
Transcription Factor ZNF683 Inhibits SIV/HIV Replication through Regulating IFNγ Secretion of CD8+ T Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Viral Loads
2.3. Immunofluorescence Assay
2.4. Microbiome Sequencing
2.5. Histological Examination
2.6. Transcriptome Sequencing
2.7. Cytokine Gene Expression Assay
2.8. Flow Cytometry
2.9. Cells
2.10. Virus Strain and Infection In Vitro
2.11. Western Blotting Assay
2.12. Luciferase Assay
2.13. Statistical Analysis
3. Results
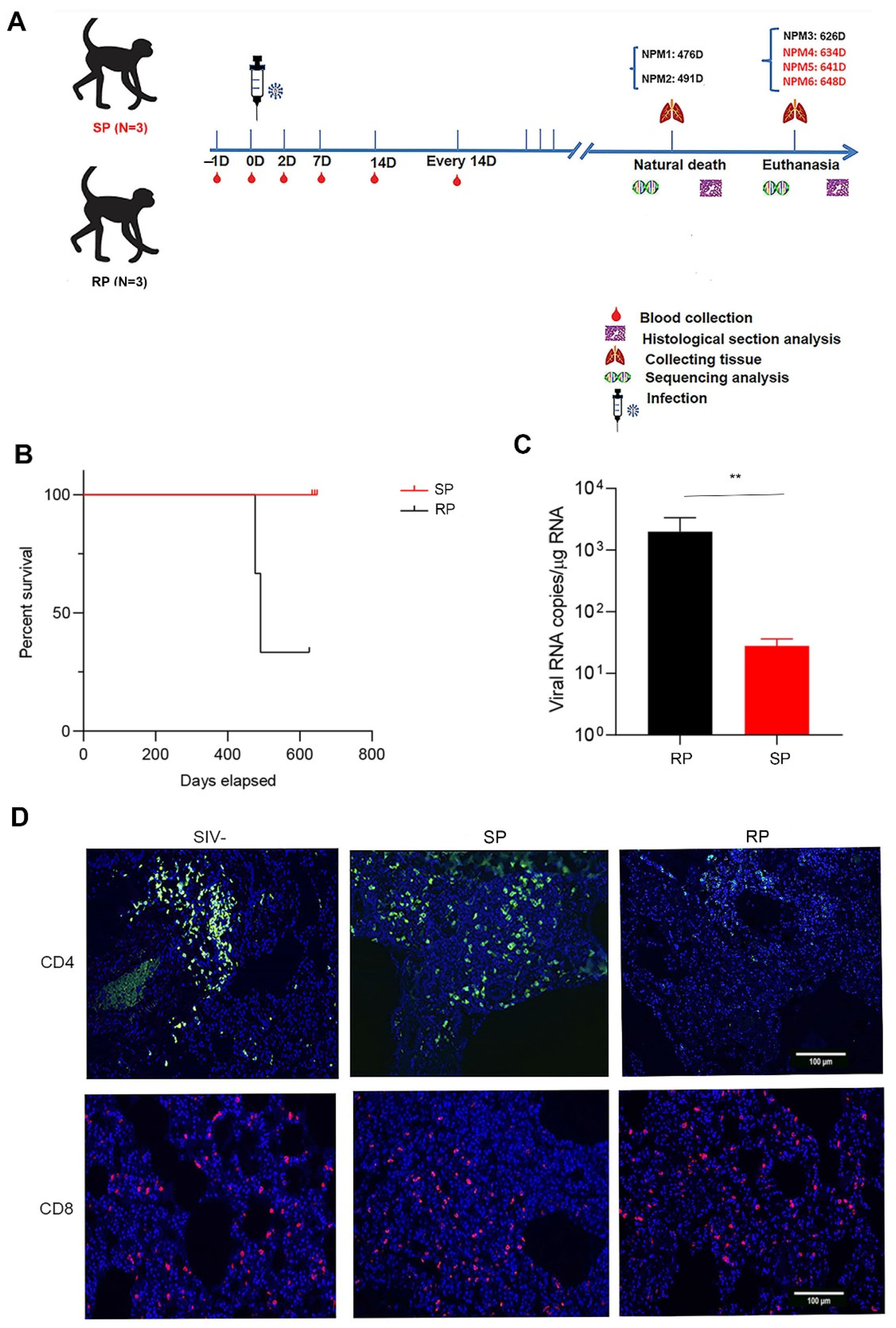
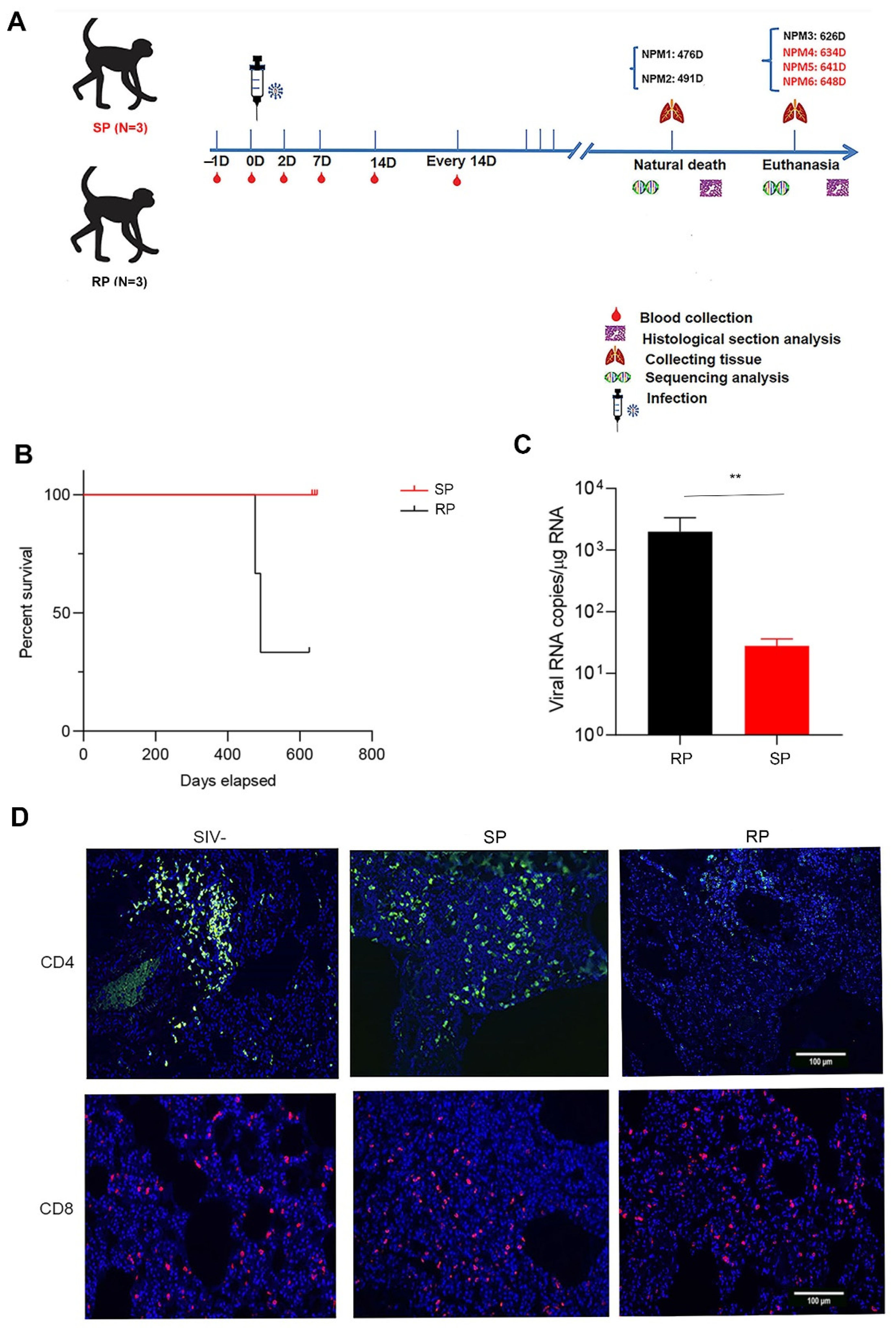
3.1. Low Levels of Viral Replication and Superior CD4+ T Cell Homeostasis in Lung Tissues Were Found in the SIVmac239-Infected SP Group
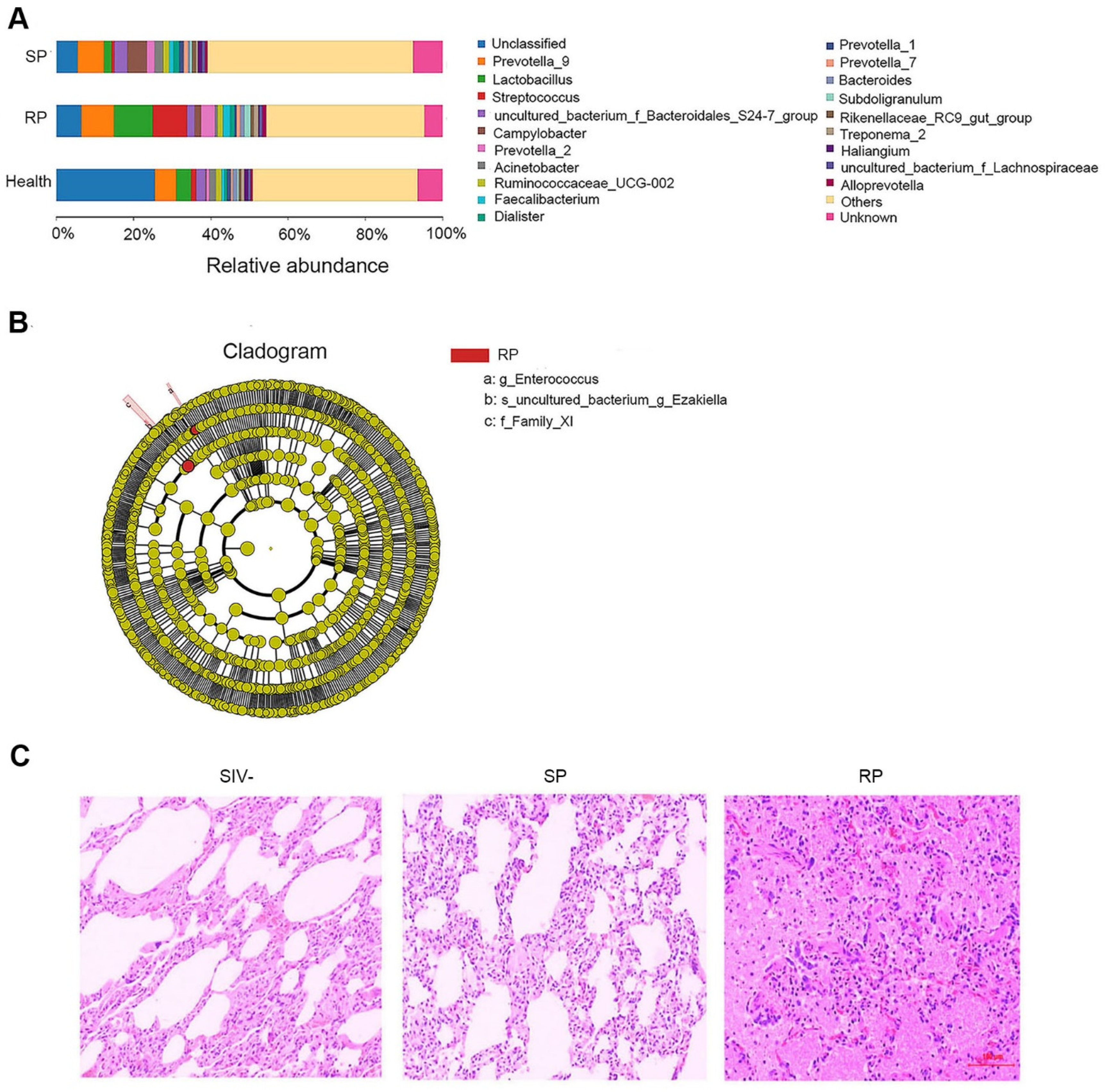
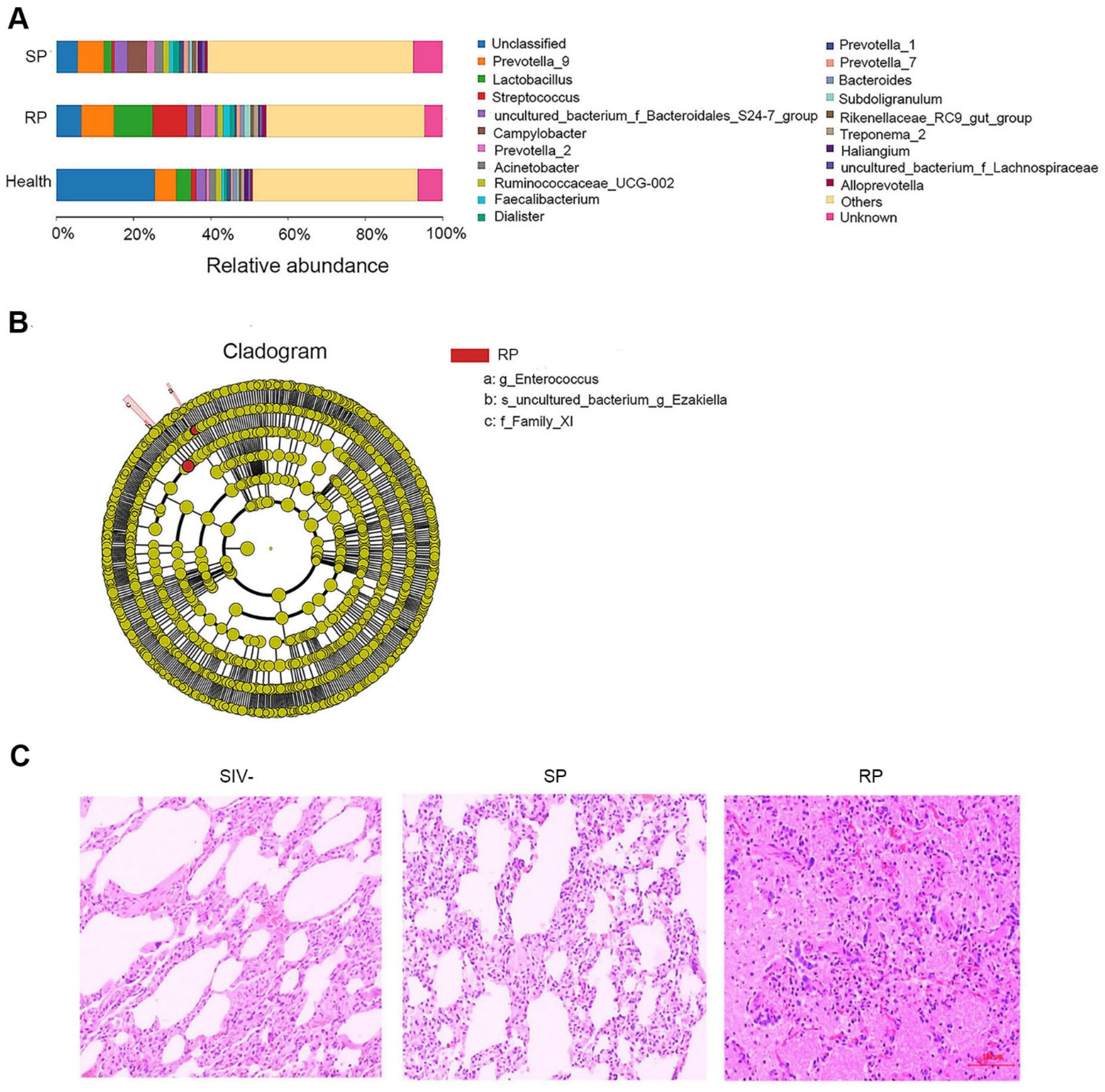
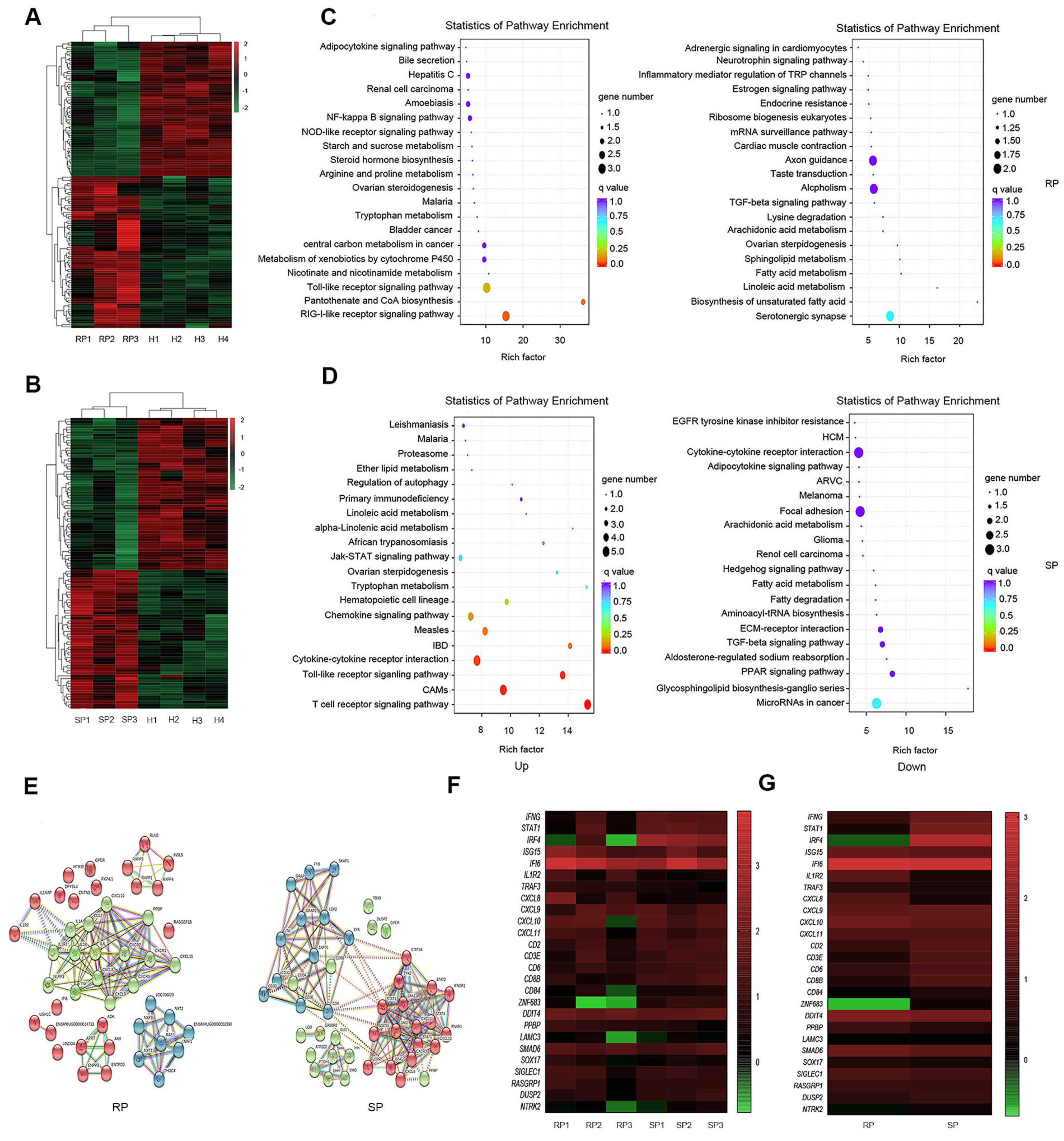
3.2. Microbial Invasion Was Significant in the Lungs of RPs
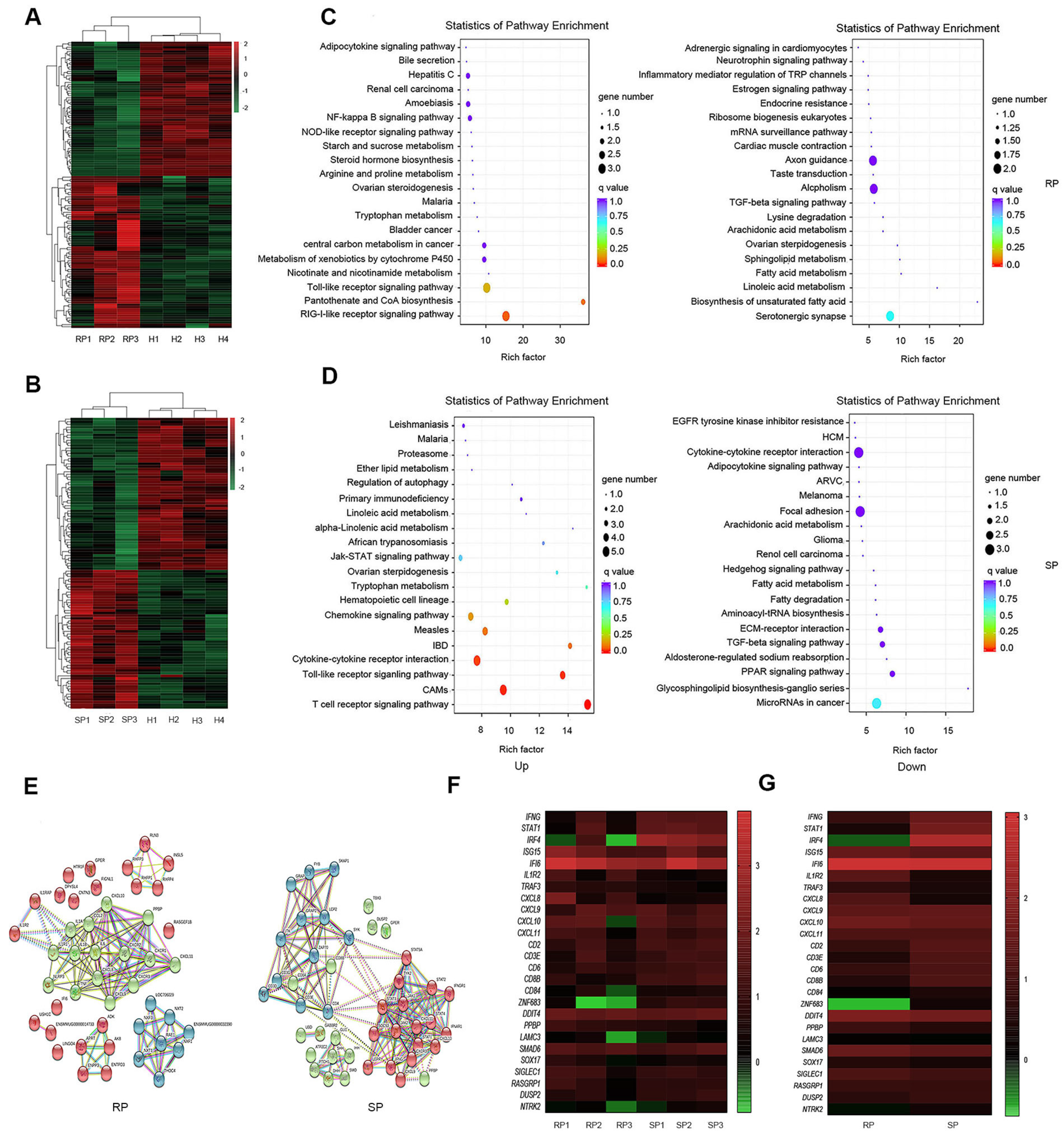
3.3. KEGG Pathway Enrichment Analysis of the DEGs
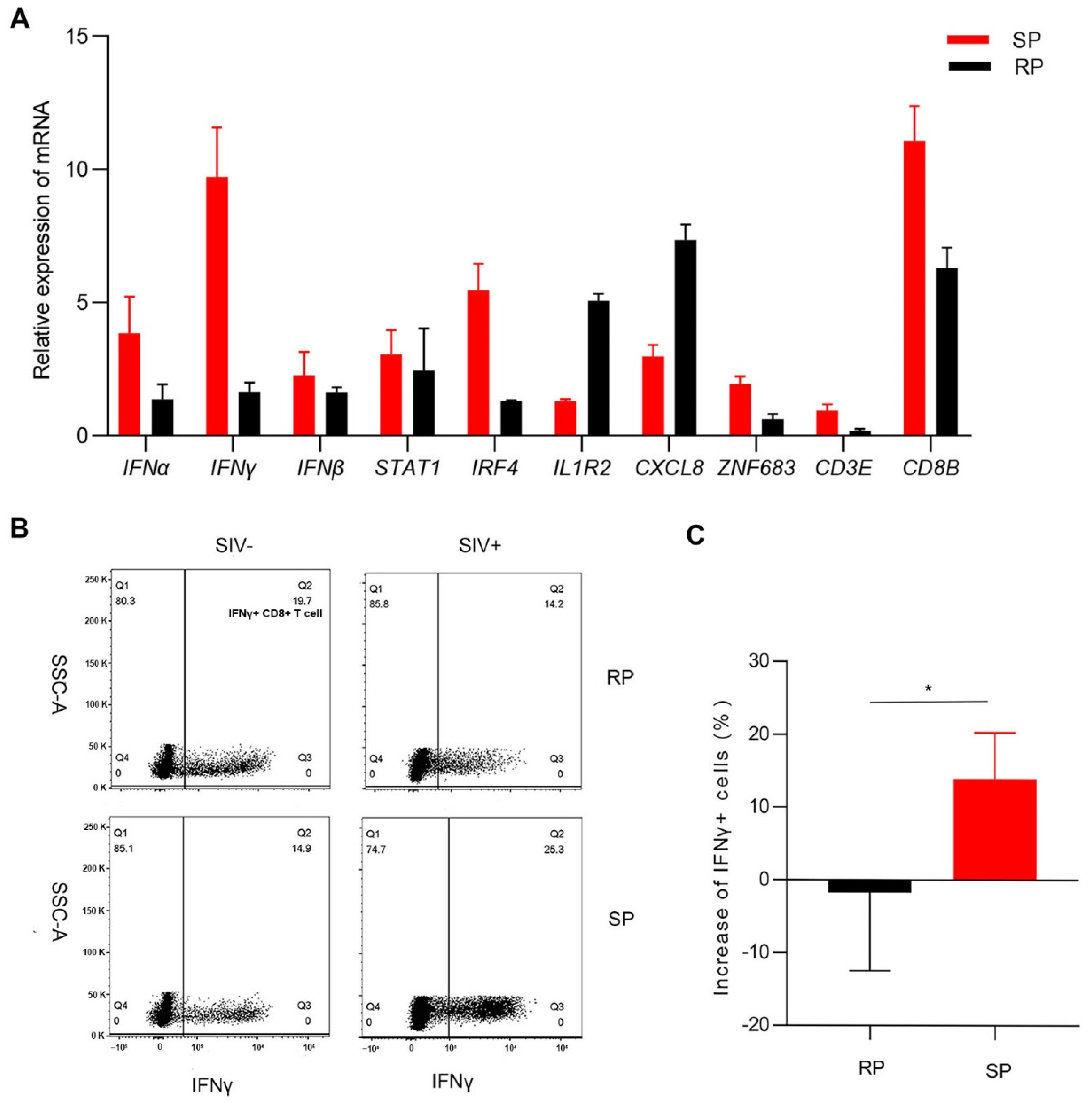
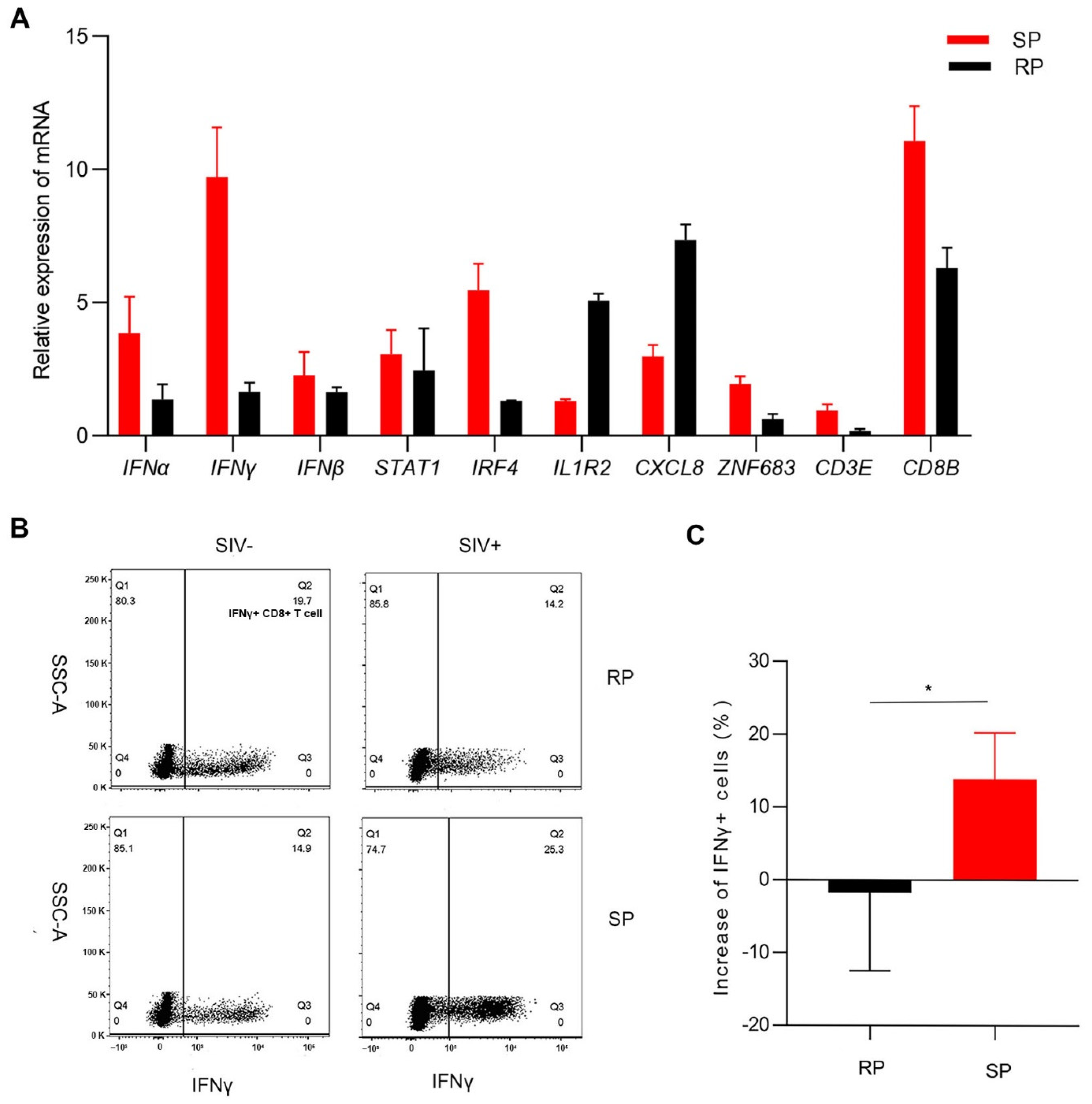
3.4. Gene Expression Confirmation by Real-Time PCR
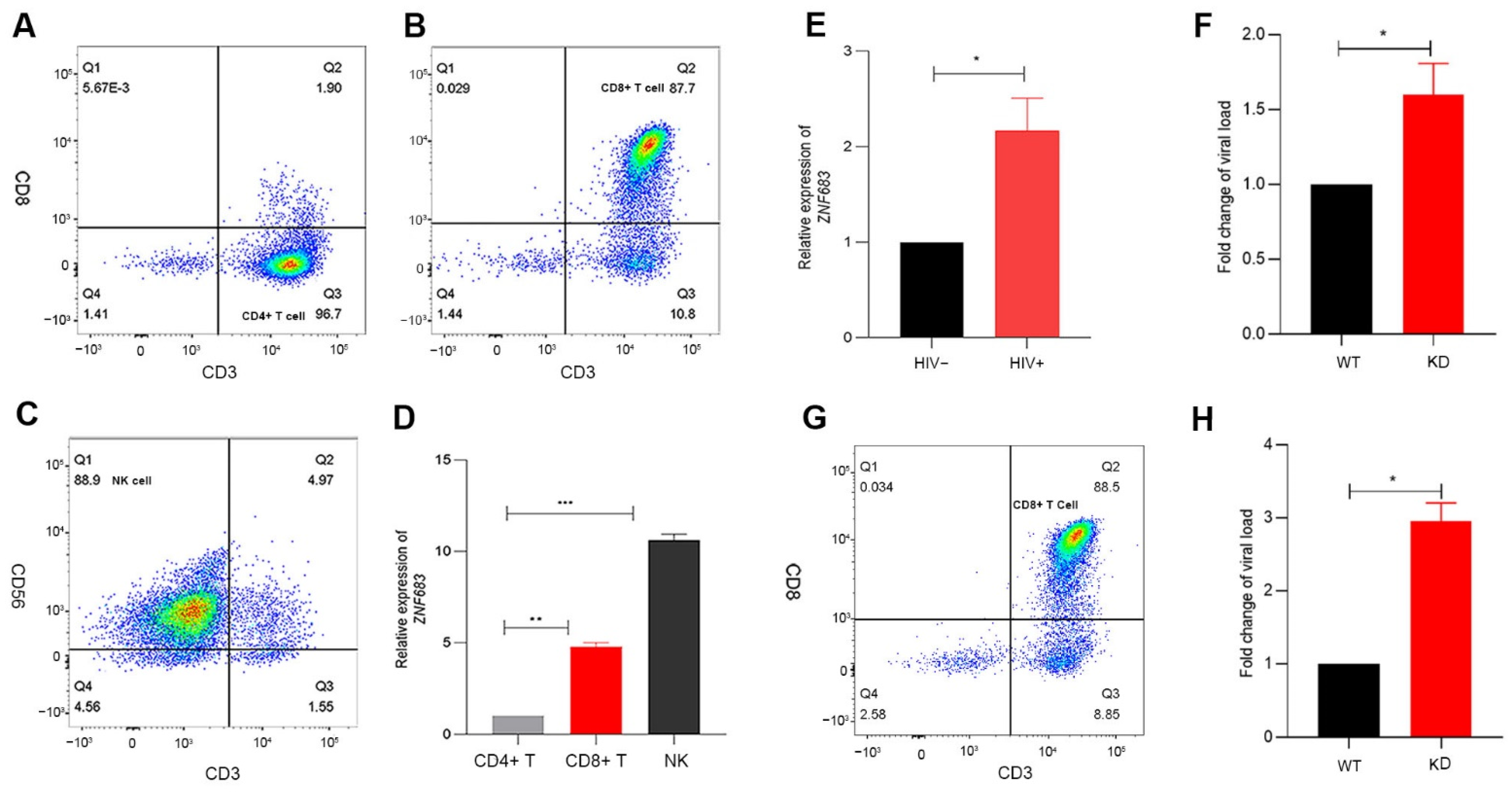
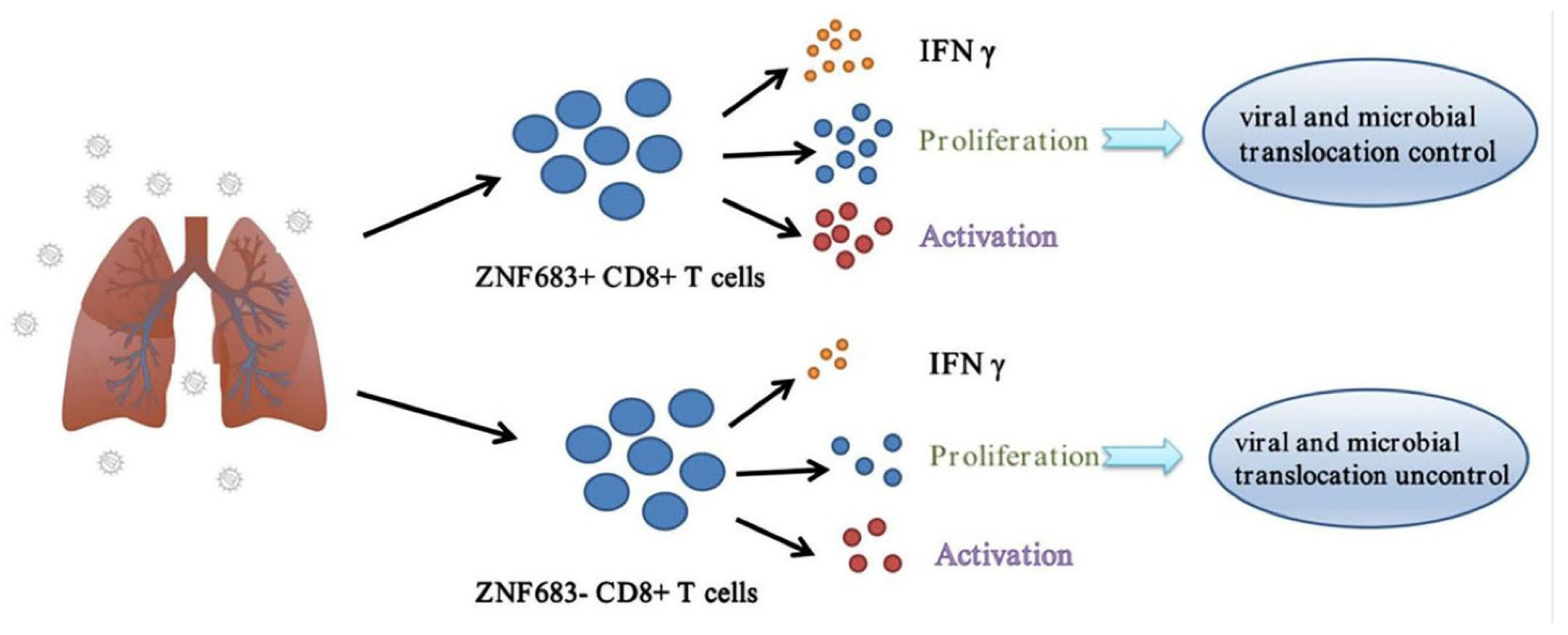
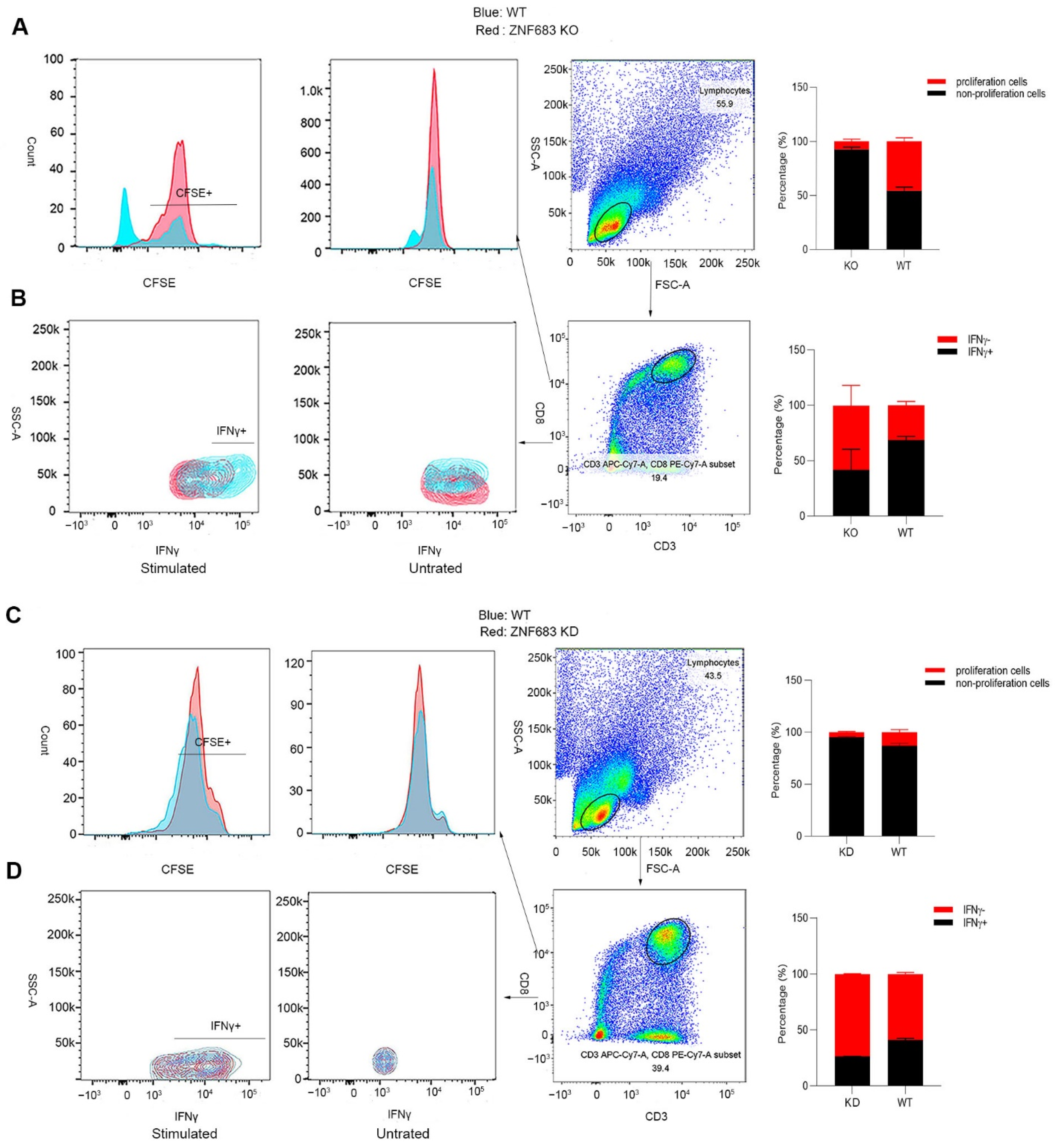
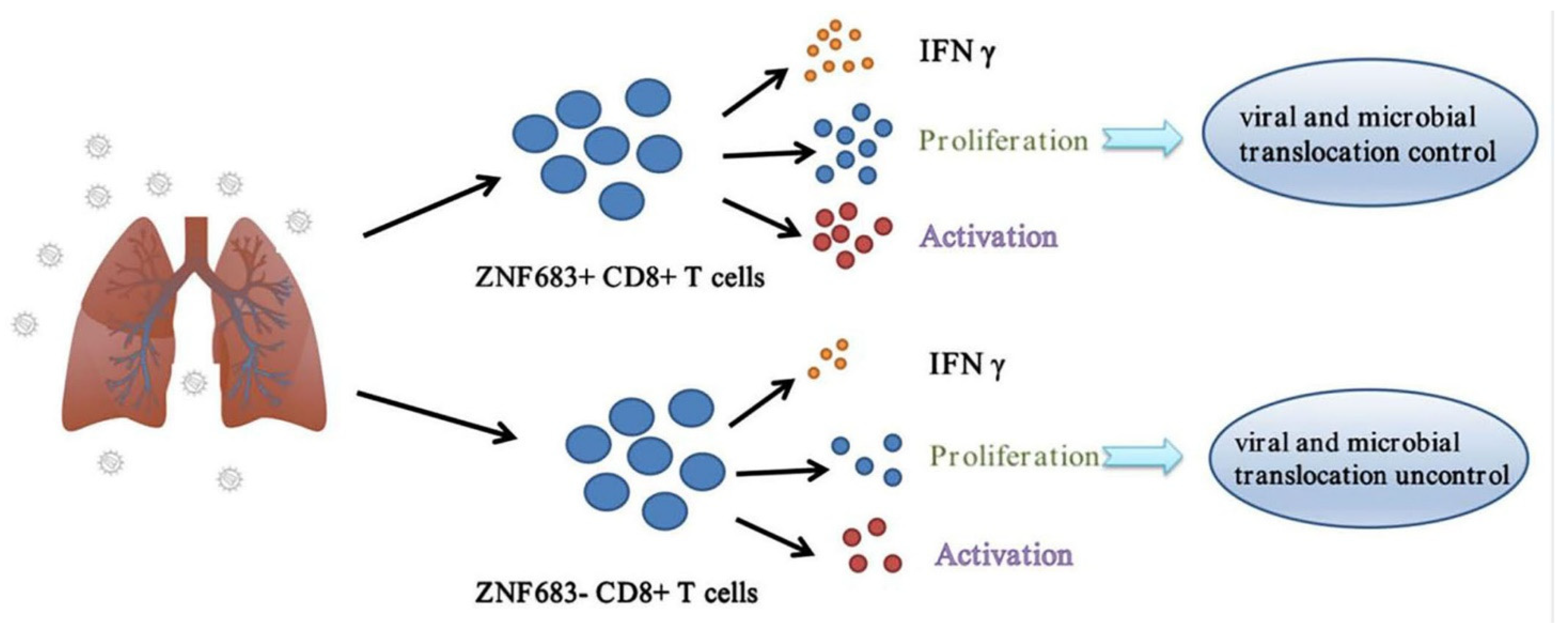
3.5. ZNF683 Regulated IFNγ Production and Proliferation of CD8+ T Cells in NPMs and Humans
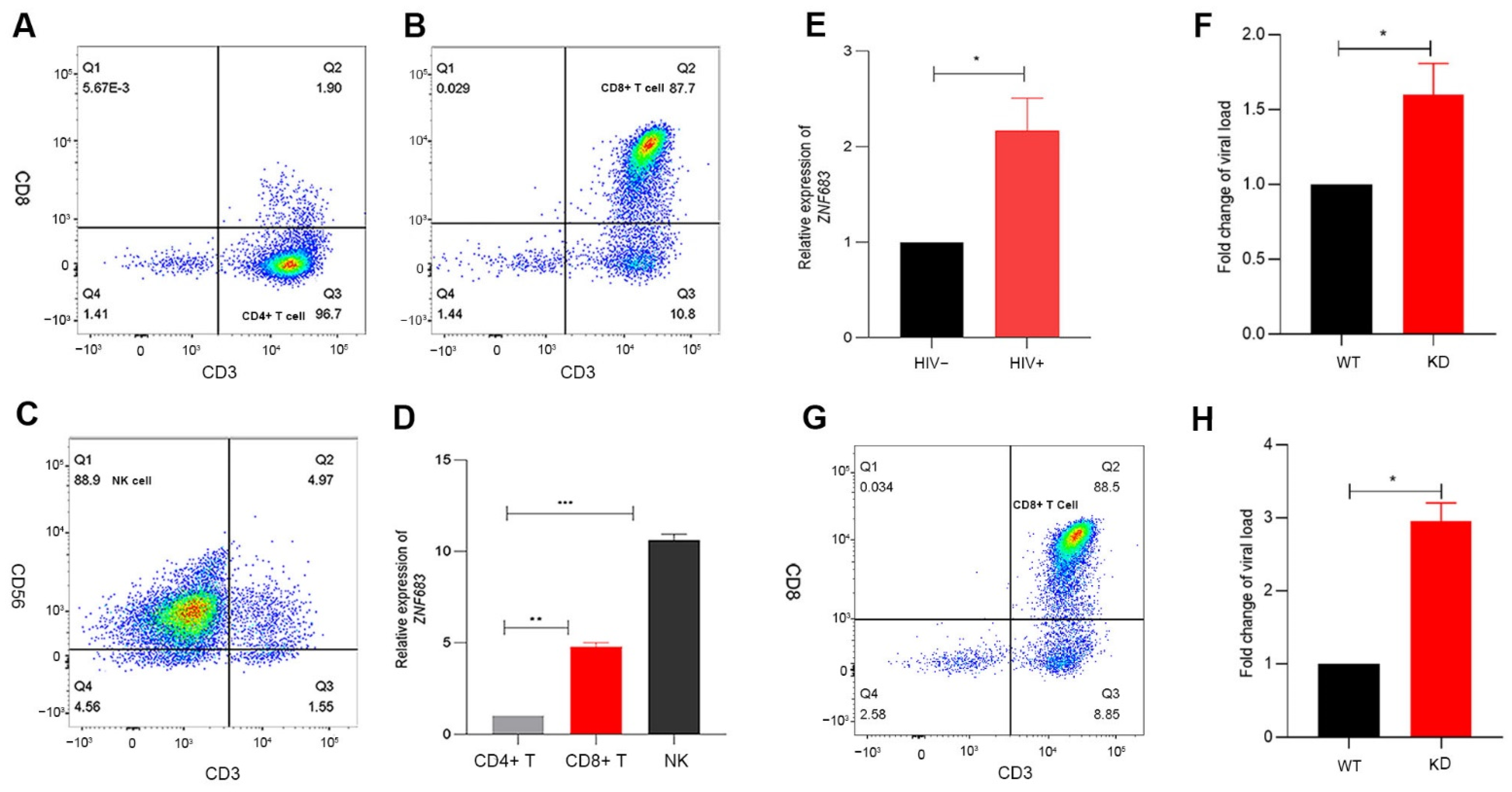
3.6. ZNF683 Was Upregulated in HIV Patients and Inhibited HIV Infection In Vitro
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suphanchaimat, R.; Sommanustweechai, A.; Khitdee, C.; Thaichinda, C.; Kantamaturapoj, K.; Leelahavarong, P.; Jumriangrit, P.; Topothai, T.; Wisaijohn, T.; Putthasri, W. HIV/AIDS health care challenges for cross-country migrants in low-and middle-income countries: A scoping review. HIV/AIDS (Auckl.) 