
Transcriptomic Profiling of Mouse Mast Cells upon Pathogenic Avian H5N1 and Pandemic H1N1 Influenza a Virus Infection



Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses and Cell Culture
2.2. In Vitro Viral Infection
2.3. RNA Extraction
2.4. cDNA Preparation and Next Generation Sequencing
2.5. Confirmation of DEGs by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
2.6. RNA Interference (RNAi)
2.7. Western Blotting
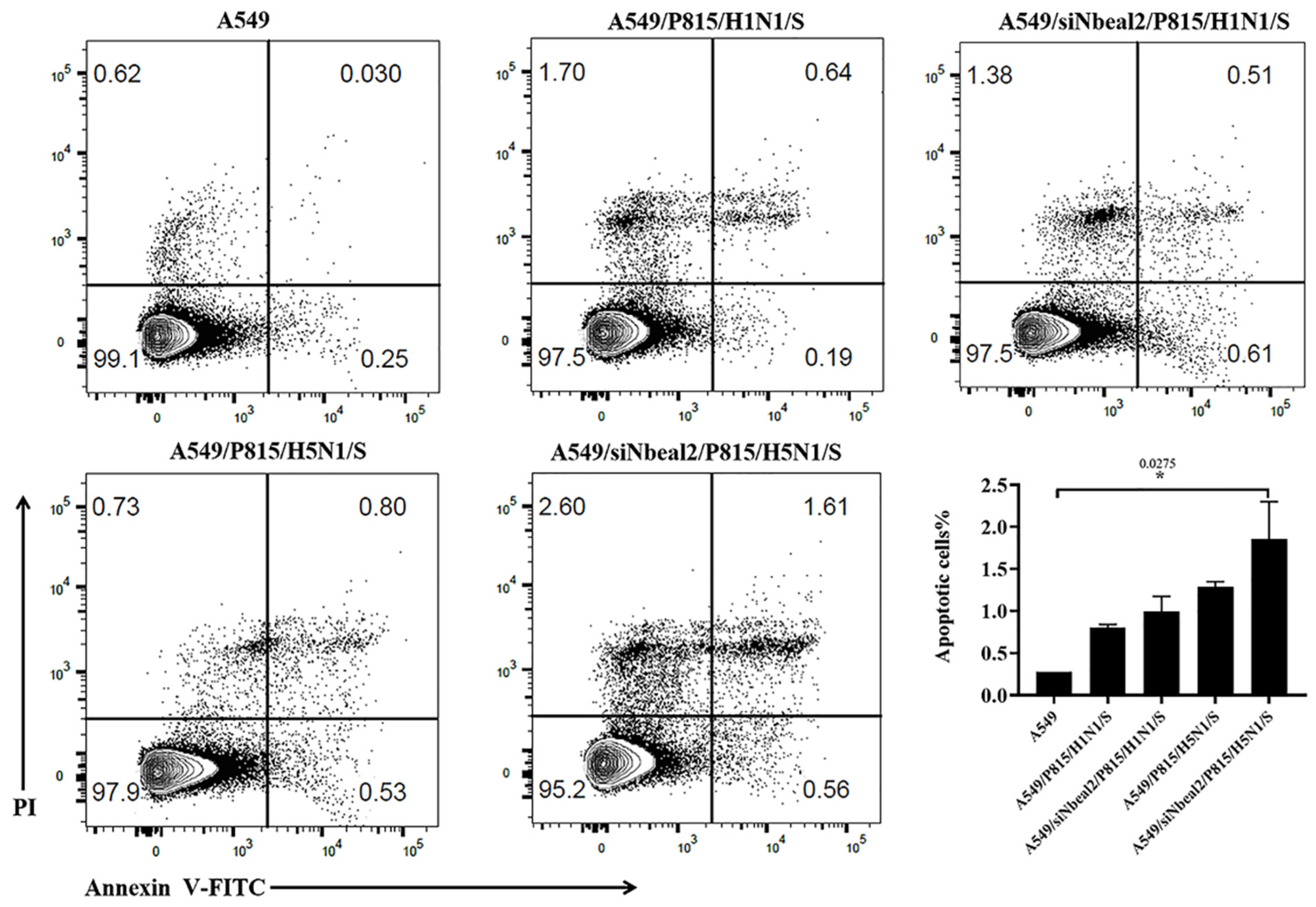
2.8. Analysis of A549 Cell Apoptosis by Flow Cytometry
2.9. Data Statistics and Analysis
3. Results
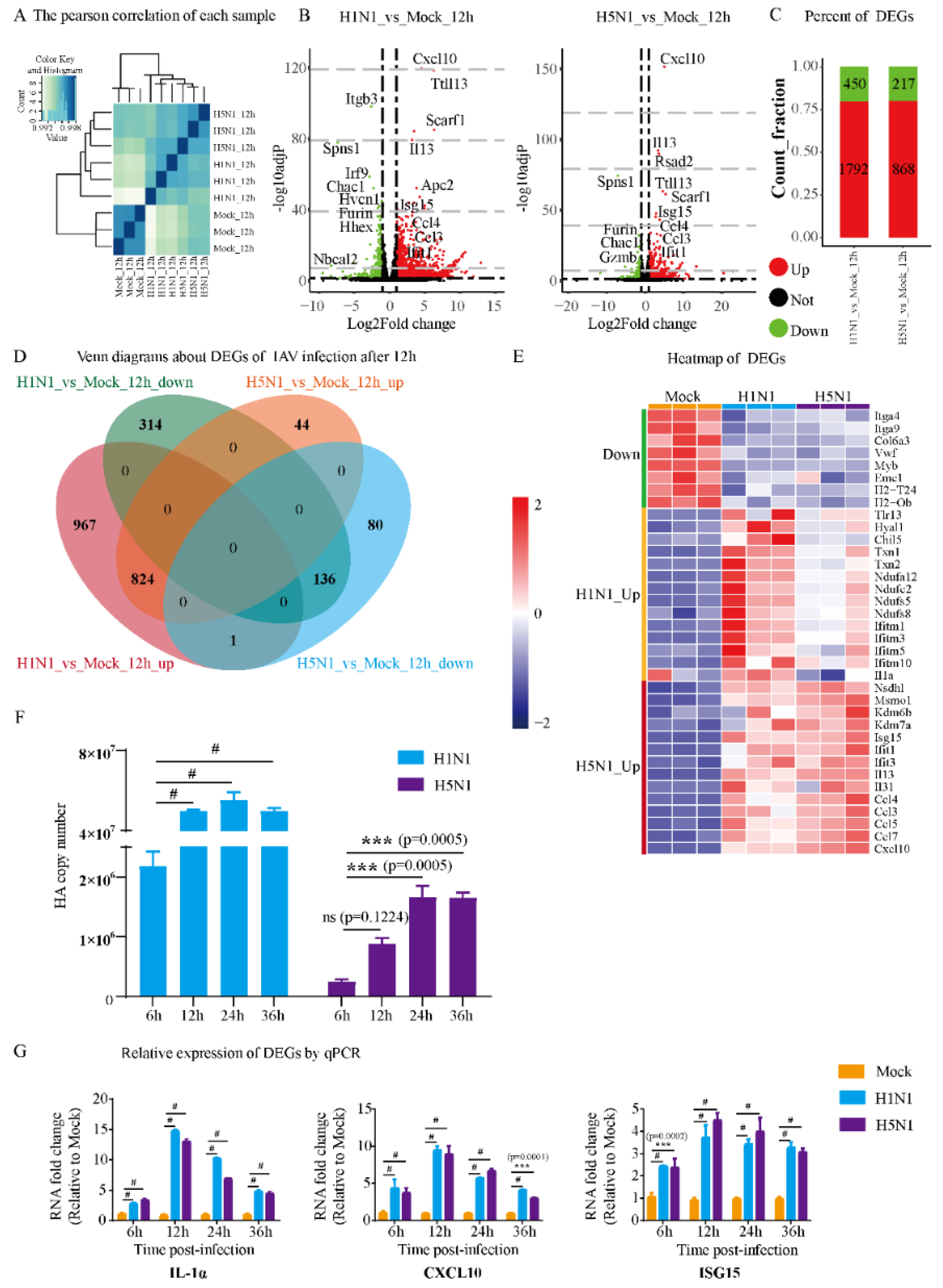
3.1. Gene Transcription Profile of Mast Cells upon IAV Infection
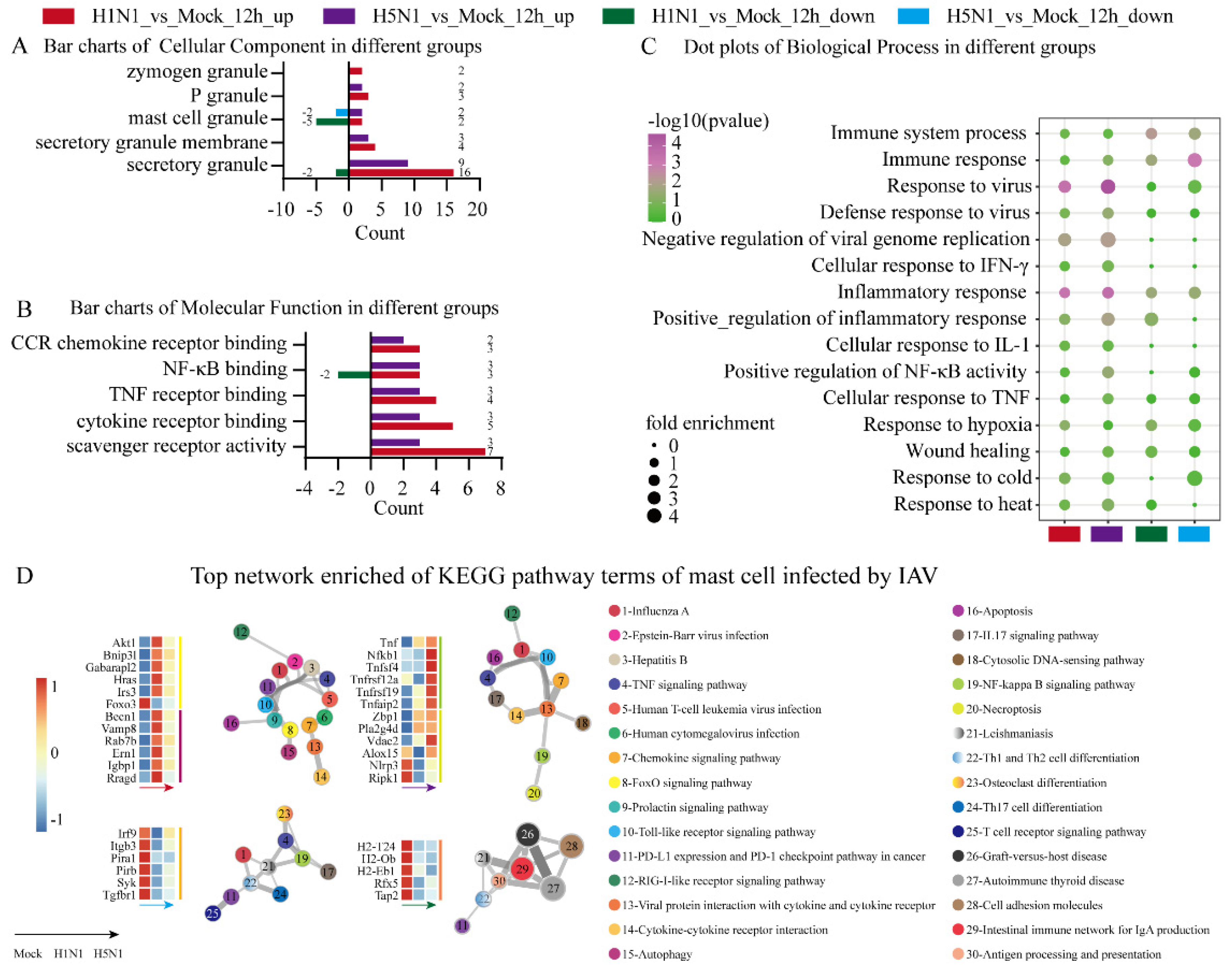
3.2. Gene Ontology Analysis of Mast Cells Infected with H1N1 and H5N1 Viruses
3.3. Comparison of H5N1- and H1N1-Infected Mast Cells
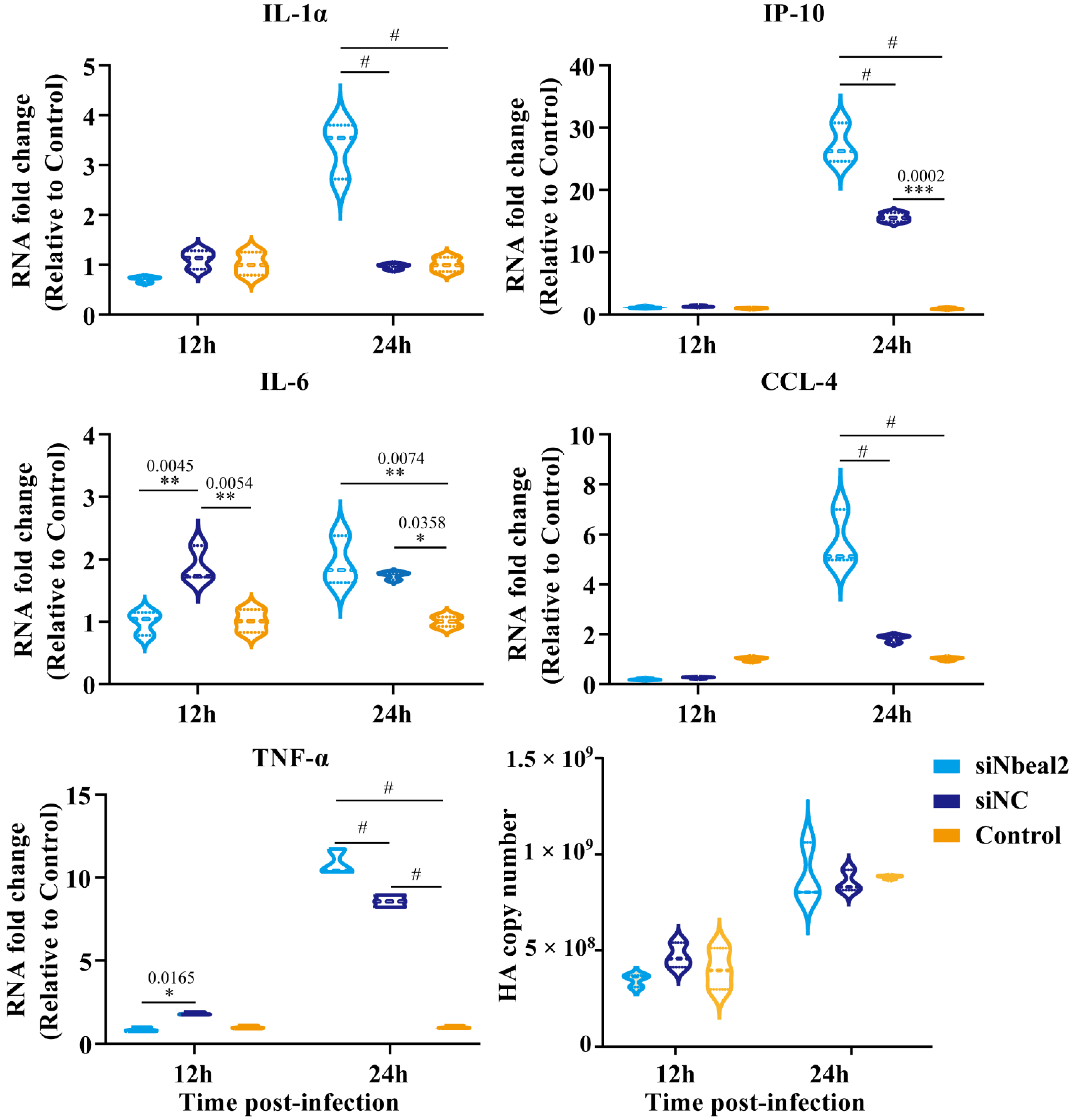
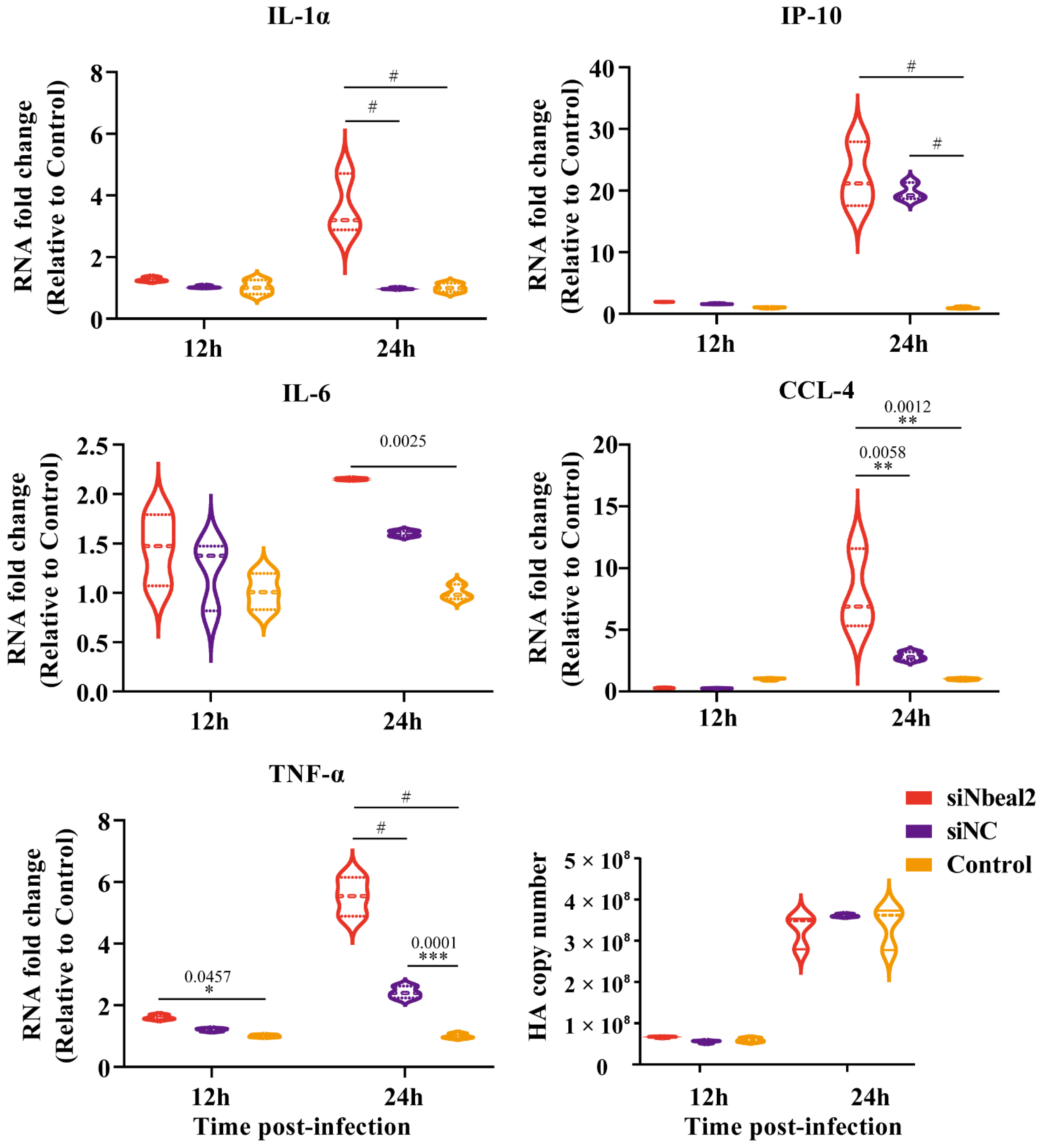
3.4. Effect of Nbeal2 on the Inflammatory Response Following H1N1 and H5N1 Virus Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richard, M.; Fouchier, R.A. Influenza A virus transmission via respiratory aerosols or droplets as it relates to pandemic potential. FEMS Microbiol. Rev. 2016, 40, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Radovanov, J.; Hrnjakovic-Cvjetkovic, I.; Nikolic, N.; Jovanovic-Galovic, A.; Kovacevic, G.; Patic, A.; Milosevic, U.; Milosevic, V. Risk factors for severe influenza a virus infections in post-2009 