
Cytolytic Properties and Genome Analysis of Rigvir® Oncolytic Virotherapy Virus and Other Echovirus 7 Isolates



Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses and Virus Culture
2.2. Cell Lines Used to Test Infectivity and Viability
2.3. Virus Infectivity and Viability Assays
2.4. Immunofluorescence Microscopy
2.5. Isolation of Viral RNA and Determination of Cycle Threshold Values
2.6. Genome Sequences and Sequence Analysis
2.7. Structural Analysis of E7–DAF Interface and Rigvir® Mutations
3. Results
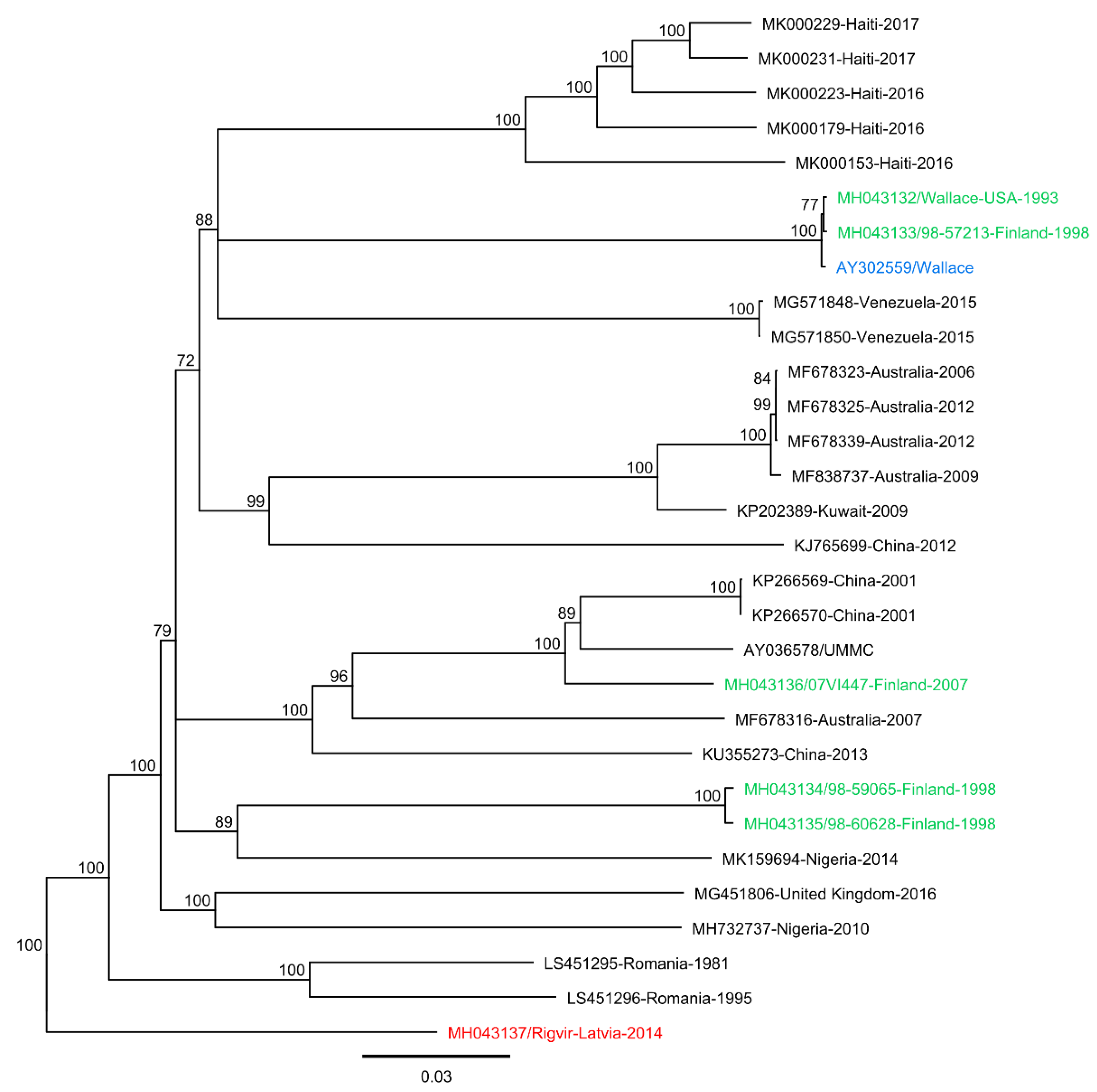
3.1. Sequence Analyses and Genome Comparisons
3.2. Structural Analysis of DAF Binding Site
3.3. Cell Infectivity and Viability Assays and Immunofluorescence Microscopy
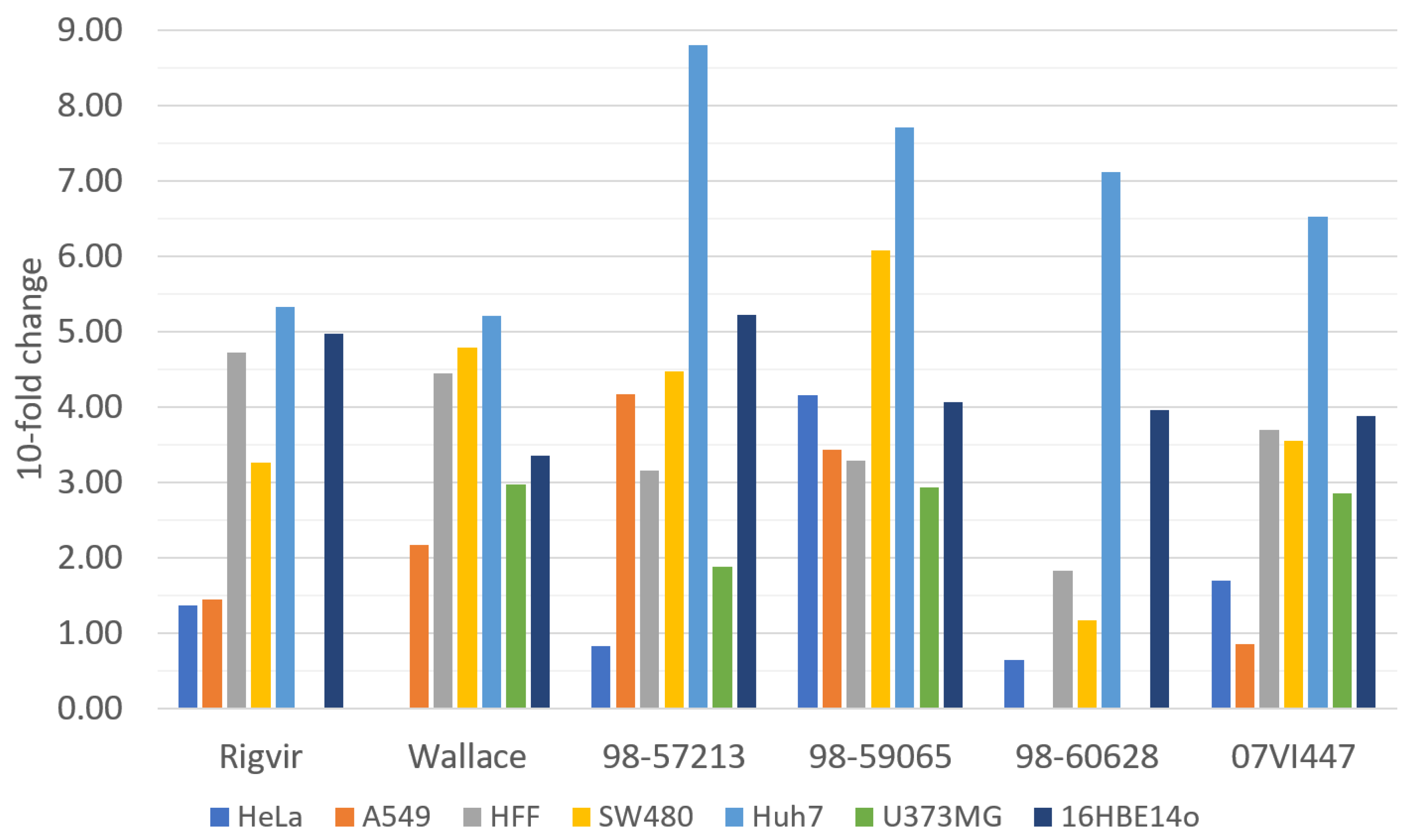
3.4. Determination of Relative Infectivities by RT-qPCR
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelly, E.; Russell, S.J. History of Oncolytic Viruses: Genesis to Genetic Engineering. Mol. Ther. 2007, 15, 651–659. [Google Scholar] [CrossRef]
- Liu, T.-C.; Galanis, E.; Kirn, D. Clinical trial results with oncolytic virotherapy: A century of promise, a decade of progress. Nat. Clin. Pract. Oncol. 2007, 4, 101–117. [Google Scholar] [CrossRef]
- Hu, J.C.C.; Coffin, R.S.; Davis, C.J.; Graham, N.J.; Groves, N.; Guest, P.J.; Harrington, K.J.; James, N.D.; Love, C.A.; McNeish, I.; et al. A Phase I Study of OncoVEX GM-CSF, a Second-Generation Oncolytic Herpes Simplex Virus Expressing Granulocyte Macrophage Colony-Stimulating Factor. Clin. Cancer Res. 2006, 12, 6737–6747. [Google Scholar] [CrossRef]
- Conry, R.M.; Westbrook, B.; McKee, S.; Norwood, T.G. Talimogene laherparepvec: First in class oncolytic virotherapy. Hum. Vaccin. Immunother. 2018, 14, 839–846. [Google Scholar] [CrossRef]
- Ylä-Pelto, J.; Tripathi, L.; Susi, P. Therapeutic Use of Native and Recombinant Enteroviruses. Viruses 2016, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, C.; Jayawardena, N.; Burga, L.N.; Bostina, M. Developing Picornaviruses for Cancer Therapy. Cancers 2019, 11, 685. [Google Scholar] [CrossRef] [PubMed]
