–3157. [Google Scholar] [CrossRef]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling expedition: Northwest Atlantic through eastern tropical Pacific. PLOS Biol. 2007, 5, e77. [Google Scholar] [CrossRef]
- Millard, A.D.; Zwirglmaier, K.; Downey, M.J.; Mann, N.H.; Scanlan, D.J. Comparative genomics of marine cyanomyoviruses reveals the widespread occurrence of Synechococcus host genes localized to a hyperplastic region: Implications for mechanisms of cyanophage evolution. Environ. Microbiol. 2009, 11, 2370–2387. [Google Scholar] [CrossRef]
- Jiang, T.; Guo, C.; Wang, M.; Wang, M.; Zhang, X.; Liu, Y.; Liang, Y.; Jiang, Y.; He, H.; Shao, H.; et al. Genome analysis of two novel Synechococcus phages that lack common auxiliary metabolic genes: Possible reasons and ecological insights by comparative analysis of cyanomyoviruses. Viruses 2020, 12, 800. [Google Scholar] [CrossRef]
- Sullivan, M.B.; Huang, K.H.; Ignacio-Espinoza, J.C.; Berlin, A.M.; Kelly, L.; Weigele, P.R.; DeFrancesco, A.S.; Kern, S.E.; Thompson, L.R.; Young, S.; et al. Genomic analysis of oceanic cyanobacterial myoviruses compared with T4-like myoviruses from diverse hosts and environments. Environ. Microbiol. 2010, 12, 3035–3056. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Wang, K.; Chen, F. Prevalence of highly host-specific cyanophages in the estuarine environment. Environ. Microbiol. 2008, 10, 300–312. [Google Scholar] [CrossRef]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A roadmap for genome-based phage taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Rosenwasser, S.; Ziv, C.; Creveld, S.G.V.; Vardi, A. Virocell metabolism: Metabolic innovations during host-virus interactions in the ocean. Trends Microbiol. 2016, 24, 821–832. [Google Scholar] [CrossRef]
- Zimmerman, A.E.; Howard-Varona, C.; Needham, D.M.; John, S.G.; Worden, A.Z.; Sullivan, M.B.; Waldbauer, J.R.; Coleman, M.L. Metabolic and biogeochemical consequences of viral infection in aquatic ecosystems. Nat. Rev. Microbiol. 2019, 18, 21–34. [Google Scholar] [CrossRef]
- Huang, X.; Jiao, N.; Zhang, R. The genomic content and context of auxiliary metabolic genes in roseophages. Environ. Microbiol. 2021, 23, 3743–3757. [Google Scholar] [CrossRef]
- Sullivan, M.B.; Lindell, D.; Lee, J.A.; Thompson, L.R.; Bielawski, J.P.; Chisholm, S.W. Prevalence and evolution of core photosystem II genes in marine cyanobacterial viruses and their hosts. PLoS Biol. 2006, 4, e234. [Google Scholar] [CrossRef]
- Zhao, Y.; Qin, F.; Zhang, R.; Giovannoni, S.J.; Zhang, Z.; Sun, J.; Du, S.; Rensing, C. Pelagiphages in the Podoviridae family integrate into host genomes. Environ. Microbiol. 2019, 21, 1989–2001. [Google Scholar] [CrossRef] [PubMed]
- Mann, N.H.; Cook, A.; Millard, A.; Bailey, S.; Clokie, M. Bacterial photosynthesis genes in a virus. Nature 2003, 424, 741. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.; Jaffe, J.D.; Johnson, Z.I.; Church, G.M.; Chisholm, S.W. Photosynthesis genes in marine viruses yield proteins during host infection. Nature 2005, 438, 86–89. [Google Scholar] [CrossRef]
- Puxty, R.J.; Millard, A.D.; Evans, D.J.; Scanlan, D.J. Viruses inhibit CO2 fixation in the most abundant phototrophs on earth. Curr. Biol. 2016, 26, 1585–1589. [Google Scholar] [CrossRef]