2014, 6, 19–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.M.; Maldarelli, F. The role of integration and clonal expansion in HIV infection: Live long and prosper. Retrovirology 2018, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Mehraj, V.; Kaufmann, D.E.; Li, T.; Routy, J.P. Elevation and persistence of CD8 T-cells in HIV infection: The Achilles heel in the ART era. J. Int. AIDS Soc. 2016, 19, 20697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalo-Gil, E.; Ikediobi, U.; Sutton, R.E. Focus: Infectious diseases: Mechanisms of virologic control and clinical characteristics of HIV+ elite/viremic controllers. Yale J. Biol. Med. 2017, 90, 245–259. [Google Scholar] [PubMed]
- Nishimura, Y.; Martin, M.A. Of mice, macaques, and men: Broadly neutralizing antibody immunotherapy for HIV-1. Cell Host Microbe 2017, 22, 207–216. [Google Scholar] [CrossRef]
- González, N.; McKee, K.; Lynch, R.M.; Georgiev, I.S.; Jimenez, L.; Grau, E.; Yuste, E.; Kwong, P.D.; Mascola, J.R.; Alcamí, J. Characterization of broadly neutralizing antibody responses to HIV-1 in a cohort of long term non-progressors. PLoS ONE 2018, 13, e0193773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringel, O.; Müller, K.; Koch, J.; Brill, B.; Wolf, T.; Stephan, C.; Vieillard, V.; Debré, P.; Dietrich, U. Optimization of the EC26-2A4 epitope in the gp41 membrane proximal external region targeted by neutralizing antibodies from an elite controller. AIDS Res. Hum. Retrovir. 2018, 34, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Buckheit, R.W.; Siliciano, R.F.; Blankson, J.N. Primary CD8+ T cells from elite suppressors effectively eliminate non-productively HIV-1 infected resting and activated CD4+ T cells. Retrovirology 2013, 10, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Sabbaj, S.; Bansal, A.; Amatya, N.; Shacka, J.J.; Goepfert, P.A.; Heath, S.L. HIV-specific CD8+ T cells from elite controllers are primed for survival. J. Virol. 