- Shafren, D.R.; Au, G.G.; Nguyen, T.; Newcombe, N.G.; Haley, E.S.; Beagley, L.; Johansson, E.S.; Hersey, P.; Barry, R.D. Systemic Therapy of Malignant Human Melanoma Tumors by a Common Cold-Producing Enterovirus, Coxsackievirus A21. Clin. Cancer Res. 2004, 10, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Au, G.G.; Lincz, L.F.; Enno, A.; Shafren, D.R. Oncolytic Coxsackievirus A21 as a novel therapy for multiple myeloma. Br. J. Haematol. 2007, 137, 133–141. [Google Scholar] [CrossRef]
- Berry, L.J.; Au, G.G.; Barry, R.D.; Shafren, D.R. Potent Oncolytic activity of human enteroviruses against human prostate cancer. Prostate 2008, 68, 577–587. [Google Scholar] [CrossRef]
- Oberste, M.S.; Peñaranda, S.; Maher, K.; Pallansch, M.A. Complete genome sequences of all members of the species Human enterovirus A. J. Gen. Virol. 2004, 85, 1597–1607. [Google Scholar] [CrossRef]
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus Taxonomy Profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Jacques, J.; Moret, H.; Minette, D.; Lévêque, N.; Jovenin, N.; Deslée, G.; Lebargy, F.; Motte, J.; Andréoletti, L. Epidemiological, Molecular, and Clinical Features of Enterovirus Respiratory Infections in French Children between 1999 and 2005. J. Clin. Microbiol. 2008, 46, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, S.; Paananen, A.; Savolainen-Kopra, C.; Hovi, T.; Roivainen, M. Eight Years of Experience with Molecular Identification of Human Enteroviruses. J. Clin. Microbiol. 2008, 46, 2410–2413. [Google Scholar] [CrossRef] [PubMed]
- Benschop, K.S.M.; van der Avoort, H.G.; Jusic, E.; Vennema, H.; van Binnendijk, R.; Duizer, E. Polio and Measles Down the Drain: Environmental Enterovirus Surveillance in the Netherlands, 2005 to 2015. Appl. Environ. Microbiol. 2017, 83, e00558-17. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Bergelson, J.M. Echovirus 7 entry into polarized intestinal epithelial cells requires clathrin and Rab7. MBio 2012, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tuthill, T.J.; Groppelli, E.; Hogle, J.M.; Rowlands, D.J. Picornaviruses. In Cell Entry by Non-Enveloped Viruses; Springer: Berlin/Heidelberg, Germany, 2010; pp. 43–89. [Google Scholar]