- Chenard, C.; Suttle, C.A. Phylogenetic diversity of sequences of cyanophage photosynthetic gene psbA in marine and freshwaters. Appl. Environ. Microbiol. 2008, 74, 5317–5324. [Google Scholar] [CrossRef]
- Puxty, R.J.; Millard, A.D.; Evans, D.J.; Scanlan, D.J. Shedding new light on viral photosynthesis. Photosynth. Res. 2015, 126, 71–97. [Google Scholar] [CrossRef]
- Clokie, M.R.; Mann, N.H. Marine cyanophages and light. Environ. Microbiol. 2006, 8, 2074–2082. [Google Scholar] [CrossRef]
- Yu, T.Y.; Schaefer, J. REDOR NMR characterization of DNA packaging in bacteriophage T4. J. Mol. Biol. 2008, 382, 1031–1042. [Google Scholar] [CrossRef]
- He, Q.; Dolganov, N.; Bjorkman, O.; Grossman, A.R. The high light-inducible polypeptides in Synechocystis PCC6803. J. Biol. Chem. 2001, 276, 306–314. [Google Scholar] [CrossRef]
- Dammeyer, T.; Bagby, S.C.; Sullivan, M.B.; Chisholm, S.W.; Frankenberg-Dinkel, N. Efficient phage-mediated pigment biosynthesis in oceanic cyanobacteria. Curr. Biol. 2008, 18, 442–448. [Google Scholar] [CrossRef]
- Jacobson, T.B.; Callaghan, M.M.; Amador-Noguez, D. Hostile takeover: How viruses reprogram prokaryotic metabolism. Annu. Rev. Microbiol. 2021, 75, 515–539. [Google Scholar] [CrossRef] [PubMed]
- McHaourab, H.S.; Godar, J.A.; Stewart, P.L. Structure and mechanism of protein stability sensors: Chaperone activity of small heat shock proteins. Biochemistry 2009, 48, 3828–3837. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Wage, T.; Hohaus, K.; Hölzer, M.; Eichhorn, E.; Pée, K.-H.V. Purification and partial characterization of tryptophan 7-halogenase (PrnA) from Pseudomonas fluorescens. Angew. Chem. Int. Ed. 2000, 39, 2300–2302. [Google Scholar] [CrossRef]
- Weigele, P.R.; Pope, W.H.; Pedulla, M.L.; Houtz, J.M.; Smith, A.L.; Conway, J.F.; King, J.; Hatfull, G.F.; Lawrence, J.G.; Hendrix, R.W. Genomic and structural analysis of Syn9, a cyanophage infecting marine Prochlorococcus and Synechococcus. Environ. Microbiol. 2007, 9, 1675–1695. [Google Scholar] [CrossRef]
- Clokie, M.R.; Millard, A.D.; Mann, N.H. T4 genes in the marine ecosystem: Studies of the T4-like cyanophages and their role in marine ecology. Virol. J. 2010, 7, 291. [Google Scholar] [CrossRef]
- Sharon, I.; Battchikova, N.; Aro, E.M.; Giglione, C.; Meinnel, T.; Glaser, F.; Pinter, R.Y.; Breitbart, M.; Rohwer, F.; Beja, O. Comparative metagenomics of microbial traits within oceanic viral communities. ISME J. 2011, 5, 1178–1190. [Google Scholar] [CrossRef]
- Kelly, L.; Ding, H.; Huang, K.H.; Osburne, M.S.; Chisholm, S.W. Genetic diversity in cultured and wild marine cyanomyoviruses reveals phosphorus stress as a strong selective agent. ISME J. 2013, 7, 1827–1841. [Google Scholar] [CrossRef]
- Goldsmith, D.B.; Crosti, G.; Dwivedi, B.; McDaniel, L.D.; Varsani, A.; Suttle, C.A.; Weinbauer, M.G.; Sandaa, R.A.; Breitbart, M. Development of phoH as a novel signature gene for assessing marine phage diversity. Appl. Environ. Microbiol. 2011, 77, 7730–7739. [Google Scholar] [CrossRef]
- Santos-Beneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef]
- Bryan, M.J.; Burroughs, N.J.; Spence, E.M.; Clokie, M.R.; Mann, N.H.; Bryan, S.J. Evidence for the intense exchange of mazG in marine cyanophages by horizontal gene transfer. PLoS ONE 2008, 3, e2048. [Google Scholar] [CrossRef]
