


Downregulation of the Protein C Signaling System Is Associated with COVID-19 Hypercoagulability—A Single-Cell Transcriptomics Analysis
 ,
, 

Abstract
1. Introduction
2. Methods
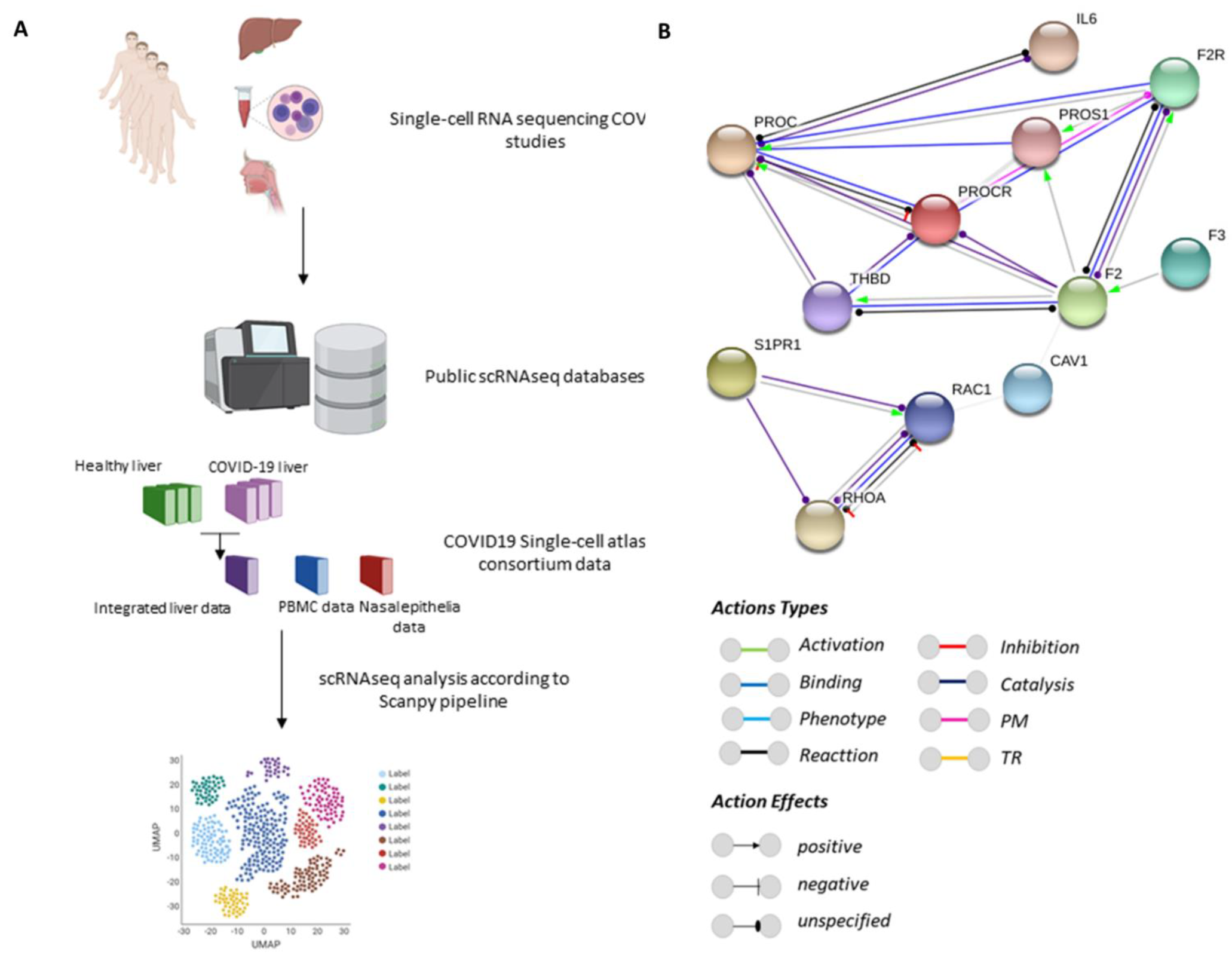
2.1. Protein–Protein Interaction Networks
2.2. Analysis of Single-Cell RNA-Sequencing (scRNA-seq) Data
2.3. Statistical Analyses
2.4. Data Availability
2.5. Code Availability
2.6. Generation of Diagrams and Illustrations
3. Results
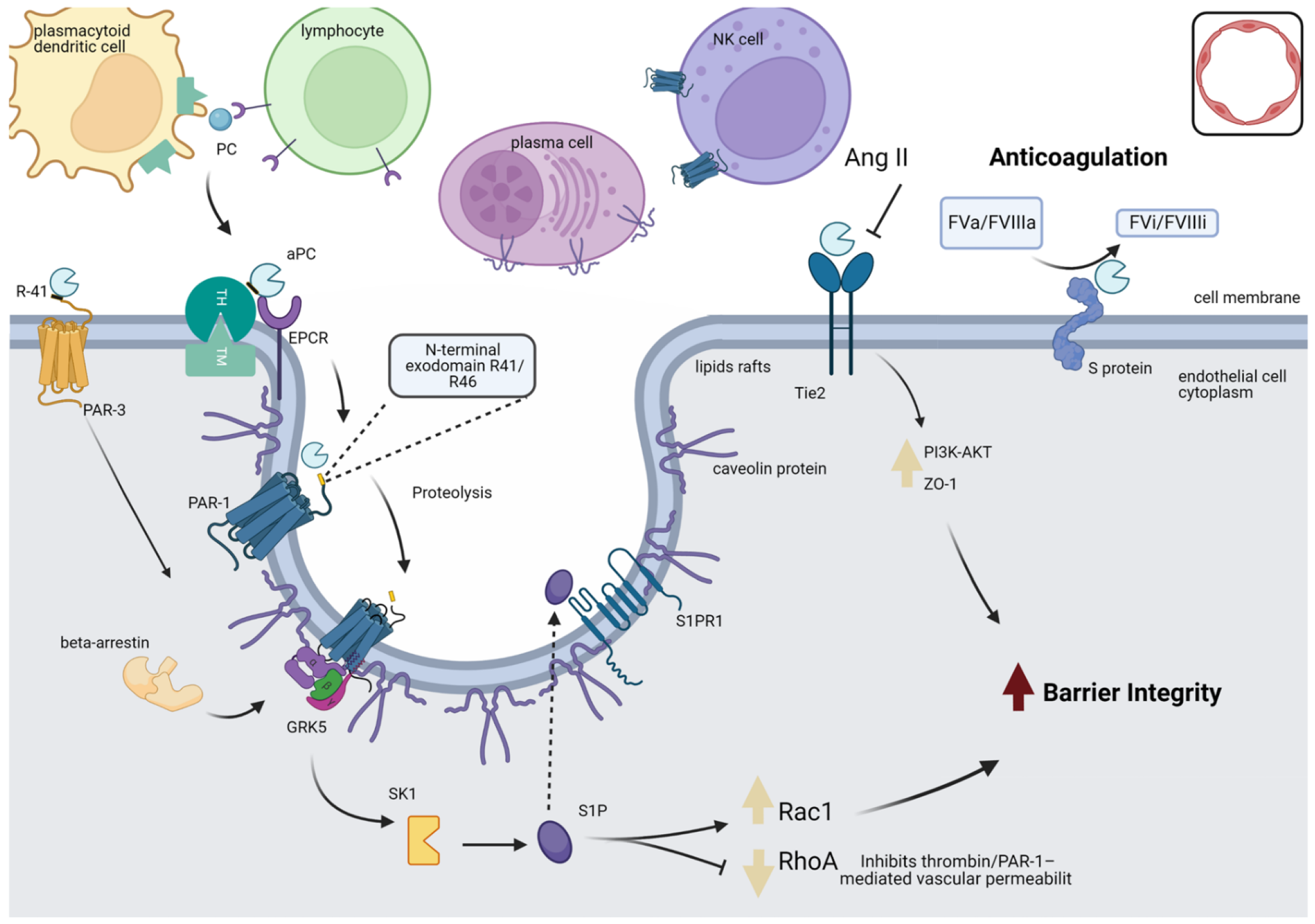
3.1. PROC Interacts with Proteins Belonging to Barrier Integrity and the Inflammatory and Coagulation Systems
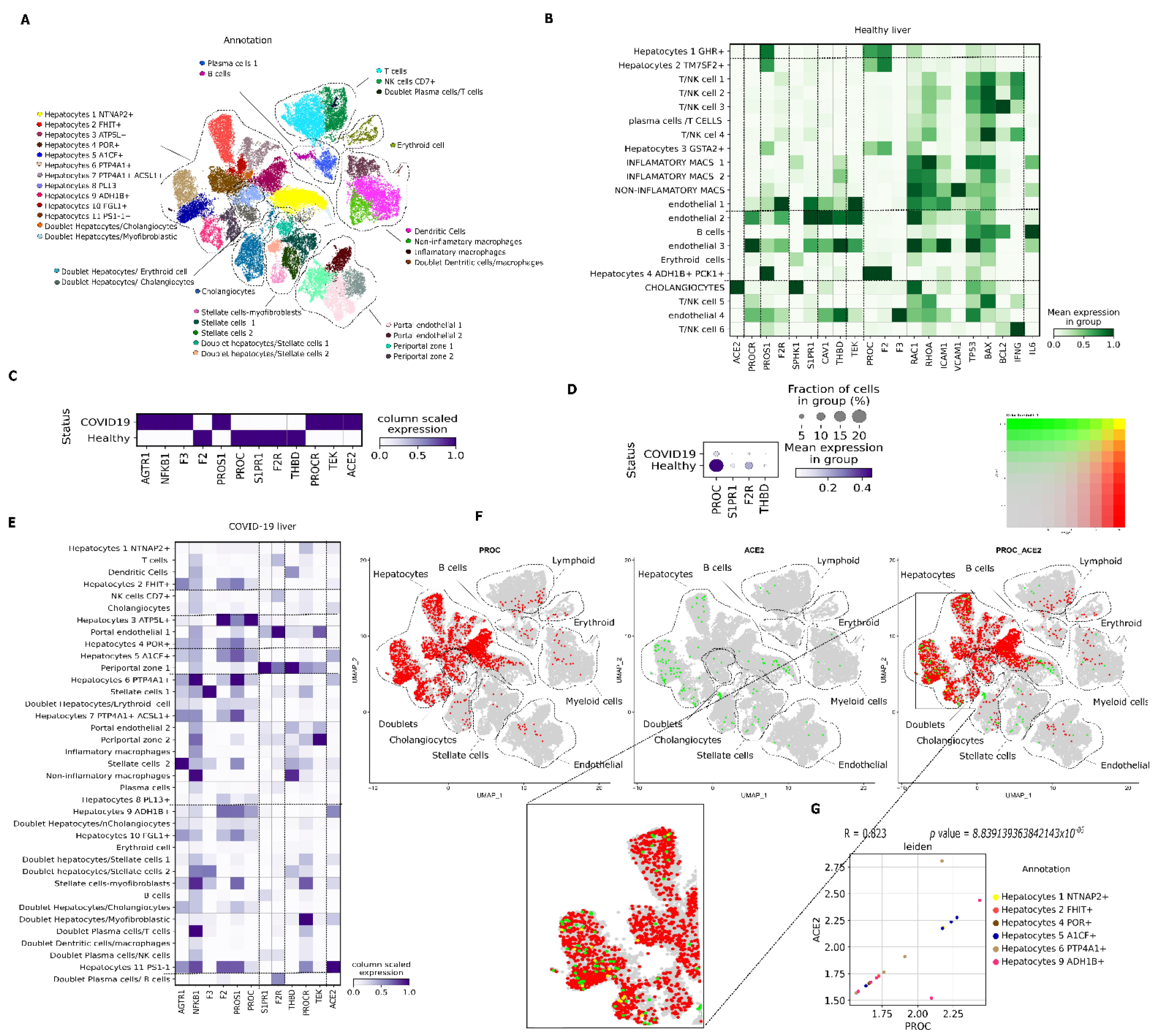
3.2. PC pathway Landscape at the Single-Cell Level in Tissues from Patients with COVID-19
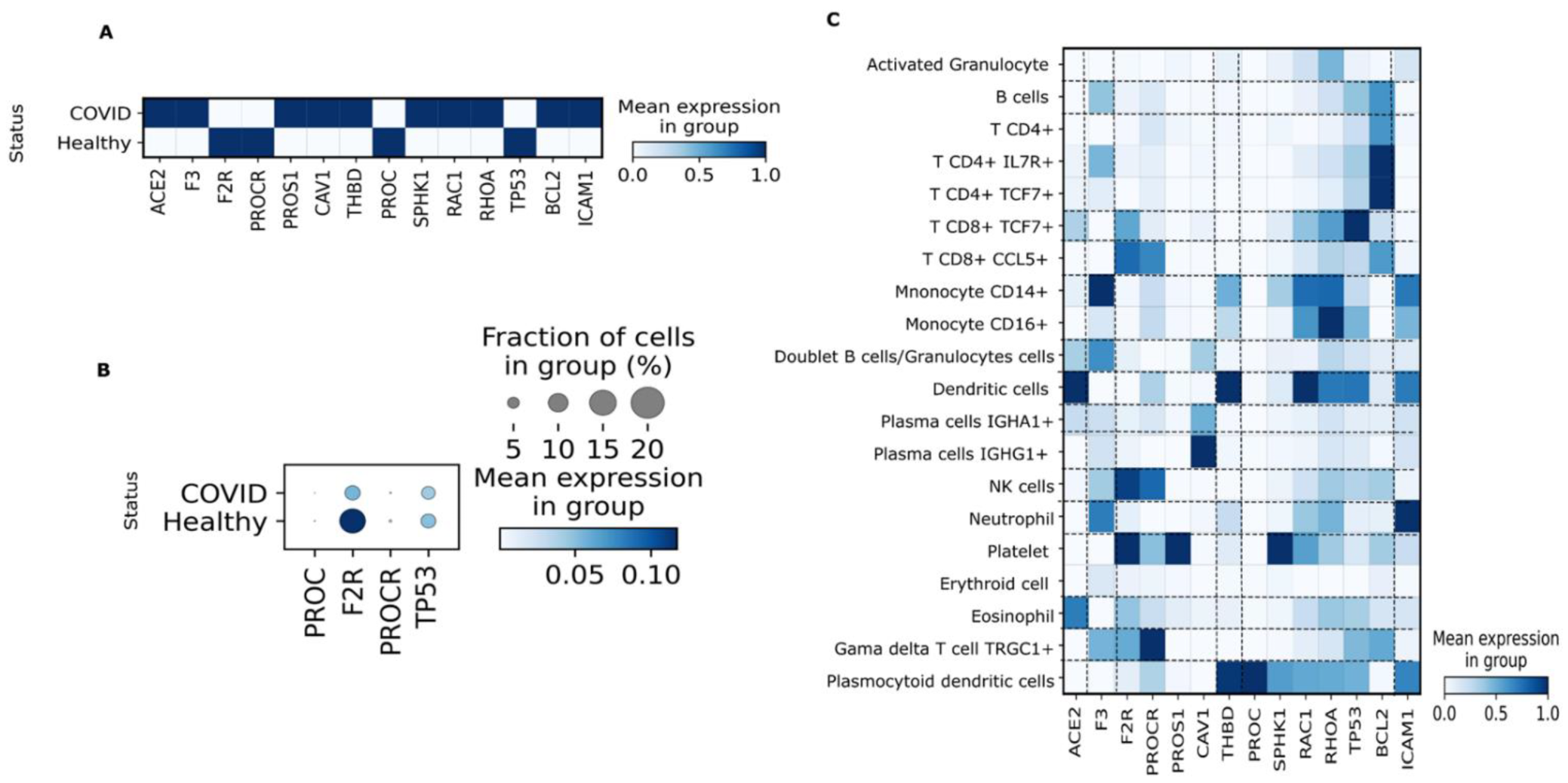
3.3. PBMC
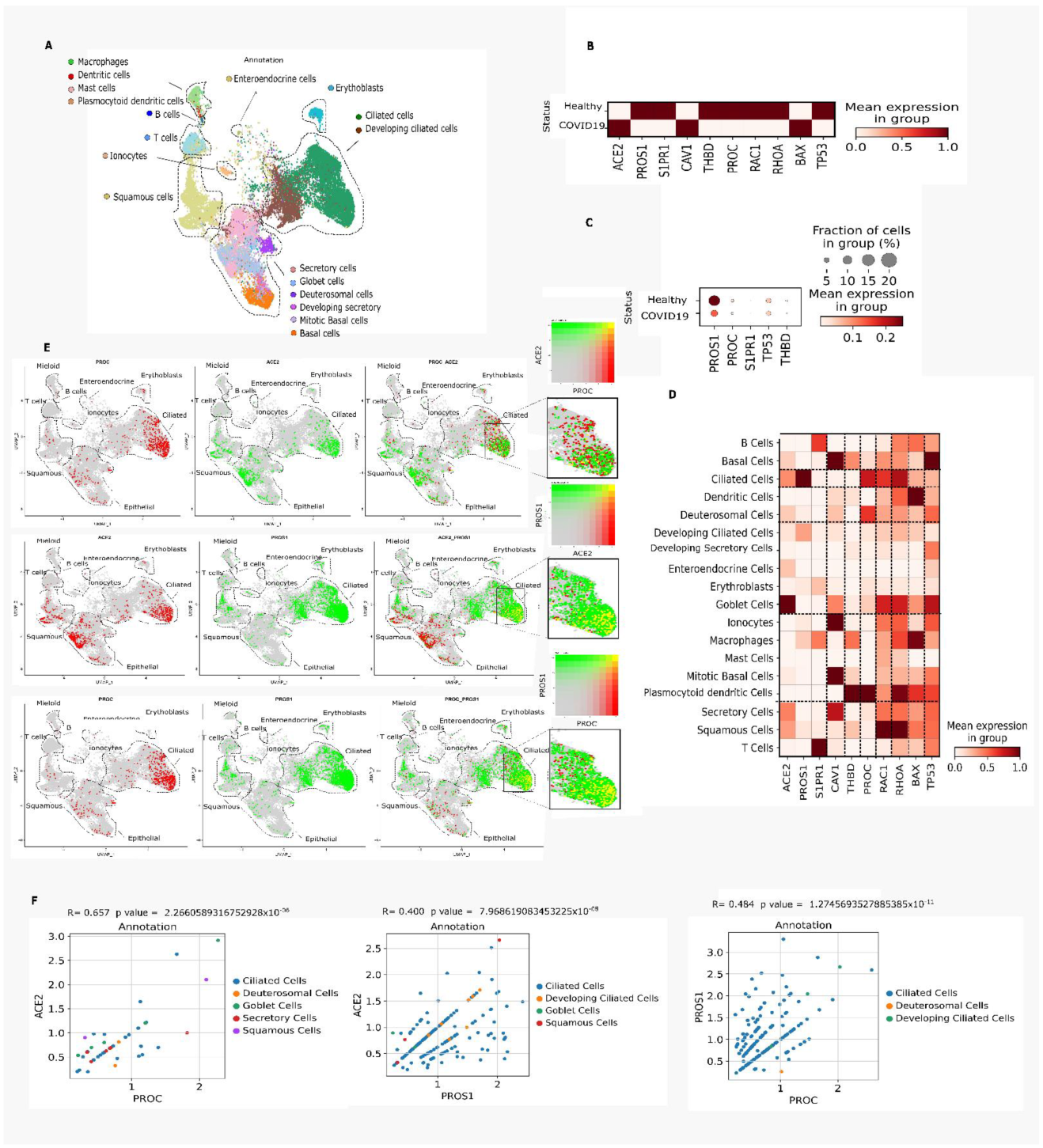
3.4. Nasal Epithelium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iba, T.; Warkentin, T.E.; Thachil, J.; Levi, M.; Levy, J.H. Proposal of the definition for COVID-19-associated coagulopathy. J. Clin. Med. 2021, 10, 191. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.; Wu, A.; Cheng, V.; Tang, B.; Chan, C.; Yung, C.; Luk, S.; Lee, T.; Chow, L.; Yuen, K. Pulmonary artery thrombosis in a patient with severe acute respiratory syndrome. Postgrad. Med. J. 2005, 81, e3. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Dolhnikoff, M.; Duarte-Neto, A.N.; de Almeida Monteiro, R.A.; da Silva, L.F.F.; de Oliveira, E.P.; Saldiva, P.H.N.; Mauad, T.; Negri, E.M. Pathological evidence of pulmonary thrombotic phenomena in severe COVID-19. J. Thromb. Haemost. 2020, 18, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- Pestka, S.B. Old drug, new Trick? The rationale for the treatment of COVID-19 with activated protein C. Med. Hypotheses 2021, 149, 110537. [Google Scholar] [CrossRef]