pandemic period. Srp. Arh. Celok. Lek. 2016, 144, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.Y.; Poon, L.L.; Lau, A.S.; Luk, W.; Lau, Y.L.; Shortridge, K.F.; Gordon, S.; Guan, Y.; Peiris, J.S. Induction of proinflammatory cytokines in human macrophages by influenza A (H5N1) viruses: A mechanism for the unusual severity of human disease? Lancet 2002, 360, 1831–1837. [Google Scholar] [CrossRef]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the eye of the cytokine storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef]
- Da, S.E.; Jamur, M.C.; Oliver, C. Mast cell function: A new vision of an old cell. J. Histochem. Cytochem. 2014, 62, 698–738. [Google Scholar]
- Marshall, J.S.; Portales-Cervantes, L.; Leong, E. Mast Cell Responses to Viruses and Pathogen Products. Int. J. Mol. Sci. 2019, 20, 4241. [Google Scholar] [CrossRef]
- Boyce, J.A. Mast cells and eicosanoid mediators: A system of reciprocal paracrine and autocrine regulation. Immunol. Rev. 2007, 217, 168–185. [Google Scholar] [CrossRef]
- King, C.A.; McCurdy, J.D.; Marshall, J.S. Mast Cell Cytokine and Chemokine Responses to Bacterial and Viral Infection. Curr. Pharm. Des. 2003, 9, 11–24. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. Mast cells: Versatile regulators of inflammation, tissue remodeling, host defense and homeostasis. J. Dermatol. Sci. 2008, 49, 7–19. [Google Scholar] [CrossRef]