2013, 87, 5170–5181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korolevskaya, L.B.; Shmagel, K.V.; Shmagel, N.G.; Saidakova, E.V. Systemic activation of the immune system in HIV infection: The role of the immune complexes (hypothesis). Med. Hypotheses 2016, 88, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Segal, L.N.; Methé, B.A.; Nolan, A.; Hoshino, Y.; Rom, W.N.; Dawson, R.; Bateman, E.; Weiden, M.D. HIV-1 and bacterial pneumonia in the era of antiretroviral therapy. Proc. Am. Thorac. Soc. 2011, 8, 282–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picker, L.J. Immunopathogenesis of acute AIDS virus infection. Curr. Opin. Immunol. 2006, 18, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.M.; Salters, K.A.; Eyawo, O.; Franco-Villalobos, C.; Jabbari, S.; Wiseman, S.M.; Press, N.; Montaner, J.S.; Man, S.P.; Hull, M. Mortality among people living with HIV/AIDS with non-small-cell lung cancer in the modern HAART Era. AIDS Care 2018, 30, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Skalski, J.H.; Limper, A.H. Fungal, viral, and parasitic pneumonias associated with human immunodeficiency virus. Semin. Respir. Crit. Care Med. 2016, 37, 257–266. [Google Scholar] [CrossRef]
- Moniuszko, M.; Bogdan, D.; Pal, R.; Venzon, D.; Stevceva, L.; Nacsa, J.; Tryniszewska, E.; Edghill-Smith, Y.; Wolinsky, S.; Franchini, G. Correlation between viral RNA levels but not immune responses in plasma and tissues of macaques with long-standing SIVmac251 infection. Virology 2005, 333, 159–168. [Google Scholar] [CrossRef]
- Qin, S.; Junecko, B.A.F.; Trichel, A.M.; Tarwater, P.M.; Murphey-Corb, M.A.; Kirschner, D.E.; Reinhart, T.A. Simian immunodeficiency virus infection alters chemokine networks in lung tissues of cynomolgus macaques: Association with Pneumocystis carinii infection. Am. J. Pathol. 2010, 177, 1274–1285. [Google Scholar] [CrossRef]
- Hessell, A.J.; Haigwood, N.L. Animal models in HIV-1 protection and therapy. Curr. Opin. HIV AIDS 2015, 10, 170–176. [Google Scholar] [CrossRef]