- Gupta, A.; Madhavan, M.V.; Sehgal, K.; Nair, N.; Mahajan, S.; Sehrawat, T.S.; Bikdeli, B.; Ahluwalia, N.; Ausiello, J.C.; Wan, E.Y.; et al. Extrapulmonary manifestations of COVID-19. Nat. Med. 2020, 26, 1017–1032. [Google Scholar] [CrossRef]
- Bonaventura, A.; Vecchie, A.; Dagna, L.; Martinod, K.; Dixon, D.L.; Van Tassell, B.W.; Dentali, F.; Montecucco, F.; Massberg, S.; Levi, M.; et al. Endothelial dysfunction and immunothrombosis as key pathogenic mechanisms in COVID-19. Nat. Rev. Immunol. 2021, 21, 319–329. [Google Scholar] [CrossRef]
- Jin, Y.; Ji, W.; Yang, H.; Chen, S.; Zhang, W.; Duan, G. Endothelial activation and dysfunction in COVID-19: From basic mechanisms to potential therapeutic approaches. Signal Transduct. Target. 2020, 5, 293. [Google Scholar] [CrossRef]
- Pan, H.; Peto, R.; Karim, Q. WHO, Solidarity trial consortium. As the members of the writing committee. Repurposed antiviral drugs for COVID-19–interim WHO SOLIDARITY trial results. medRxiv 2020, 10, 15.20209817. [Google Scholar]
- Gharbharan, A.; Jordans, C.C.; GeurtsvanKessel, C.; den Hollander, J.G.; Karim, F.; Mollema, F.P.; Stalenhoef, J.E.; Dofferhoff, A.; Ludwig, I.; Koster, A. Convalescent plasma for COVID-19. A randomized clinical trial. medRxiv 2020. [Google Scholar] [CrossRef]