- Alberts, P.; Tilgase, A.; Rasa, A.; Bandere, K.; Venskus, D. The advent of oncolytic virotherapy in oncology: The Rigvir® story. Eur. J. Pharmacol. 2018, 837, 117–126. [Google Scholar] [CrossRef]
- Rigvir Therapeutic Characteristics. Available online: https://www.rigvir.com/products/rigvir.php (accessed on 19 February 2022).
- Alberts, P.; Olmane, E.; Brokāne, L.; Krastiņa, Z.; Romanovska, M.; Kupčs, K.; Isajevs, S.; Proboka, G.; Erdmanis, R.; Nazarovs, J.; et al. Long-term treatment with the oncolytic ECHO-7 virus Rigvir of a melanoma stage IV M1c patient, a small cell lung cancer stage IIIA patient, and a histiocytic sarcoma stage IV patient-three case reports. APMIS 2016, 124, 896–904. [Google Scholar] [CrossRef]
- Doniną, S.; Strele, I.; Proboka, G.; Auziņš, J.; Alberts, P.; Jonsson, B.; Venskus, D.; Muceniece, A. Adapted ECHO-7 virus Rigvir immunotherapy (oncolytic virotherapy) prolongs survival in melanoma patients after surgical excision of the tumour in a retrospective study. Melanoma Res. 2015, 25, 421–426. [Google Scholar] [CrossRef]
- Gruenert, D.C.; Basbaum, C.B.; Welsh, M.J.; Li, M.; Finkbeiner, W.E.; Nadel, J.A. Characterization of human tracheal epithelial cells transformed by an origin-defective simian virus 40. Proc. Natl. Acad. Sci. USA 1988, 85, 5951–5955. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Stringer, C.; Wang, T.; Michaelos, M.; Pachitariu, M. Cellpose: A generalist algorithm for cellular segmentation. Nat. Methods 2021, 18, 100–106. [Google Scholar] [CrossRef]