- Mallard, J.; Williams, K.C. Animal models of HIV-associated disease of the central nervous system. Handb. Clin. Neurol. 2018, 152, 41–53. [Google Scholar]
- Veazey, R.S.; Lackner, A.A. Nonhuman primate models and understanding the pathogenesis of HIV infection and AIDS. ILAR J. 2017, 58, 160–171. [Google Scholar] [CrossRef]
- Gardner, M. SIV vaccines: Current status: The role of the SIV—Macaque model in AIDS research. Vaccine 1991, 9, 787–791. [Google Scholar] [CrossRef]
- Antony, J.M.; MacDonald, K.S. A critical analysis of the cynomolgus macaque, Macaca fascicularis, as a model to test HIV-1/SIV vaccine efficacy. Vaccine 2015, 33, 3073–3083. [Google Scholar] [CrossRef]
- Zhang, M.X.; Zheng, H.Y.; Jiang, J.; Song, J.H.; Chen, M.; Xiao, Y.; Lian, X.D.; Song, T.Z.; Tian, R.R.; Pang, W. Predict disease progression from T-cell phenotypes in northern pig-tailed macaques (Macaca leonina) during SIVmac239 infection. Immunology 2017, 152, 574–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.X.; Zheng, H.Y.; Jiang, J.; Song, J.H.; Chen, M.; Xiao, Y.; Lian, X.D.; Song, T.Z.; Tian, R.R.; Pang, W. Northern pig-tailed macaques (Macaca leonina) maintain superior CD4+ T-cell homeostasis during SIVmac239 infection. Eur. J. Immunol. 2018, 48, 384–385. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Song, J.-H.; Lu, Y.; Zhang, X.-L.; Zheng, H.-Y.; Jiang, J.; Zheng, Y.-T. Host restriction factors APOBEC3G/3F and other interferon-related gene expressions affect early HIV-1 infection in northern pig-tailed macaque (Macaca leonina). Front. Immunol. 2018, 9, 1965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tian, R.; Zheng, H.; Pan, G.; Tuo, X.; Xia, H.; Xia, X.; Pang, W.; Zheng, Y. Translocation of microbes and changes of immunocytes in the gut of rapid-and slow-progressor Chinese rhesus macaques infected with SIV mac239. Immunology 2016, 147, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, A.T.; Wolfe, D. Tissue processing and hematoxylin and eosin staining. Methods Mol. Biol. 2014, 1180, 31–43. [Google Scholar]