- Meng, D.; Huo, C.; Wang, M.; Xiao, J.; Liu, B.; Wei, T.; Dong, H.; Zhang, G.; Hu, Y.; Sun, L. Influenza A Viruses Replicate Productively in Mouse Mastocytoma Cells (P815) and Trigger Pro-inflammatory Cytokine and Chemokine Production through TLR3 Signaling Pathway. Front. Microbiol. 2016, 7, 2130. [Google Scholar] [CrossRef]
- Cai, R.; Tang, G.; Zhang, Q.; Yong, W.; Zhang, W.; Xiao, J.; Wei, C.; He, C.; Yang, G.; Pang, W. A Novel lnc-RNA, Named lnc-ORA, Is Identified by RNA-Seq Analysis, and Its Knockdown Inhibits Adipogenesis by Regulating the PI3K/AKT/mTOR Signaling Pathway. Cells 2019, 8, 477. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Nanduri, B.; Tan, W.; Cheng, B.; Fan, R.; Pruett, S.B. Sodium methyldithiocarbamate exerts broad inhibition of cellular signaling and expression of effector molecules of inflammation. Toxicol. Sci. 2013, 136, 430–442. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, Y.; Li, Y.; Burt, D.W.; Chen, H.; Zhang, Y.; Qian, W.; Kim, H.; Gan, S.; Zhao, Y.; Li, J.; et al. The duck genome and transcriptome provide insight into an avian influenza virus reservoir species. Nat. Genet. 2013, 45, 776–783. [Google Scholar] [CrossRef]
- Manchanda, H.; Seidel, N.; Blaess, M.; Claus, R.A.; Linde, J.; Slevogt, H.; Sauerbrei, A.; Guthke, R.; Schmidtke, M. Differential Biphasic Transcriptional Host Response Associated with Coevolution of Hemagglutinin Quasispecies of Influenza A Virus. Front. Microbiol. 2016, 7, 1167. [Google Scholar] [CrossRef] [PubMed]
