
DEAD-Box RNA Helicase 21 (DDX21) Positively Regulates the Replication of Porcine Reproductive and Respiratory Syndrome Virus via Multiple Mechanisms

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus, and Reagents
2.2. Plasmid Construction
2.3. siRNA and Transfection
2.4. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
2.5. TCID50 Assay
2.6. Indirect Immunofluorescence Assay (IFA)
2.7. Western Blot Analysis
2.8. Co-Immunoprecipitation (Co-IP) Assay
2.9. Nuclear Cytosol Fractionation Assay
2.10. Statistical Analysis
3. Results
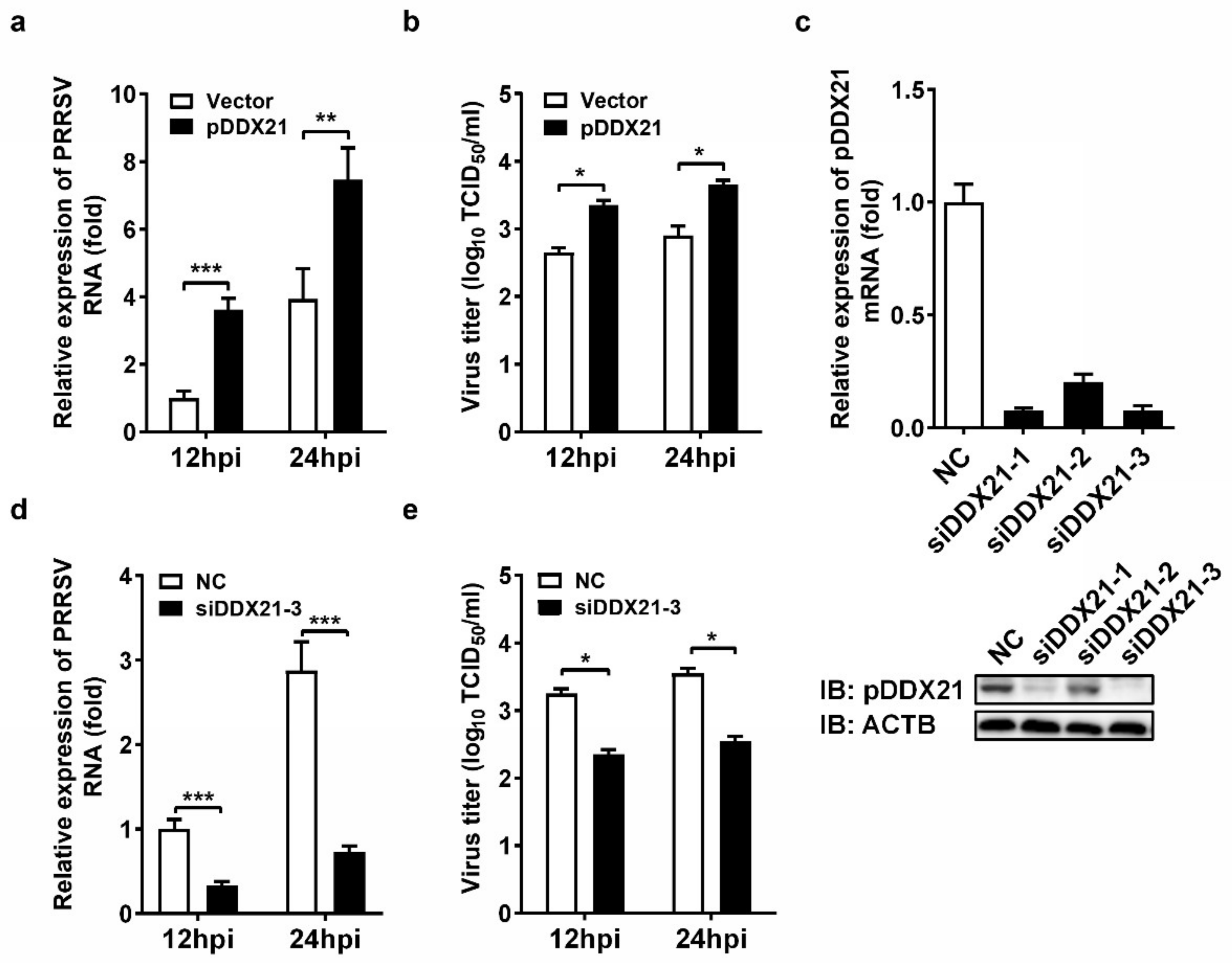
3.1. pDDX21 Positively Regulates PRRSV Replication in iPAM Cells
3.2. pDDX21 Promotes PRRSV Replication Independently of Its ATPase, RNA Helicase, and Foldase Activity
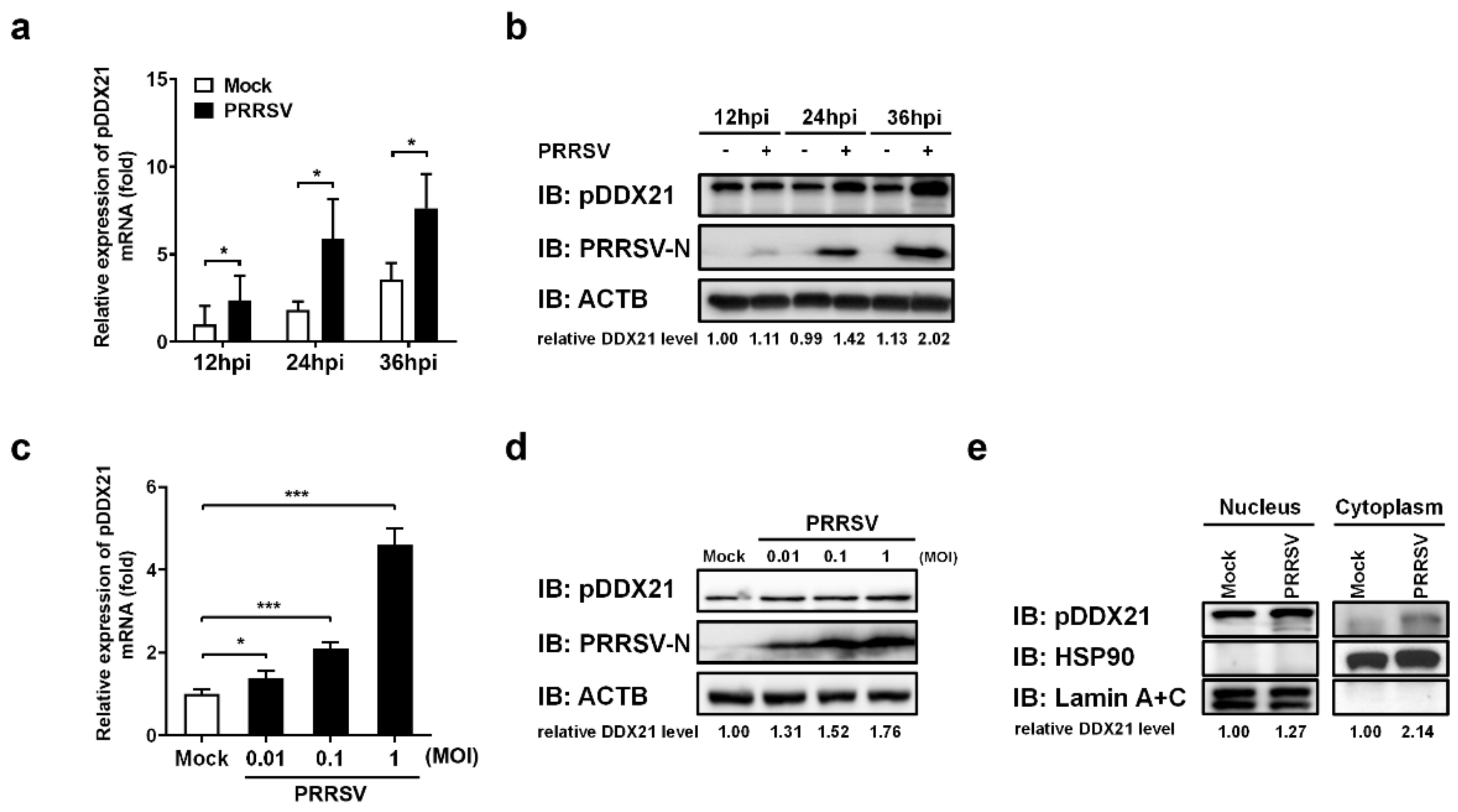
3.3. PRRSV Infection Promotes pDDX21 Translocation from the Nucleus to the Cytoplasm
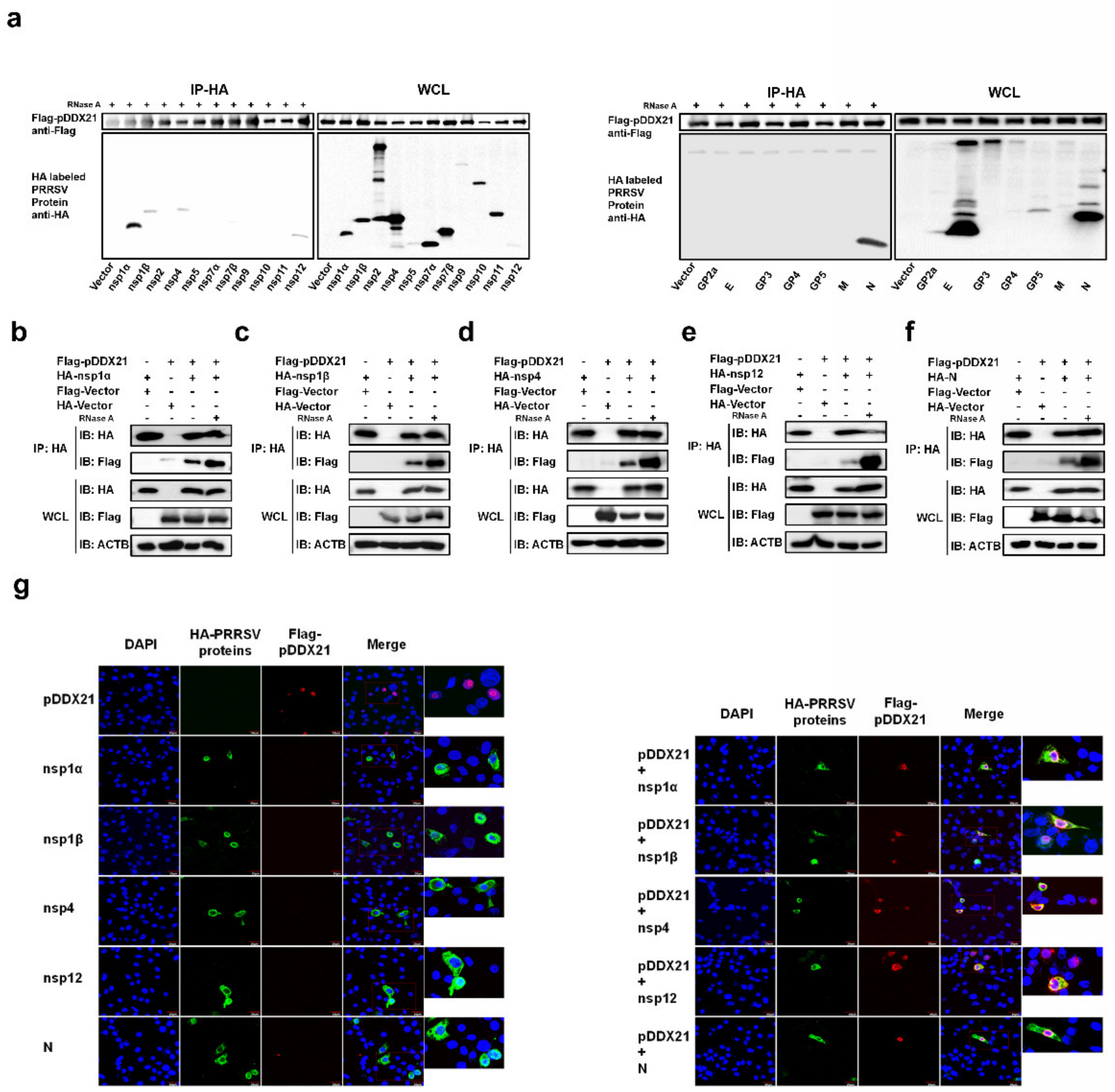
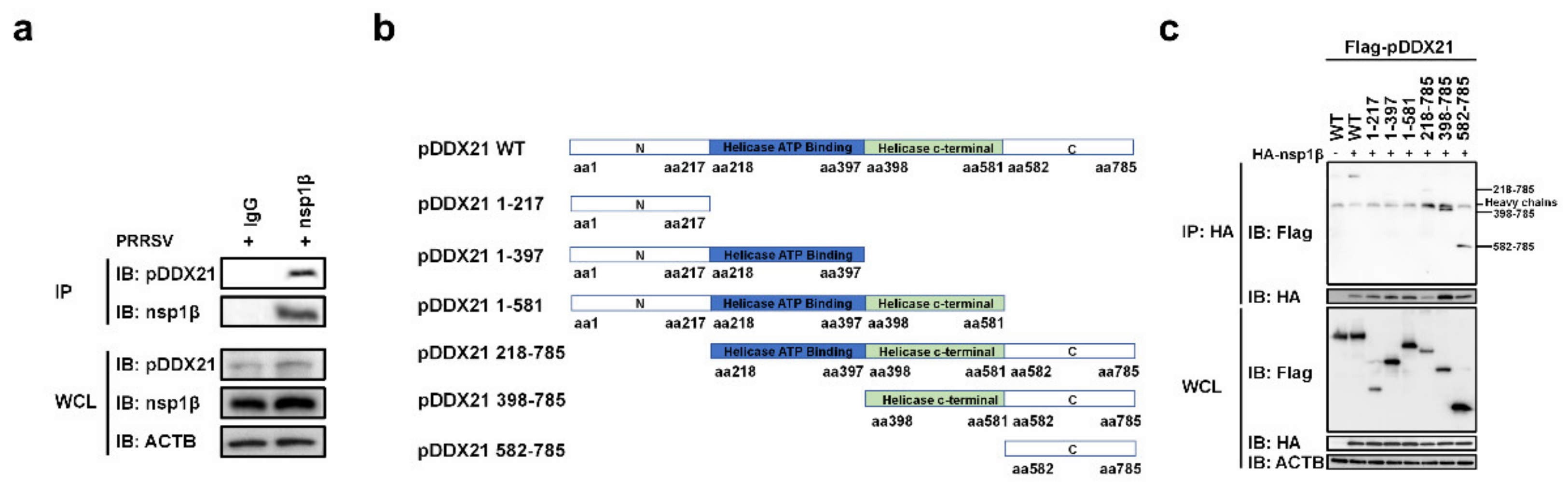
3.4. pDDX21 Interacts with Multiple PRRSV-Encoded Proteins
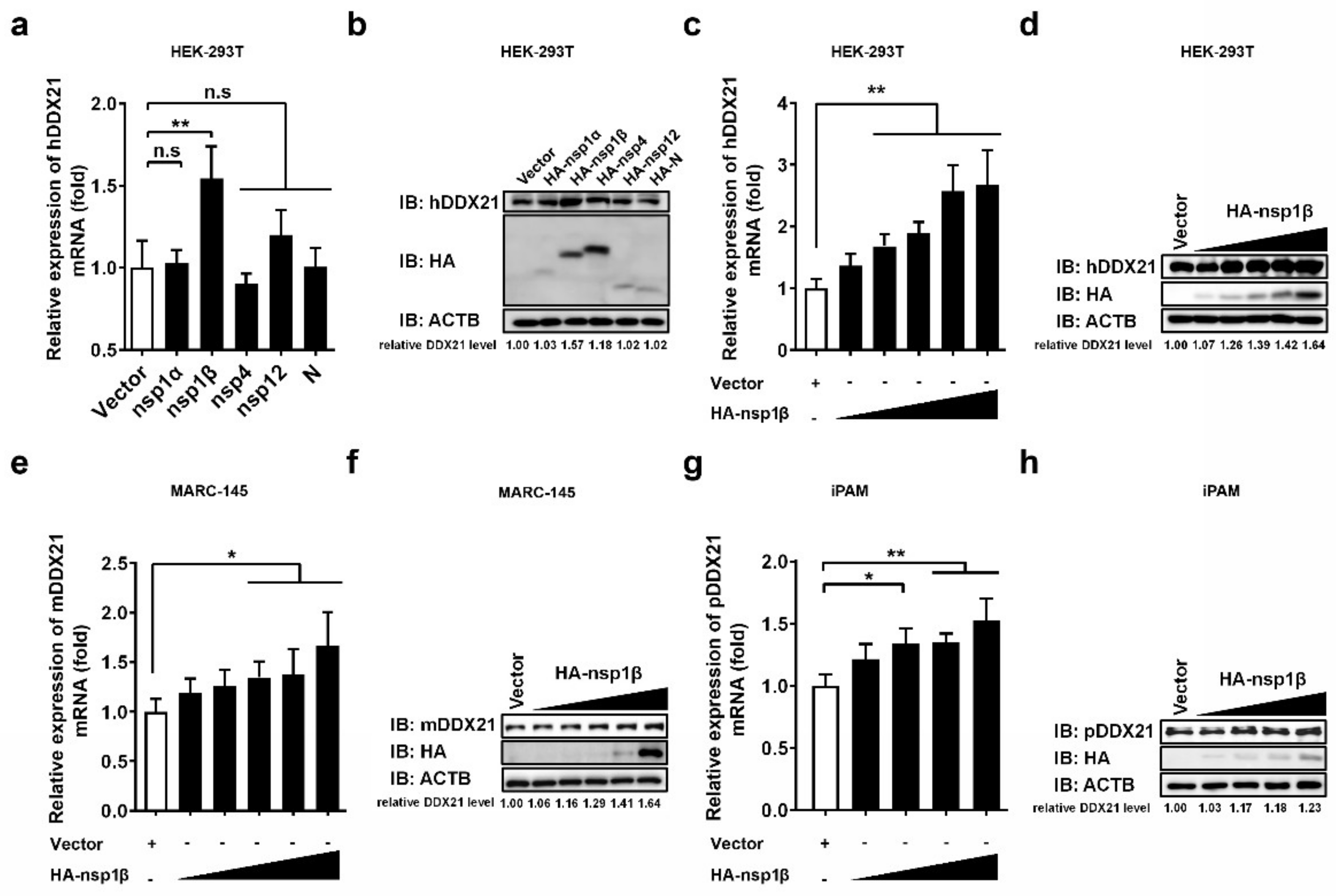
3.5. PRRSV nsp1β Upregulates DDX21 Transcription and Protein Expression
3.6. nsp1β Interacts with the C-Terminal Domain of pDDX21
3.7. pDDX21 Stabilizes the Expression of PRRSV nsp1α, nsp1β, and N Protein
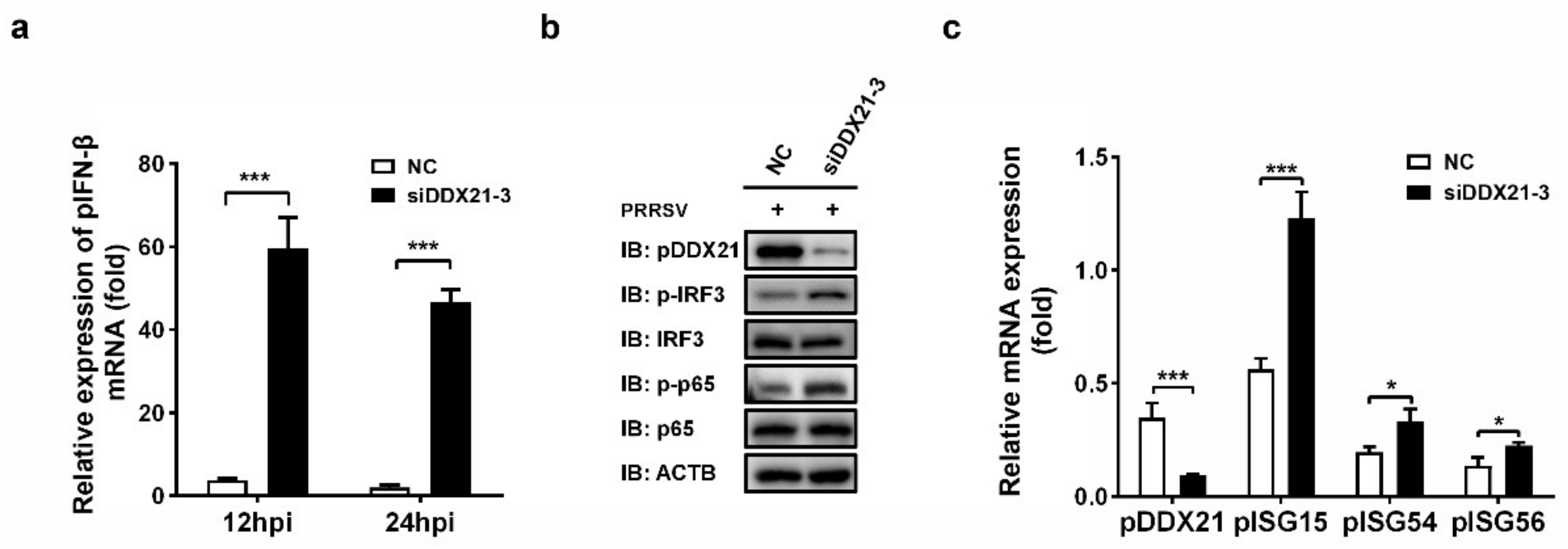
3.8. Knockdown of pDDX21 Activates the IFN-β Signaling Pathway in PRRSV-Infected Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christianson, W.T.; Collins, J.E.; Benfield, D.A.; Harris, L.; Joo, H.S. Experimental reproduction of swine infertility and respiratory syndrome in pregnant sows. Am. J. Vet. Res. 1992, 53, 485–488. [Google Scholar]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479, 475–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wensvoort, G.; Terpstra, C.; Pol, J.M. ‘Lelystad agent’—The cause of abortus blauw (mystery swine disease). Tijdschr. Diergeneeskd. 1991, 116, 675. [Google Scholar] [PubMed]
- Meulenberg, J.; Hulst, M.M.; Meijer, E.; Moonen, P.; Besten, A.D.; Kluyver, E.; Wensvoort, G.; Moormann, R. Lelystad virus, the causative agent of porcine epidemic abortion and respiratory syndrome (PEARS), is related to LDV and EAV. Virology 1993, 192, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Snijder, E.J. The PRRSV replicase: Exploring the multifunctionality of an intriguing set of nonstructural proteins. Virus Res. 2010, 154, 61–76. [Google Scholar] [CrossRef]