- Ding, Z.; Ma, M.; Tao, L.; Peng, Y.; Han, Y.; Sun, L.; Dai, X.; Ji, Z.; Bai, R.; Jian, M. Rhesus brain transcriptomic landscape in an ex vivo model of the interaction of live Borrelia burgdorferi with frontal cortex tissue explants. Front. Neurosci. 2019, 13, 651. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Zhang, M.; Pang, W.; Zheng, Y. Aged Chinese rhesus macaques suffer severe phenotypic T- and B-cell aging accompanied with sex differences. Exp. Gerontol. 2014, 55, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Vieira Braga, F.A.; Hertoghs, K.M.; Kragten, N.A.; Doody, G.M.; Barnes, N.A.; Remmerswaal, E.B.; Hsiao, C.C.; Moerland, P.D.; Wouters, D.; Derks, I.A. Blimp-1 homolog Hobit identifies effector-type lymphocytes in humans. Eur. J. Immunol. 2015, 45, 2945–2958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kragten, N.A.; Behr, F.M.; Vieira Braga, F.A.; Remmerswaal, E.B.; Wesselink, T.H.; Oja, A.E.; Hombrink, P.; Kallies, A.; van Lier, R.A.; Stark, R. Blimp-1 induces and Hobit maintains the cytotoxic mediator granzyme B in CD8 T cells. Eur. J. Immunol. 2018, 48, 1644–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, G.; Tincati, C.; Silvestri, G. Microbial translocation in the pathogenesis of HIV infection and AIDS. Clin. Microbiol. Rev. 2013, 26, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinh, D.M.; Volpe, G.E.; Duffalo, C.; Bhalchandra, S.; Tai, A.K.; Kane, A.V.; Wanke, C.A. Intestinal microbiota, microbial translocation, and systemic inflammation in chronic HIV infection. J. Infect. Dis. 2015, 211, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Watabe, T.; Kato, K.; Tsukimi, T.; Sato, M.P.; Odamaki, T.; Tomita, M.; Fukuda, S. Draft genome sequences of enterococcus faecalis strains isolated from healthy Japanese individuals. Microbiol. Resour. Announc. 