- Rihtman, B.; Bowman-Grahl, S.; Millard, A.; Corrigan, R.M.; Clokie, M.R.J.; Scanlan, D.J. Cyanophage MazG is a pyrophosphohydrolase but unable to hydrolyse magic spot nucleotides. Environ. Microbiol. Rep. 2019, 11, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Weynberg, K.D.; Allen, M.J.; Gilg, I.C.; Scanlan, D.J.; Wilson, W.H. Genome sequence of Ostreococcus tauri virus OtV-2 throws light on the role of picoeukaryote niche separation in the ocean. J. Virol. 2011, 85, 4520–4529. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Allen, L.Z.; Palenik, B. Diversity and genome dynamics of marine cyanophages using metagenomic analyses. Environ. Microbiol. Rep. 2014, 6, 583–594. [Google Scholar] [CrossRef]
- Maggio-Hall, L.A.; Claas, K.R.; Escalante-Semerena, J.C. The last step in coenzyme B (12) synthesis is localized to the cell membrane in bacteria and archaea. Microbiology 2004, 150, 1385–1395. [Google Scholar] [CrossRef]
- Gao, E.B.; Huang, Y.; Ning, D. Metabolic genes within cyanophage genomes: Implications for diversity and evolution. Genes 2016, 7, 80. [Google Scholar] [CrossRef]
- Ignacio-Espinoza, J.C.; Sullivan, M.B. Phylogenomics of T4 cyanophages: Lateral gene transfer in the ’core’ and origins of host genes. Environ. Microbiol. 2012, 14, 2113–2126. [Google Scholar] [CrossRef]
- Doron, S.; Fedida, A.; Hernandez-Prieto, M.A.; Sabehi, G.; Karunker, I.; Stazic, D.; Feingersch, R.; Steglich, C.; Futschik, M.; Lindell, D.; et al. Transcriptome dynamics of a broad host-range cyanophage and its hosts. ISME J. 2015, 10, 1437–1455. [Google Scholar] [CrossRef]
- Pattridge, K.A.; Weber, C.H.; Friesen, J.A.; Sanker, S.; Kent, C.; Ludwig, M.L. Glycerol-3-phosphate cytidylyltransferase: Structural changes induced by binding of CDP-glycerol and the role of lysine residues in catalysis. J. Biol. Chem. 2003, 278, 51863–51871. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, F.; Chu, X.; Zhang, H.; Luo, H.; Qin, F.; Zhai, Z.; Yang, M.; Sun, J.; Zhao, Y. Diverse, abundant, and novel viruses infecting the marine Roseobacter RCA lineage. Msystems 2019, 4, e00494. [Google Scholar] [CrossRef]
- Pooley, H.; Abellan, F.; Karamata, D. A conditional-lethal mutant of Baciffus subtifis 168 with a thermosensitive glycerol-3-phosphate cytidylyltransferase, an enzyme specific for the synthesis of the major wall teichoic acid. J. Gen. Microbiol. 1991, 137, 921–928. [Google Scholar] [CrossRef]
- Mesyanzhinov, V.V.; Leiman, P.G.; Kostyuchenko, V.A.; Kurochkina, L.P.; Miroshnikov, K.A.; Sykilinda, N.N.; Shneider, M.M. Molecular architecture of bacteriophage T4. Biochemistry 2004, 69, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Fokine, A.; Rossmann, M.J.S. Common evolutionary origin of procapsid proteases, phage tail tubes, and tubes of bacterial type VI secretion systems. Structure 2016, 24, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Fokine, A.; Leiman, P.G.; Shneider, M.M.; Ahvazi, B.; Boeshans, K.M.; Steven, A.C.; Black, L.W.; Mesyanzhinov, V.V.; Rossmann, M.G. Structural and functional similarities between the capsid proteins of bacteriophages T4 and HK97 point to a common ancestry. Proc. Natl. Acad. Sci. USA 2005, 102, 7163–7168. [Google Scholar] [CrossRef] [PubMed]