- Li, Y.; Tas, A.; Snijder, E.J.; Fang, Y. Identification of porcine reproductive and respiratory syndrome virus ORF1a-encoded non-structural proteins in virus-infected cells. J. Gen. Virol. 2012, 93, 829–839. [Google Scholar] [CrossRef]
- Li, Y.; Tas, A.; Sun, Z.; Snijder, E.J.; Fang, Y. Proteolytic processing of the porcine reproductive and respiratory syndrome virus replicase. Virus Res. 2015, 202, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Snijder, E.J.; Kikkert, M. Arteriviruses. Encyclopedia of Virology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 176–186. [Google Scholar]
- Firth, J. Rheumatoid arthritis: Diagnosis and multidisciplinary management. Br. J. Nurs. 2011, 20, 1179–1180. [Google Scholar] [CrossRef]
- Overend, C.; Mitchell, R.; He, D.; Rompato, G.; Grubman, M.J.; Garmendia, A.E. Recombinant swine beta interferon protects swine alveolar macrophages and MARC-145 cells from infection with Porcine reproductive and respiratory syndrome virus. J. Virol. 2007, 88, 925–931. [Google Scholar] [CrossRef]
- Beura, L.K.; Sarkar, S.N.; Kwon, B.; Subramaniam, S.; Jones, C.; Pattnaik, A.K.; Osorio, F.A. Porcine reproductive and respiratory syndrome virus nonstructural protein 1beta modulates host innate immune response by antagonizing IRF3 activation. J. Virol. 2010, 84, 1574–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagong, M.; Lee, C. Porcine reproductive and respiratory syndrome virus nucleocapsid protein modulates interferon-β production by inhibiting IRF3 activation in immortalized porcine alveolar macrophages. Arch. Virol. 2011, 156, 2187–2195. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Nan, Y.; Yu, Y.; Zhang, Y.J. Porcine reproductive and respiratory syndrome virus Nsp1β inhibits interferon-activated JAK/STAT signal transduction by inducing karyopherin-α1 degradation. J. Virol. 2013, 87, 5219–5228. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zheng, Z.; Zhou, P.; Zhang, B.; Shi, Z.; Hu, Q.; Wang, H. The cysteine protease domain of porcine reproductive and respiratory syndrome virus non-structural protein 2 antagonizes interferon regulatory factor 3 activation. J. Gen. Virol. 2010, 91, 2947–2958. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, D.; Sun, Z.; Gao, L.; Zhu, X.; Guo, J.; Xu, S.; Fang, L.; Li, K.; Xiao, S. Arterivirus nsp4 Antagonizes Interferon Beta Production by Proteolytically Cleaving NEMO at Multiple Sites. J. Virol. 2019, 93, e00385-19. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Chen, J.; Yu, C.; Zhu, X.; Xu, S.; Fang, L.; Xiao, S. Porcine Reproductive and Respiratory Syndrome Virus nsp11 Antagonizes Type I Interferon Signaling by Targeting IRF9. J. Virol. 2019, 93, e00623-19. [Google Scholar] [CrossRef] [Green Version]
- Linder, P.; Jankowsky, E. From unwinding to clamping—The DEAD box RNA helicase family. Nat. Rev. Mol. Cell Biol. 2011, 12, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Diot, C.; Fournier, G.; Dos Santos, M.; Magnus, J.; Komarova, A.; van der Werf, S.; Munier, S.; Naffakh, N. Influenza A Virus Polymerase Recruits the RNA Helicase DDX19 to Promote the Nuclear Export of Viral mRNAs. Sci. Rep. 2016, 6, 33763. [Google Scholar] [CrossRef]
- Ullah, R.; Li, J.; Fang, P.; Shaobo, X.; Fang, L. DEAD/H-box helicases: Anti-viral and pro-viral roles during infections. Virus Res. 2021, 309, 198658. [Google Scholar] [CrossRef] [PubMed]
- Meier-Stephenson, V.; Mrozowich, T.; Pham, M.; Patel, T.R. DEAD-box helicases: The Yin and Yang roles in viral infections. Biotechnol. Genet. Eng. Rev. 2018, 34, 3–32. [Google Scholar] [CrossRef]
- Taschuk, F.; Cherry, S. DEAD-Box Helicases: Sensors, Regulators, and Effectors for Antiviral Defense. Viruses 2020, 12, 181. [Google Scholar] [CrossRef] [Green Version]
- Dehghani, M.; Lasko, P. Multiple Functions of the DEAD-Box Helicase Vasa in Drosophila Oogenesis. Results Probl. Cell Differ. 2017, 63, 127–147. [Google Scholar] [PubMed]
- Ali, M.A.M. The DEAD-box protein family of RNA helicases: Sentinels for a myriad of cellular functions with emerging roles in tumorigenesis. Int. J. Clin. Oncol. 2021, 26, 795–825. [Google Scholar] [CrossRef] [PubMed]