2019, 8, e00832-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.Y.; Ahn, Y.; Khare, S.; Gokulan, K.; Piñeiro, S.A.; Cerniglia, C.E. An in vitro study to assess the impact of tetracycline on the human intestinal microbiome. Anaerobe 2018, 49, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Diop, A.; Diop, K.; Tomei, E.; Raoult, D.; Fenollar, F.; Fournier, P.-E. Draft genome sequence of Ezakiella peruensis Strain M6.X2, a human gut gram-positive anaerobic coccus. Genome Announc. 2018, 6, e01487-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenchley, J.M.; Proce, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lamnotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, G.; Cozzi-Lepri, A.; Merlini, E.; Bellistrì, G.M.; Castagna, A.; Galli, M.; Verucchi, G.; Antinori, A.; Costantini, A.; Giacometti, A.; et al. Microbial translocation predicts disease progression of HIV-infected antiretroviral-naïve patients with high CD4+ cell count. Aids 2011, 25, 1385–1394. [Google Scholar] [CrossRef]
- Sandler, N.G.; Wand, H.; Roque, A.; Law, M.; Nason, M.C.; Nixon, D.E.; Pedersen, C.; Ruxrungtham, K.; Lewin, S.R.; Emery, D.; et al. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J. Infect. Dis. 2011, 203, 780–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipp, H.; Zemlin, A.E.; Erasmus, R.T.; Glashoff, R.H. Role of inflammation in HIV-1 disease progression and prognosis. Crit. Rev. Clin. Lab. Sci. 2014, 51, 98–111. [Google Scholar] [CrossRef]
- Tian, R.; Zhang, M.; Zhang, L.; Zhang, X.; Zheng, H.; Zhu, L.; Pang, W.; Zhang, G.; Zheng, Y. High immune activation and abnormal expression of cytokines contribute to death of SHIV89. 6-infected Chinese rhesus macaques. Arch. Virol. 2015, 160, 1953–1966. [Google Scholar] [CrossRef] [PubMed]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Wilson, S.J.; Panis, M.; Murphy, M.Y.; Jones, C.T.; Bieniasz, P.; Rice, C.M. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature 2011, 472, 481–485. [Google Scholar] [CrossRef]