- Wang, D.; Fu, B.; Peng, Z.; Yang, D.; Han, M.; Li, M.; Yang, Y.; Yang, T.; Sun, L.; Li, W. Tocilizumab in patients with moderate or severe COVID-19: A randomized, controlled, open-label, multicenter trial. Front. Med. 2021, 15, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.D.; Millar, J.E.; Baillie, J.K. Clinical evidence does not support corticosteroid treatment for 2019-nCoV lung injury. Lancet 2020, 395, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Mansour, E.; Palma, A.C.; Ulaf, R.G.; Ribeiro, L.C.; Bernardes, A.F.; Nunes, T.A.; Agrela, M.V.; Bombassaro, B.; Monfort-Pires, M.; Camargo, R.L. Safety and outcomes associated with the pharmacological inhibition of the kinin–kallikrein system in severe COVID-19. Viruses 2021, 13, 309. [Google Scholar] [CrossRef] [PubMed]
- Suthar, A.B.; Wang, J.; Seffren, V.; Wiegand, R.E.; Griffing, S.; Zell, E. Public health impact of covid-19 vaccines in the US: Observational study. BMJ 2022, 377, e069317. [Google Scholar] [CrossRef] [PubMed]
- Fiolet, T.; Kherabi, Y.; MacDonald, C.-J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: A narrative review. Clin. Microbiol. Infect. 2021, 12, 202–221. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- ATTACC, A.-a.; Investigators, R.-C. Therapeutic anticoagulation with heparin in noncritically ill patients with Covid-19. N. Engl. J. Med. 2021, 385, 790–802. [Google Scholar]
- Willyard, C. Coronavirus blood-clot mystery intensifies. Nature 2020, 581, 250. [Google Scholar] [CrossRef]
- Esmon, C.; Stenflo, J.; Suttie, J. A new vitamin K-dependent protein. A phospholipid-binding zymogen of a serine esterase. J. Biol. Chem. 1976, 251, 3052–3056. [Google Scholar] [CrossRef]