- McRae, E.K.S.; Booy, E.P.; Moya-Torres, A.; Ezzati, P.; Stetefeld, J.; McKenna, S.A. Human DDX21 binds and unwinds RNA guanine quadruplexes. Nucleic Acids Res. 2017, 45, 6656–6668. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.W.; Ye, W.; Chen, H.S.; Nie, T.J.; Cheng, L.F.; Zhang, L.; Han, P.J.; Wu, X.A.; Xu, Z.K.; Lei, Y.F.; et al. In-Cell Western Assays to Evaluate Hantaan Virus Replication as a Novel Approach to Screen Antiviral Molecules and Detect Neutralizing Antibody Titers. Front. Cell Infect. Microbiol. 2017, 7, 269. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Ye, W.; Yang, J.; Han, P.; Wang, Y.; Ye, C.; Weng, D.; Zhang, F.; Xu, Z.; Lei, Y. DDX21 translocates from nucleus to cytoplasm and stimulates the innate immune response due to dengue virus infection. Biochem. Biophys. Res. Commun. 2016, 473, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Qu, Y.; Yu, S.; Wang, S.; Yin, Y.; Liu, Q.; Meng, C.; Liao, Y.; Ur Rehman, Z.; Tan, L.; et al. Caspase-Dependent Cleavage of DDX21 Suppresses Host Innate Immunity. mBio 2021, 12, e0100521. [Google Scholar] [CrossRef]
- Chen, G.; Liu, C.H.; Zhou, L.; Krug, R.M. Cellular DDX21 RNA helicase inhibits influenza A virus replication but is counteracted by the viral NS1 protein. Cell Host Microbe. 2014, 15, 484–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naji, S.; Ambrus, G.; Cimermančič, P.; Reyes, J.R.; Johnson, J.R.; Filbrandt, R.; Huber, M.D.; Vesely, P.; Krogan, N.J.; Yates, J.R., 3rd; et al. Host cell interactome of HIV-1 Rev includes RNA helicases involved in multiple facets of virus production. Mol. Cell Proteom. 2012, 11, M111.015313. [Google Scholar] [CrossRef] [Green Version]
- Hao, H.; Han, T.; Xuan, B.; Sun, Y.; Tang, S.; Yue, N.; Qian, Z. Dissecting the Role of DDX21 in Regulating Human Cytomegalovirus Replication. J. Virol. 2019, 93, e01222-19. [Google Scholar] [CrossRef]
- Wang, T.Y.; Liu, Y.G.; Li, L.; Wang, G.; Wang, H.M.; Zhang, H.L.; Zhao, S.F.; Gao, J.C.; An, T.Q.; Tian, Z.J.; et al. Porcine alveolar macrophage CD163 abundance is a pivotal switch for porcine reproductive and respiratory syndrome virus infection. Oncotarget 2018, 9, 12174–12185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Fang, L.; Liu, S.; Zhao, F.; Jiang, Y.; He, K.; Chen, H.; Xiao, S. The genomic diversity of Chinese porcine reproductive and respiratory syndrome virus isolates from 1996 to 2009. Vet. Microbiol. 