- Lee, A.; Ashkar, A. The dual nature of type I and type II interferons. Front. Immunol. 2018, 9, 2061. [Google Scholar]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [Green Version]
- Muema, D.M.; Mthembu, M.; Schiff, A.E.; Singh, U.; Corleis, B.; Chen, D.; Bassett, T.; Rasehlo, S.S.; Nyamande, K.; Khan, D.F. Contrasting inflammatory signatures in peripheral blood and bronchoalveolar cells reveal compartment-specific effects of HIV infection. Front. Immunol. 2020, 11, 864. [Google Scholar] [CrossRef]
- Mackay, L.K.; Minnich, M.; Kragten, N.A.; Liao, Y.; Nota, B.; Seillet, C.; Zaid, A.; Man, K.; Preston, S.; Freestone, D. Hobit and Blimp1 instruct a universal transcriptional program of tissue residency in lymphocytes. Science 2016, 352, 459–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, J.; Frentsch, M.; Thiel, A. Hobit and human effector T-cell differentiation: The beginning of a long journey. Eur. J. Immunol. 2015, 45, 2762–2765. [Google Scholar] [CrossRef] [PubMed]
- Gisbergen, K.V.; Kragten, N.; Hertoghs, K.; Wensveen, F.M.; Jonjic, S.; Hamann, J.; Nolte, M.A.; Liver, R.V. Mouse hobit is a homolog of the transcriptional repressor blimp-1 that regulates nkt cell effector differentiation. Nat Immunol. 2012, 13, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Doody, G.M.; Care, M.A.; Burgoyne, N.J.; Bradford, J.R.; Bota, M.; Bonifer, C.; Westhead, D.R.; Tooze, R.M. An extended set of PRDM1/BLIMP1 target genes links binding motif type to dynamic repression. Nucleic Acids Res. 2010, 38, 5336–5350. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, T.H.; Appell, K.C.; Berg, L. Janus-kinase-3-dependent signals induce chromatin remodeling at the Ifng locus during T helper 1 cell differentiation. Immunity 2008, 28, 763–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | 5′→3′ |
|---|---|