- Forero, A.; Tisoncik-Go, J.; Watanabe, T.; Zhong, G.; Hatta, M.; Tchitchek, N.; Selinger, C.; Chang, J.; Barker, K.; Morrison, J.; et al. The 1918 Influenza Virus PB2 Protein Enhances Virulence through the Disruption of Inflammatory and Wnt-Mediated Signaling in Mice. J. Virol. 2016, 90, 2240–2253. [Google Scholar] [CrossRef]
- Cypryk, W.; Lorey, M.; Puustinen, A.; Nyman, T.A.; Matikainen, S. Proteomic and Bioinformatic Characterization of Extracellular Vesicles Released from Human Macrophages upon Influenza A Virus Infection. J. Proteome Res. 2017, 16, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Vijayakumar, P.; Gandhale, P.N.; Ranaware, P.B.; Kumar, H.; Kulkarni, D.D.; Raut, A.A.; Mishra, A. Genome-wide gene expression pattern underlying differential host response to high or low pathogenic H5N1 avian influenza virus in ducks. Acta Virol. 2017, 61, 66–76. [Google Scholar] [CrossRef]
- Yu, G.; Liang, W.; Liu, J.; Meng, D.; Wei, L.; Chai, T.; Cai, Y. Proteomic Analysis of Differential Expression of Cellular Proteins in Response to Avian H9N2 Virus Infection of A549 Cells. Front. Microbiol. 2016, 7, 1962. [Google Scholar] [CrossRef]
- Huo, C.; Jin, Y.; Zou, S.; Qi, P.; Xiao, J.; Tian, H.; Wang, M.; Hu, Y. Lethal influenza A virus preferentially activates TLR3 and triggers a severe inflammatory response. Virus Res. 2018, 257, 102–112. [Google Scholar] [CrossRef]
- Huo, C.; Xiao, K.; Zhang, S.; Tang, Y.; Wang, M.; Qi, P.; Xiao, J.; Tian, H.; Hu, Y. H5N1 Influenza a Virus Replicates Productively in Pancreatic Cells and Induces Apoptosis and Pro-Inflammatory Cytokine Response. Front. Cell. Infect. Microbiol. 2018, 8, 386. [Google Scholar] [CrossRef]