- Fokine, A.; Battisti, A.J.; Kostyuchenko, V.A.; Black, L.W.; Rossmann, M.G. Cryo-EM structure of a bacteriophage T4 gp24 bypass mutant: The evolution of pentameric vertex proteins in icosahedral viruses. J. Struct. Biol. 2006, 154, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Showe, M.K.; Isobe, E.; Onorato, L. Bacteriophage T4 prehead proteinase: I. Purification and properties of a bacteriophage enzyme which cleaves the capsid precursor proteins. J. Mol. Biol. 1976, 107, 35–54. [Google Scholar] [CrossRef]
- Showe, M.K.; Isobe, E.; Onorato, L. Bacteriophage T4 prehead proteinase: II. Its cleavage from the product of gene 21 and regulation in phage-infected cells. J. Mol. Biol. 1976, 107, 55–69. [Google Scholar] [CrossRef]
- Black, L.W.; Rao, V.B. Structure, assembly, and DNA packaging of the bacteriophage T4 head. Adv. Virus Res. 2012, 82, 119–153. [Google Scholar]
- Müller-Salamin, L.; Onorato, L.; Showe, M.K. Localization of minor protein components of the head of bacteriophage T4. J. Virol. 1977, 24, 121–134. [Google Scholar] [CrossRef]
- Leiman, P.G.; Chipman, P.R.; Kostyuchenko, V.A.; Mesyanzhinov, V.V.; Rossmann, M.G. Three-dimensional rearrangement of proteins in the tail of bacteriophage T4 on infection of its host. Cell 2004, 118, 419–429. [Google Scholar] [CrossRef]
- Enav, H.; Kirzner, S.; Lindell, D.; Mandel-Gutfreund, Y.; Beja, O. Adaptation to sub-optimal hosts is a driver of viral diversification in the ocean. Nat. Commun. 2018, 9, 4698. [Google Scholar] [CrossRef]
- Linke, D.; Riess, T.; Autenrieth, I.B.; Lupas, A.; Kempf, V.A.J. Trimeric autotransporter adhesins: Variable structure, common function. Trends Microbiol. 2006, 14, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Eitel, J.; Dersch, P. The YadA protein of Yersinia pseudotuberculosis mediates high-efficiency uptake into human cells under environmental conditions in which invasin is repressed. Infect. Immun. 2002, 70, 4880–4891. [Google Scholar] [CrossRef] [PubMed]
- Casutt-Meyer, S.; Renzi, F.; Schmaler, M.; Jann, N.J.; Amstutz, M.; Cornelis, G.R. Oligomeric coiled-coil adhesin YadA is a double-edged sword. PLoS ONE 2010, 5, e15159. [Google Scholar] [CrossRef] [PubMed]
- Switt, A.; Orsi, R.H.; Bakker, H.; Vongkamjan, K.; Wiedmann, M. Genomic characterization provides new insight into Salmonella phage diversity. BMC Genom. 2013, 14, 481. [Google Scholar]
- Mizuno, C.; Ghai, R.; Rodriguez-Valera, F. Evidence for metaviromic islands in marine phages. Front. Microbiol. 2014, 5, 27. [Google Scholar] [CrossRef]
- Kim, S.-K.; Makino, K.; Amemura, M.; Shinagwa, H.; Nakata, A. Molecular analysis of the phoH gene, belonging to the phosphate regulon in Escherichia coli. J. Bacteriol. 1993, 175, 1316–1324. [Google Scholar] [CrossRef]
- Lillig, C.H.; Berndt, C.; Holmgren, A. Glutaredoxin systems. Biochim. Biophys. Acta 2008, 1780, 1304–1317. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Tag Number | Putative Function | Mass (kDa) | aa Length | Peptides | Coverage | Score | p-Value | emPAI |
|---|---|---|---|---|---|---|---|---|
| S-SZBM1_176 | Major capsid protein | 48.46 | 452 | 27 | 42.3% | 50,627 | 0 | 27.64 |
| S-SZBM1_180 | Portal vertex of the head | 62.08 | 535 | 19 | 32.5% | 4979 | 0 | 2.83 |
| S-SZBM1_195 | Hypothetical protein | 9.06 | 87 | 2 | 43.7% | 953 | 5.01 × 10−96 | 2.66 |
| S-SZBM1_174 | Tail completion protein | 21.59 | 191 | 5 | 22.0% | 491 | 7.94 × 10−50 | 1.38 |
| S-SZBM1_204 | Baseplate wedge | 55.49 | 511 | 13 | 31.3% | 6096 | 0 | 1.37 |