| IFNγ-F | CTGTTACTGCCAGGACCCAT |
| IFNγ-R | TGCTACATCTGGATCACCTGC |
| IFNα-F | GCCTGAAGGACAGACATGACTTT |
| IFNα-R | GGATGGTTTGAGCCTTTTGG |
| GAPDH-F | GCT TGAGGGTTTGCTACAACATG |
| GAPDH-R | GACGCCTGCTTCACTACCTT |
| IFNβ-F | TGCCTCAAGGACAGGATGAAC |
| IFNβ-R | GCGTCCTCCTTCTGGAACTG |
| CD3E-F | GCCGCTTCTTCCTTTGAAGC |
| CD3E-R | ATCCAAGGGGGAGGGAATGA |
| CD8B-F | GCGTTCTGGTTTTGCTGGTT |
| CD8B-R | TGTAGTTTCCGTGCAGGCAT |
| ZNF683-F | CATATGTGGCAAGAGCTTTGG |
| ZNF683-R | AGAGCTTCACTCAACTTGCC-3 |
| STAT1-F | TCTTCTGCCGGGTAGTTTCG |
| STAT1-R | CTCGAGGATGGCATACAGCA |
| IRF4-F | TGTGAAAATGGTTGCCAGGTG |
| IRF4-R | TCACGAGGATTTCCCGGTAG |
| IL1R2-F | TGCTCTTAAAAACTAGCCACGCA |
| IL1R2-R | CGTGGCAGAACCTGCTTTGA |
| CXCL8-F | ACTCCAAACCTTTCCACCCC |
| CXCL8-R | TTCCTTGGGGTCCAGACAGA |
| 18S-F | GGACAACAAGCTCCGTGAAGA |
| 18S-R | CAGAAGTGACGCAGCCCTCTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Zhang, M.-X.; Pang, W.; Song, T.-Z.; Zheng, H.-Y.; Tian, R.-R.; Zheng, Y.-T. Transcription Factor ZNF683 Inhibits SIV/HIV Replication through Regulating IFNγ Secretion of CD8+ T Cells. Viruses 2022, 14, 719. https://doi.org/10.3390/v14040719
Lu Y, Zhang M-X, Pang W, Song T-Z, Zheng H-Y, Tian R-R, Zheng Y-T. Transcription Factor ZNF683 Inhibits SIV/HIV Replication through Regulating IFNγ Secretion of CD8+ T Cells. Viruses. 2022; 14(4):719. https://doi.org/10.3390/v14040719
Chicago/Turabian StyleLu, Ying, Ming-Xu Zhang, Wei Pang, Tian-Zhang Song, Hong-Yi Zheng, Ren-Rong Tian, and Yong-Tang Zheng. 2022. "Transcription Factor ZNF683 Inhibits SIV/HIV Replication through Regulating IFNγ Secretion of CD8+ T Cells" Viruses 14, no. 4: 719. https://doi.org/10.3390/v14040719
APA StyleLu, Y., Zhang, M.-X., Pang, W., Song, T.-Z., Zheng, H.-Y., Tian, R.-R., & Zheng, Y.-T. (2022). Transcription Factor ZNF683 Inhibits SIV/HIV Replication through Regulating IFNγ Secretion of CD8+ T Cells. Viruses, 14(4), 719. https://doi.org/10.3390/v14040719