- Liu, B.; Meng, D.; Wei, T.; Zhang, S.; Hu, Y.; Wang, M. Apoptosis and Pro-inflammatory Cytokine Response of Mast Cells Induced by Influenza A Viruses. PLoS ONE 2014, 9, e100109. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jin, Y.; Han, D.; Zhang, G.; Cao, S.; Xie, J.; Xue, J.; Li, Y.; Meng, D.; Fan, X.; et al. Mast Cell-Induced Lung Injury in Mice Infected with H5N1 Influenza Virus. J. Virol. 2012, 86, 3347–3356. [Google Scholar] [CrossRef] [PubMed]
- Gunay-Aygun, M.; Falik-Zaccai, T.C.; Vilboux, T.; Zivony-Elboum, Y.; Gumruk, F.; Cetin, M.; Khayat, M.; Boerkoel, C.F.; Kfir, N.; Huang, Y.; et al. NBEAL2 is mutated in gray platelet syndrome and is required for biogenesis of platelet α-granules. Nat. Genet. 2011, 43, 732–734. [Google Scholar] [CrossRef]
- Mutz, K.-O.; Heilkenbrinker, A.; Lönne, M.; Walter, J.-G.; Stahl, F. Transcriptome analysis using next-generation sequencing. Curr. Opin. Biotechnol. 2013, 24, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhou, X.; Li, R.; Michal, J.J.; Zhang, S.; Dodson, M.V.; Zhang, Z.; Harland, R.M. Whole transcriptome analysis with sequencing: Methods, challenges and potential solutions. Cell. Mol. Life Sci. 2015, 72, 3425–3439. [Google Scholar] [CrossRef]