- He, X.; Shen, L.; Bjartell, A.; Malm, J.; Lilja, H.; Dahlbäck, B. The gene encoding vitamin K-dependent anticoagulant protein C is expressed in human male reproductive tissues. J. Histochem. Cytochem. 1995, 43, 563–570. [Google Scholar] [CrossRef]
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.-K.; Amini, B.; Andersen, E.; Andersson, A.-C.; Angelidou, P.; Asplund, A.; Asplund, C. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef] [PubMed]
- Van Cott, K.E.; Williams, B.; Velander, W.H.; Gwazdauskas, F.; Lee, T.; Lubon, H.; Drohan, W.N. Affinity purification of biologically active and inactive forms of recombinant human protein C produced in porcine mammary gland. J. Mol. Recognit. 1996, 9, 407–414. [Google Scholar] [CrossRef]
- Riewald, M.; Petrovan, R.J.; Donner, A.; Ruf, W. Activated protein C signals through the thrombin receptor PAR1 in endothelial cells. J. Endotoxin Res. 2003, 9, 317–321. [Google Scholar] [CrossRef]
- Stearns-Kurosawa, D.J.; Kurosawa, S.; Mollica, J.S.; Ferrell, G.L.; Esmon, C.T. The endothelial cell protein C receptor augments protein C activation by the thrombin-thrombomodulin complex. Proc. Natl. Acad. Sci. USA 1996, 93, 10212–10216. [Google Scholar] [CrossRef]
- Schuepbach, R.A.; Feistritzer, C.; Brass, L.F.; Riewald, M. Activated protein C-cleaved protease activated receptor-1 is retained on the endothelial cell surface even in the presence of thrombin. Blood 2008, 111, 2667–2673. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.A.; Lay, F.T.; Poon, I.K.; Kvansakul, M.; Hulett, M.D. Tumor cell membrane-targeting cationic antimicrobial peptides: Novel insights into mechanisms of action and therapeutic prospects. Cell. Mol. Life Sci. 2017, 74, 3809–3825. [Google Scholar] [CrossRef]