- Peltola, V.; Waris, M.; Osterback, R.; Susi, P.; Ruuskanen, O.; Hyypiä, T. Rhinovirus transmission within families with children: Incidence of symptomatic and asymptomatic infections. J. Infect. Dis. 2008, 197, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Lönnrot, M.; Sjöroos, M.; Salminen, K.; Maaronen, M.; Hyypiä, T.; Hyöty, H. Diagnosis of enterovirus and rhinovirus infections by RT-PCR and time-resolved fluorometry with lanthanide chelate labeled probes. J. Med. Virol. 1999, 59, 378–384. [Google Scholar] [CrossRef]
- McLeish, N.J.; Witteveldt, J.; Clasper, L.; McIntyre, C.; McWilliam Leitch, E.C.; Hardie, A.; Bennett, S.; Gunson, R.; Carman, W.F.; Feeney, S.A.; et al. Development and Assay of RNA Transcripts of Enterovirus Species A to D, Rhinovirus Species A to C, and Human Parechovirus: Assessment of Assay Sensitivity and Specificity of Real-Time Screening and Typing Methods. J. Clin. Microbiol. 2012, 50, 2910–2917. [Google Scholar] [CrossRef]
- Hietanen, E.; Smura, T.; Hakanen, M.; Chansaenroj, J.; Merilahti, P.; Nevalainen, J.; Pandey, S.; Koskinen, S.; Tripathi, L.; Poovorawan, Y.; et al. Genome Sequences of RIGVIR Oncolytic Virotherapy Virus and Five Other Echovirus 7 Isolates. Genome Announc. 2018, 6, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Chua, B.H.; McMinn, P.C.; Lam, S.K.; Chua, K.B. Comparison of the complete nucleotide sequences of echovirus 7 strain UMMC and the prototype (Wallace) strain demonstrates significant genetic drift over time. J. Gen. Virol. 2001, 82, 2629–2639. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]
- Plevka, P.; Hafenstein, S.; Harris, K.G.; Cifuente, J.O.; Zhang, Y.; Bowman, V.D.; Chipman, P.R.; Bator, C.M.; Lin, F.; Medof, M.E.; et al. Interaction of Decay-Accelerating Factor with Echovirus 7. J. Virol. 2010, 84, 12665–12674. [Google Scholar] [CrossRef]
- Tilgase, A.; Patetko, L.; Blāķe, I.; Ramata-Stunda, A.; Borodušķis, M.; Alberts, P. Effect of the oncolytic ECHO-7 virus Rigvir® on the viability of cell lines of human origin in vitro. J. Cancer 2018, 9, 1033–1049. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, M.; Tolf, C.; Jonsson, N.; Polacek, C.; Precechtelova, J.; Badurova, M.; Sojka, M.; Mohlin, C.; Israelsson, S.; Johansson, K.; et al. A Single Coxsackievirus B2 Capsid Residue Controls Cytolysis and Apoptosis in Rhabdomyosarcoma Cells. J. Virol. 2010, 84, 5868–5879. [Google Scholar] [CrossRef] [PubMed]