- Huo, C.; Wu, H.; Xiao, J.; Meng, D.; Zou, S.; Wang, M.; Qi, P.; Tian, H.; Hu, Y. Genomic and Bioinformatic Characterization of Mouse Mast Cells (P815) Upon Different Influenza A Virus (H1N1, H5N1, and H7N2) Infections. Front. Genet. 2019, 10, 595. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, R.; Fan, X.; Lian, Z.; Hu, Y. FoxOs could play an important role during influenza A viruses infection via microarray analysis based on GEO database. Infect. Genet. Evol. 2019, 75, 104009. [Google Scholar] [CrossRef] [PubMed]
- Schmolke, M.; Viemann, D.; Roth, J.; Ludwig, S. Essential impact of NF-kappaB signaling on the H5N1 influenza A virus-induced transcriptome. J. Immunol. 2009, 183, 5180–5189. [Google Scholar] [CrossRef]
- Albers, C.A.; Cvejic, A.; Favier, R.; Bouwmans, E.E.; Alessi, M.C.; Bertone, P.; Jordan, G.; Kettleborough, R.N.; Kiddle, G.; Kostadima, M.; et al. Exome sequencing identifies NBEAL2 as the causative gene for gray platelet syndrome. Nat. Genet. 2011, 43, 735–737. [Google Scholar] [CrossRef]
- Kahr, W.H.; Hinckley, J.; Li, L.; Schwertz, H.; Christensen, H.; Rowley, J.W.; Pluthero, F.; Urban, D.; Fabbro, S.; Nixon, B.; et al. Mutations in NBEAL2, encoding a BEACH protein, cause gray platelet syndrome. Nat. Genet. 2011, 43, 738–740. [Google Scholar] [CrossRef]
- Claushuis, T.; de Stoppelaar, S.F.; de Vos, A.F.; Grootemaat, A.E.; van der Wel, N.N.; Roelofs, J.; Ware, J.; van’t Veer, C.; van der Poll, T. Nbeal2 Deficiency Increases Organ Damage but Does Not Affect Host Defense During Gram-Negative Pneumonia-Derived Sepsis. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1772–1784. [Google Scholar] [CrossRef] [PubMed]