| S-SZBM1_54 | YadA domain-containing structural protein | 47.55 | 444 | 12 | 35.1% | 2493 | 5.01 × 10−250 | 1.24 |
| S-SZBM1_53 | Structural protein | 76.76 | 734 | 13 | 27.7% | 4393 | 0 | 1.21 |
| S-SZBM1_90 | DUF680 domain-containing protein | 12.58 | 122 | 2 | 25.4% | 1159 | 1.26 × 10−116 | 1.06 |
| S-SZBM1_203 | Baseplate wedge tail fiber connector | 269.22 | 2446 | 53 | 27.1% | 17,271 | 0 | 1.03 |
| S-SZBM1_206 | Baseplate wedge protein | 51.38 | 498 | 9 | 24.9% | 2195 | 3.16 × 10−220 | 0.98 |
| S-SZBM1_136 | Virion structural protein | 21.23 | 199 | 2 | 24.1% | 7040 | 0 | 0.80 |
| S-SZBM1_74 | Baseplate wedge component | 33.50 | 292 | 6 | 19.9% | 858 | 1.58 × 10−86 | 0.76 |
| S-SZBM1_182 | Tail sheath protein | 100.56 | 924 | 12 | 17.9% | 4763 | 0 | 0.72 |
| S-SZBM1_197 | Tail sheath stabilizer and completion protein | 30.07 | 265 | 4 | 22.3% | 1414 | 3.98 × 10−142 | 0.69 |
| S-SZBM1_200 | Structural protein | 215.00 | 2073 | 25 | 17.0% | 10,556 | 0 | 0.67 |
| S-SZBM1_199 | Neck protein | 31.07 | 264 | 5 | 14.0% | 541 | 7.94 × 10−55 | 0.66 |
| S-SZBM1_52 | Structural protein | 129.96 | 1192 | 16 | 20.2% | 5256 | 0 | 0.60 |
| S-SZBM1_208 | Baseplate wedge initiator | 724.20 | 6619 | 71 | 13.5% | 11,794 | 0 | 0.41 |
| S-SZBM1_139 | Hypothetical protein | 48.43 | 457 | 4 | 10.5% | 2382 | 6.31 × 10−239 | 0.39 |
| S-SZBM1_205 | Baseplate wedge initiator | 65.69 | 615 | 6 | 10.9% | 1559 | 1.26 × 10−156 | 0.34 |
| S-SZBM1_178 | Prohead core scaffolding protein and protease | 23.91 | 215 | 2 | 21.9% | 361 | 7.94 × 10−37 | 0.30 |
| S-SZBM1_140 | Cytidylyltransferase | 40.96 | 362 | 2 | 13.5% | 169 | 1.26 × 10−17 | 0.26 |
| S-SZBM1_209 | Baseplate wedge subunit | 76.50 | 691 | 5 | 9.7% | 507 | 2.00 × 10−51 | 0.23 |
| S-SZBM1_190 | Virion structural protein | 46.62 | 424 | 3 | 7.1% | 461 | 7.94 × 10−47 | 0.23 |
| S-SZBM1_177 | Prohead core protein | 37.27 | 339 | 2 | 19.8% | 248 | 1.58 × 10−25 | 0.19 |
| S-SZBM1_65 | Hypothetical protein | 60.80 | 536 | 3 | 10.8% | 183 | 5.01 × 10−19 | 0.17 |
| S-SZBM1_181 | Tail tube protein | 24.75 | 224 | 1 | 7.6% | 98 | 1.58 × 10−10 | 0.14 |
| S-SZBM1_207 | Structural protein | 58.61 | 532 | 1 | 2.6% | 245 | 3.16 × 10−25 | 0.12 |
| S-SZBM1_51 | Hypothetical protein | 30.51 | 287 | 1 | 4.9% | 120 | 1.00 × 10−12 | 0.11 |
| S-SZBM1_198 | Neck protein | 32.36 | 291 | 1 | 8.6% | 74 | 3.98 × 10−8 | 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rong, C.; Zhou, K.; Li, S.; Xiao, K.; Xu, Y.; Zhang, R.; Yang, Y.; Zhang, Y. Isolation and Characterization of a Novel Cyanophage Encoding Multiple Auxiliary Metabolic Genes. Viruses 2022, 14, 887. https://doi.org/10.3390/v14050887
Rong C, Zhou K, Li S, Xiao K, Xu Y, Zhang R, Yang Y, Zhang Y. Isolation and Characterization of a Novel Cyanophage Encoding Multiple Auxiliary Metabolic Genes. Viruses. 2022; 14(5):887. https://doi.org/10.3390/v14050887
Chicago/Turabian StyleRong, Cuhuang, Kun Zhou, Shuiming Li, Kang Xiao, Ying Xu, Rui Zhang, Yunlan Yang, and Yu Zhang. 2022. "Isolation and Characterization of a Novel Cyanophage Encoding Multiple Auxiliary Metabolic Genes" Viruses 14, no. 5: 887. https://doi.org/10.3390/v14050887
APA StyleRong, C., Zhou, K., Li, S., Xiao, K., Xu, Y., Zhang, R., Yang, Y., & Zhang, Y. (2022). Isolation and Characterization of a Novel Cyanophage Encoding Multiple Auxiliary Metabolic Genes. Viruses, 14(5), 887. https://doi.org/10.3390/v14050887