2010, 146, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Fang, L.; Wang, D.; Zhang, R.; Zeng, S.; An, K.; Chen, H.; Xiao, S. Quantitative interactome reveals that porcine reproductive and respiratory syndrome virus nonstructural protein 2 forms a complex with viral nucleocapsid protein and cellular vimentin. J. Proteom. 2016, 142, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Fang, L.; Tao, R.; Li, Y.; Jing, H.; Wang, D.; Xiao, S. Porcine Reproductive and Respiratory Syndrome Virus E Protein Degrades Porcine Cholesterol 25-Hydroxylase via the Ubiquitin-Proteasome Pathway. J. Virol. 2019, 93, e00767-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, W.; Fang, L.; Jing, H.; Tao, R.; Wang, T.; Li, Y.; Long, S.; Wang, D.; Xiao, S. Cholesterol 25-Hydroxylase Inhibits Porcine Reproductive and Respiratory Syndrome Virus Replication through Enzyme Activity-Dependent and -Independent Mechanisms. J. Virol. 2017, 91, e00827-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzi, M. Sampling variation of the fifty percent end-point, determined by the Reed-Muench (Behrens) method. Hum. Biol. 1950, 22, 151–190. [Google Scholar]
- Valdez, B.C. Structural domains involved in the RNA folding activity of RNA helicase II/Gu protein. Eur. J. Biochem. 2000, 267, 6395–6402. [Google Scholar] [CrossRef] [PubMed]
- Valdez, B.C.; Henning, D.; Perumal, K.; Busch, H. RNA-unwinding and RNA-folding activities of RNA helicase II/Gu—Two activities in separate domains of the same protein. Eur. J. Biochem. 1997, 250, 800–807. [Google Scholar] [CrossRef]
- Ou, Y.; Fritzler, M.J.; Valdez, B.C.; Rattner, J.B. Mapping and characterization of the functional domains of the nucleolar protein RNA helicase II/Gu. Exp. Cell Res. 1999, 247, 389–398. [Google Scholar] [CrossRef]
- Zhang, Z.; Kim, T.; Bao, M.; Facchinetti, V.; Jung, S.Y.; Ghaffari, A.A.; Qin, J.; Cheng, G.; Liu, Y.J. DDX1, DDX21, and DHX36 helicases form a complex with the adaptor molecule TRIF to sense dsRNA in dendritic cells. Immunity 2011, 34, 866–878. [Google Scholar] [CrossRef] [Green Version]