- Cordey, S.; Petty, T.J.; Schibler, M.; Martinez, Y.; Gerlach, D.; van Belle, S.; Turin, L.; Zdobnov, E.; Kaiser, L.; Tapparel, C. Identification of site-specific adaptations conferring increased neural cell tropism during human enterovirus 71 infection. PLoS Pathog. 2012, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Yee, P.T.I.; Tan, K.O.; Othman, I.; Poh, C.L. Identification of molecular determinants of cell culture growth characteristics of Enterovirus 71. Virol. J. 2016, 13, 194. [Google Scholar] [CrossRef]
- Yee, P.T.I.; Mohamed, R.A.H.; Ong, S.-K.; Tan, K.O.; Poh, C.L. Characterization of significant molecular determinants of virulence of Enterovirus 71 sub-genotype B4 in Rhabdomyosarcoma cells. Virus Res. 2017, 238, 243–252. [Google Scholar] [CrossRef][Green Version]
- Mikesch, J.-H.; Buerger, H.; Simon, R.; Brandt, B. Decay-accelerating factor (CD55): A versatile acting molecule in human malignancies. Biochim. Biophys. Acta-Rev. Cancer 2006, 1766, 42–52. [Google Scholar] [CrossRef]
- Rossmann, M.G.; Arnold, E.; Erickson, J.W.; Frankenberger, E.A.; Griffith, J.P.; Hecht, H.J.; Johnson, J.E.; Kamer, G.; Luo, M.; Mosser, A.G. Structure of a human common cold virus and functional relationship to other picornaviruses. Nature 1985, 317, 145–153. [Google Scholar] [CrossRef]
- Israelsson, S.; Jonsson, N.; Gullberg, M.; Lindberg, A.M. Cytolytic replication of echoviruses in colon cancer cell lines. Virol. J. 2011, 8, 473. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Capsid Protein | Echovirus 7 Residue | Structure Position | Rigvir® Residue | Surface Exposure |
|---|---|---|---|---|
| 1 | I/L | 76 | F | Yes |
| 1 | V | 153 | I | Yes |
| 2 | L | 17 | H | No |
| 2 | K/N/L | 138 | R | Yes |
| 2 | E/Q/N | 162 | G | Yes |
| 2 | V | 176 | I | No |
| 2 | T/A | 231 | N | Yes |
| 3 | I | 158 | L | No |