- Whetstone, W.D.; Walker, B.; Trivedi, A.; Lee, S.; Noble-Haeusslein, L.J.; Hsu, J.C. Protease-Activated Receptor-1 Supports Locomotor Recovery by Biased Agonist Activated Protein C after Contusive Spinal Cord Injury. PLoS ONE 2017, 12, e0170512. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Delorey, T.M.; Ziegler, C.G.; Heimberg, G.; Normand, R.; Yang, Y.; Segerstolpe, Å.; Abbondanza, D.; Fleming, S.J.; Subramanian, A.; Montoro, D.T. COVID-19 tissue atlases reveal SARS-CoV-2 pathology and cellular targets. Nature 2021, 595, 107–113. [Google Scholar] [CrossRef]
- MacParland, S.A.; Liu, J.C.; Ma, X.Z.; Innes, B.T.; Bartczak, A.M.; Gage, B.K.; Manuel, J.; Khuu, N.; Echeverri, J.; Linares, I.; et al. Single cell RNA sequencing of human liver reveals distinct intrahepatic macrophage populations. Nat. Commun. 2018, 9, 4383. [Google Scholar] [CrossRef]
- Ziegler, C.G.; Miao, V.N.; Owings, A.H.; Navia, A.W.; Tang, Y.; Bromley, J.D.; Lotfy, P.; Sloan, M.; Laird, H.; Williams, H.B. Impaired local intrinsic immunity to SARS-CoV-2 infection in severe COVID-19. Cell 2021, 184, 4713–4733.e4722. [Google Scholar] [CrossRef]
- COVID CZIS-C; Ballestar, E.; Farber, D.L.; Glover, S.; Horwitz, B.; Meyer, K.; Nikolić, M.; Ordovas-Montanes, J.; Sims, P.; Shalek, A. Single cell profiling of COVID-19 patients: An international data resource from multiple tissues. MedRxiv 2020. [Google Scholar] [CrossRef]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martínez-Colón, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T. A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef]