- Calo, E.; Flynn, R.A.; Martin, L.; Spitale, R.C.; Chang, H.Y.; Wysocka, J. RNA helicase DDX21 coordinates transcription and ribosomal RNA processing. Nature 2015, 518, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Liu, S.; Sun, W.; Chen, L.; Yoo, D.; Li, F.; Ren, S.; Guo, L.; Cong, X.; Li, J. Nuclear export signal of PRRSV NSP1α is necessary for type I IFN inhibition. Virology 2016, 499, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Zhou, L.; Ge, X.; Zhang, H.; Zhang, R.; Wang, C.; Wang, L.; Zhang, Z.; Yang, H.; Guo, X. Cellular DEAD-box RNA helicase 18 (DDX18) Promotes the PRRSV Replication via Interaction with Virus nsp2 and nsp10. Virus Res. 2017, 238, 204–212. [Google Scholar] [CrossRef]
- Zhao, S.; Ge, X.; Wang, X.; Liu, A.; Guo, X.; Zhou, L.; Yu, K.; Yang, H. The DEAD-box RNA helicase 5 positively regulates the replication of porcine reproductive and respiratory syndrome virus by interacting with viral Nsp9 in vitro. Virus Res. 2015, 195, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, Q.; Liu, D.; Wang, D.; Chen, H.; Xiao, S.; Fang, L. Molecular cloning, functional characterization and antiviral activity of porcine DDX3X. Biochem. Biophys. Res. Commun. 2014, 443, 1169–1175. [Google Scholar] [CrossRef]
- Liu, L.; Tian, J.; Nan, H.; Tian, M.; Li, Y.; Xu, X.; Huang, B.; Zhou, E.; Hiscox, J.A.; Chen, H. Porcine Reproductive and Respiratory Syndrome Virus Nucleocapsid Protein Interacts with Nsp9 and Cellular DHX9 To Regulate Viral RNA Synthesis. J. Virol. 2016, 90, 5384–5398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Hu, L.; Liu, Y.; Huang, L.; Mu, Y.; Cai, X.; Weng, C. DDX19A Senses Viral RNA and Mediates NLRP3-Dependent Inflammasome Activation. J. Immunol. 2015, 195, 5732–5749. [Google Scholar] [CrossRef] [Green Version]
- Holmström, T.H.; Mialon, A.; Kallio, M.; Nymalm, Y.; Mannermaa, L.; Holm, T.; Johansson, H.; Black, E.; Gillespie, D.; Salminen, T.A.; et al. c-Jun Supports Ribosomal RNA Processing and Nucleolar Localization of RNA Helicase DDX21. J. Biol. Chem. 2008, 283, 7046–7053. [Google Scholar] [CrossRef] [Green Version]