- Sowerby, J.M.; Thomas, D.C.; Clare, S.; Espeli, M.; Guerrero, J.A.; Hoenderdos, K.; Harcourt, K.; Marsden, M.; Abdul-Karim, J.; Clement, M.; et al. NBEAL2 is required for neutrophil and NK cell function and pathogen defense. J. Clin. Investig. 2017, 127, 3521–3526. [Google Scholar] [CrossRef] [PubMed]
- Drube, S.; Grimlowski, R.; Deppermann, C.; Fröbel, J.; Kraft, F.; Andreas, N.; Stegner, D.; Dudeck, J.; Weber, F.; Rödiger, M.; et al. The Neurobeachin-like 2 Protein Regulates Mast Cell Homeostasis. J. Immunol. 2017, 199, 2948–2957. [Google Scholar] [CrossRef] [PubMed]
- Zarnegar, B.; Westin, A.; Evangelidou, S.; Hallgren, J. Innate Immunity Induces the Accumulation of Lung Mast Cells During Influenza Infection. Front. Immunol. 2018, 9, 2288. [Google Scholar] [CrossRef]
- Lee, I.H.; Kim, H.S.; Seo, S.H. Porcine mast cells infected with H1N1 influenza virus release histamine and inflammatory cytokines and chemokines. Arch. Virol. 2017, 162, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Dawicki, W.; Marshall, J.S. New and emerging roles for mast cells in host defense. Curr. Opin. Immunol. 2007, 19, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Deppermann, C.; Cherpokova, D.; Nurden, P.; Schulz, J.-N.; Thielmann, I.; Kraft, P.; Vögtle, T.; Kleinschnitz, C.; Dütting, S.; Krohne, G.; et al. Gray platelet syndrome and defective thrombo-inflammation in Nbeal2-deficient mice. J. Clin. Investig. 2013, 123, 3331–3342. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences |
|---|---|
| Nbeal2-Mus-1177 | F: 5′-GCUACAAGCCACCUUCCUUTT-3′ |
| R: 5′-AAGGAAGGUGGCUUGUAGCTT-3′ | |
| Nbeal2-Mus-3274 | F: 5′-GCACCUGCUCUUCAACUUUTT-3′ |
| R: 5′-AAAGUUGAAGAGCAGGUGCTT-3′ | |
| Nbeal2-Mus-8095 | F: 5′-GCACCUGUAUUCAGUGAAUTT-3′ |
| R: 5′-AUUCACUGAAUACAGGUGCTT-3′ | |
| negative control | F: 5′-UUCUCCGAACGUGUCACGUTT -3′ |
| R: 5′-ACGUGACACGUUCGGAGAATT -3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Wu, H.; Huo, C.; Zou, S.; Hu, Y.; Yang, H. Transcriptomic Profiling of Mouse Mast Cells upon Pathogenic Avian H5N1 and Pandemic H1N1 Influenza a Virus Infection. Viruses 2022, 14, 292. https://doi.org/10.3390/v14020292
Tang Y, Wu H, Huo C, Zou S, Hu Y, Yang H. Transcriptomic Profiling of Mouse Mast Cells upon Pathogenic Avian H5N1 and Pandemic H1N1 Influenza a Virus Infection. Viruses. 2022; 14(2):292. https://doi.org/10.3390/v14020292
Chicago/Turabian StyleTang, Yuling, Hongping Wu, Caiyun Huo, Shumei Zou, Yanxin Hu, and Hanchun Yang. 2022. "Transcriptomic Profiling of Mouse Mast Cells upon Pathogenic Avian H5N1 and Pandemic H1N1 Influenza a Virus Infection" Viruses 14, no. 2: 292. https://doi.org/10.3390/v14020292
APA StyleTang, Y., Wu, H., Huo, C., Zou, S., Hu, Y., & Yang, H. (2022). Transcriptomic Profiling of Mouse Mast Cells upon Pathogenic Avian H5N1 and Pandemic H1N1 Influenza a Virus Infection. Viruses, 14(2), 292. https://doi.org/10.3390/v14020292