- Wolf, F.A.; Angerer, P.; Theis, F.J. SCANPY: Large-scale single-cell gene expression data analysis. Genome Biol. 2018, 19, 15. [Google Scholar] [CrossRef]
- Luecken, M.D.; Theis, F.J. Current best practices in single-cell RNA-seq analysis: A tutorial. Mol. Syst. Biol. 2019, 15, e8746. [Google Scholar] [CrossRef]
- Traag, V.A.; Waltman, L.; Van Eck, N.J. From Louvain to Leiden: Guaranteeing well-connected communities. Sci. Rep. 2019, 9, 5233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Garmire, L.X. Data Analysis in Single-Cell RNA-Seq. In Single-Cell Omics; Debmalya Barh, V.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 419–432. [Google Scholar]
- White, B.; Schmidt, M.; Murphy, C.; Livingstone, W.; O’toole, D.; Lawler, M.; O’Neill, L.; Kelleher, D.; Schwarz, H.; Smith, O. Activated protein C inhibits lipopolysaccharide-induced nuclear translocation of nuclear factor κB (NF-κB) and tumour necrosis factor α (TNF-α) production in the THP-1 monocytic cell line. Br. J. Haematol. 2000, 110, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Sampaziotis, F. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Della Valle, P.; Pavani, G.; D’Angelo, A. The protein C pathway and sepsis. Thromb. Res. 2012, 129, 296–300. [Google Scholar] [CrossRef]
- Gu, X.; Zhou, F.; Wang, Y.; Fan, G.; Cao, B. Respiratory viral sepsis: Epidemiology, pathophysiology, diagnosis and treatment. Eur. Respir. Rev. 2020, 29, 200038. [Google Scholar] [CrossRef] [PubMed]