- Henning, D.; So, R.B.; Jin, R.; Lau, L.F.; Valdez, B.C. Silencing of RNA Helicase II/Guα Inhibits Mammalian Ribosomal RNA Production. J. Biol. Chem. 2003, 278, 52307–52314. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Du, Y.; Song, C.; Yoo, D. Degradation of CREB-binding protein and modulation of type I interferon induction by the zinc finger motif of the porcine reproductive and respiratory syndrome virus nsp1α subunit. Virus Res. 2013, 172, 54–65. [Google Scholar] [CrossRef]
- Han, M.; Ke, H.; Zhang, Q.; Yoo, D. Nuclear imprisonment of host cellular mRNA by nsp1β protein of porcine reproductive and respiratory syndrome virus. Virology 2017, 505, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Ge, X.; Liu, Y.; Jiang, P.; Wang, Z.; Zhang, R.; Zhou, L.; Guo, X.; Han, J.; Yang, H. Targeting Swine Leukocyte Antigen Class I Molecules for Proteasomal Degradation by the nsp1α Replicase Protein of the Chinese Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Strain JXwn06. J. Virol. 2016, 90, 682–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, H.; Han, M.; Kim, J.; Gustin, K.E.; Yoo, D. Porcine Reproductive and Respiratory Syndrome Virus Nonstructural Protein 1 Beta Interacts with Nucleoporin 62 To Promote Viral Replication and Immune Evasion. J. Virol. 2019, 93, e00469-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Li, L.-W.; Jiang, Y.-F.; Gao, F.; Zhang, Y.-J.; Zhao, W.-Y.; Li, G.-X.; Yu, L.-X.; Zhou, Y.-J.; Tong, G.-Z. Nucleocapsid protein of porcine reproductive and respiratory syndrome virus antagonizes the antiviral activity of TRIM25 by interfering with TRIM25-mediated RIG-I ubiquitination. Vet. Microbiol. 2019, 233, 140–146. [Google Scholar] [CrossRef]
- Flores-Rozas, H.; Hurwitz, J. Characterization of a new RNA helicase from nuclear extracts of HeLa cells which translocates in the 5′ to 3′ direction. J. Biol. Chem. 1993, 268, 21372–21383. [Google Scholar] [CrossRef]
- Valdez, B.C.; Henning, D.; Busch, R.K.; Woods, K.; Flores-Rozas, H.; Hurwitz, J.; Perlaky, L.; Busch, H. A nucleolar RNA helicase recognized by autoimmune antibodies from a patient with watermelon stomach disease. Nucleic Acids Res. 1996, 24, 1220–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Hotz-Wagenblatt, A.; Voit, R.; Grummt, I. SIRT7 and the DEAD-box helicase DDX21 cooperate to resolve genomic R loops and safeguard genome stability. Genes Dev. 2017, 31, 1370–1381. [Google Scholar] [CrossRef]