| 3 | A | 235 | T | Yes |
| Isolate | GenBank Acc. No. | Cell Line | |||||||
|---|---|---|---|---|---|---|---|---|---|
| HeLa | A549 | HFF | SW480 | Huh7 | U373MG | 16HBE14o | MCF-7 | ||
| Wallace | MH043132 | −/+ | +++ | +++ | ++ | +++ | ++ | ++ | − |
| Rigvir® | MH043137 | + | ++ | +++ | +++ | +++ | −/+ | +++ | − |
| 98-57213 | MH043133 | + | ++ | ++ | ++ | ++ | ++ | +++ | − |
| 98-59065 | MH043134 | + | + | ++ | + | +++ | + | +++ | − |
| 98-60628 | MH043135 | + | + | + | −/+ | ++ | −/+ | + | − |
| 07VI447 | MH043136 | + | +++ | ++ | + | + | −/+ | +++ | − |
| Isolate | Cell Line | |||||||
|---|---|---|---|---|---|---|---|---|
| HeLa | A549 | HFF | SW480 | Huh7 | U373MG | 16HBE14o | MCF-7 | |
| Wallace | 0.9 | 4.1 | 15.5 | 4.0 | 11.1 | 1.6 | 2.9 | 0.0 |
| 98-57213 | 2.9 | 19.4 | 83.5 | 7.3 | 36.6 | 5.2 | 39.2 | 0.0 |
| 98-59065 | 3.6 | 7.8 | 16.0 | 1.6 | 7.8 | 0.4 | 43.9 | 0.0 |
| 98-60628 | 2.7 | 4.6 | 43.7 | 0.5 | 2.4 | 0.4 | 0.3 | 0.0 |
| 07VI447 | 1.6 | 42.6 | 28.4 | 1.4 | 40.6 | 0.2 | 44.1 | 0.0 |
| Rigvir® | 1.1 | 9.3 | 31.4 | 7.1 | 48.9 | 0.4 | 11.7 | 0.0 |
| Control | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hietanen, E.; Koivu, M.K.A.; Susi, P. Cytolytic Properties and Genome Analysis of Rigvir® Oncolytic Virotherapy Virus and Other Echovirus 7 Isolates. Viruses 2022, 14, 525. https://doi.org/10.3390/v14030525
Hietanen E, Koivu MKA, Susi P. Cytolytic Properties and Genome Analysis of Rigvir® Oncolytic Virotherapy Virus and Other Echovirus 7 Isolates. Viruses. 2022; 14(3):525. https://doi.org/10.3390/v14030525
Chicago/Turabian StyleHietanen, Eero, Marika K. A. Koivu, and Petri Susi. 2022. "Cytolytic Properties and Genome Analysis of Rigvir® Oncolytic Virotherapy Virus and Other Echovirus 7 Isolates" Viruses 14, no. 3: 525. https://doi.org/10.3390/v14030525
APA StyleHietanen, E., Koivu, M. K. A., & Susi, P. (2022). Cytolytic Properties and Genome Analysis of Rigvir® Oncolytic Virotherapy Virus and Other Echovirus 7 Isolates. Viruses, 14(3), 525. https://doi.org/10.3390/v14030525