- Phua, J.; Ngerng, W.J.; See, K.C.; Tay, C.K.; Kiong, T.; Lim, H.F.; Chew, M.Y.; Yip, H.S.; Tan, A.; Khalizah, H.J. Characteristics and outcomes of culture-negative versus culture-positive severe sepsis. Crit. Care 2013, 17, R202. [Google Scholar] [CrossRef] [PubMed]
- Ljungström, L.R.; Jacobsson, G.; Claesson, B.E.; Andersson, R.; Enroth, H. Respiratory viral infections are underdiagnosed in patients with suspected sepsis. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1767–1776. [Google Scholar] [CrossRef]
- Bakalli, I. Liver dysfunction in severe sepsis from respiratory syncytial virus. J. Pediatr. Intensive Care 2018, 7, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Farcas, G.A.; Poutanen, S.M.; Mazzulli, T.; Willey, B.M.; Butany, J.; Asa, S.L.; Faure, P.; Akhavan, P.; Low, D.E.; Kain, K.C. Fatal severe acute respiratory syndrome is associated with multiorgan involvement by coronavirus. J. Infect. Dis. 2005, 191, 193–197. [Google Scholar] [CrossRef]
- Nin, N.; Lorente, J.; Soto, L.; Ríos, F.; Hurtado, J.; Arancibia, F.; Ugarte, S.; Echevarría, E.; Cardinal, P.; Saldarini, F. Acute kidney injury in critically ill patients with 2009 influenza A (H1N1) viral pneumonia: An observational study. Intensive Care Med. 2011, 37, 768–774. [Google Scholar] [CrossRef]
- Gao, C.; Wang, Y.; Gu, X.; Shen, X.; Zhou, D.; Zhou, S.; Huang, J.-a.; Cao, B.; Guo, Q. Association between cardiac injury and mortality in hospitalized patients infected with avian influenza A (H7N9) virus. Crit. Care Med. 2020, 48, 451. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; Yu, T. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H. Sepsis-induced coagulopathy and disseminated intravascular coagulation. Anesthesiology 2020, 132, 1238–1245. [Google Scholar] [CrossRef]