- Watanabe, Y.; Ohtaki, N.; Hayashi, Y.; Ikuta, K.; Tomonaga, K. Autogenous translational regulation of the Borna disease virus negative control factor X from polycistronic mRNA using host RNA helicases. PLoS Pathog. 2009, 5, e1000654. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Ge, L.-L.; Li, P.-P.; Wang, Y.; Sun, M.-X.; Huang, L.; Ishag, H.; Di, D.-D.; Shen, Z.-Q.; Fan, W.X. The DEAD-box RNA helicase DDX5 acts as a positive regulator of Japanese encephalitis virus replication by binding to viral 3′ UTR–ScienceDirect. Antivir. Res. 2013, 100, 487–499. [Google Scholar] [CrossRef]
- Abdullah, S.W.; Wu, J.; Zhang, Y.; Bai, M.; Guan, J.; Liu, X.; Sun, S.; Guo, H. DDX21, a Host Restriction Factor of FMDV IRES-Dependent Translation and Replication. Viruses 2021, 13, 1765. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wang, D.; Fang, P.; Pang, Y.; Zhou, Y.; Fang, L.; Xiao, S. DEAD-Box RNA Helicase 21 (DDX21) Positively Regulates the Replication of Porcine Reproductive and Respiratory Syndrome Virus via Multiple Mechanisms. Viruses 2022, 14, 467. https://doi.org/10.3390/v14030467
Li J, Wang D, Fang P, Pang Y, Zhou Y, Fang L, Xiao S. DEAD-Box RNA Helicase 21 (DDX21) Positively Regulates the Replication of Porcine Reproductive and Respiratory Syndrome Virus via Multiple Mechanisms. Viruses. 2022; 14(3):467. https://doi.org/10.3390/v14030467
Chicago/Turabian StyleLi, Jia, Dang Wang, Puxian Fang, Yu Pang, Yanrong Zhou, Liurong Fang, and Shaobo Xiao. 2022. "DEAD-Box RNA Helicase 21 (DDX21) Positively Regulates the Replication of Porcine Reproductive and Respiratory Syndrome Virus via Multiple Mechanisms" Viruses 14, no. 3: 467. https://doi.org/10.3390/v14030467
APA StyleLi, J., Wang, D., Fang, P., Pang, Y., Zhou, Y., Fang, L., & Xiao, S. (2022). DEAD-Box RNA Helicase 21 (DDX21) Positively Regulates the Replication of Porcine Reproductive and Respiratory Syndrome Virus via Multiple Mechanisms. Viruses, 14(3), 467. https://doi.org/10.3390/v14030467