- Manson, J.J.; Crooks, C.; Naja, M.; Ledlie, A.; Goulden, B.; Liddle, T.; Khan, E.; Mehta, P.; Martin-Gutierrez, L.; Waddington, K.E. COVID-19-associated hyperinflammation and escalation of patient care: A retrospective longitudinal cohort study. Lancet Rheumatol. 2020, 2, e594–e602. [Google Scholar] [CrossRef]
- Samavati, L.; Uhal, B.D. ACE2, much more than just a receptor for SARS-COV-2. Front. Cell. Infect. Microbiol. 2020, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, J.; Montezano, A.C.; Touyz, R.M. ACE2/Ang-(1-7)/Mas1 axis and the vascular system: Vasoprotection to COVID-19-associated vascular disease. Clin. Sci. 2021, 135, 387–407. [Google Scholar] [CrossRef] [PubMed]
- Meini, S.; Zanichelli, A.; Sbrojavacca, R.; Iuri, F.; Roberts, A.T.; Suffritti, C.; Tascini, C. Understanding the pathophysiology of COVID-19: Could the contact system be the key? Front. Immunol. 2020, 11, 2014. [Google Scholar] [CrossRef] [PubMed]
- Gando, S.; Wada, T. Thromboplasminflammation in COVID-19 coagulopathy: Three viewpoints for diagnostic and therapeutic strategies. Front. Immunol. 2021, 12, 649122. [Google Scholar] [CrossRef]
- Li, G.; He, X.; Zhang, L.; Ran, Q.; Wang, J.; Xiong, A.; Wu, D.; Chen, F.; Sun, J.; Chang, C. Assessing ACE2 expression patterns in lung tissues in the pathogenesis of COVID-19. J. Autoimmun. 2020, 112, 102463. [Google Scholar] [CrossRef]
- Capettini, L.S.A.; Montecucco, F.; Mach, F.; Stergiopulos, N.; Santos, R.A.S.; da Silva, R.F. Role of renin-angiotensin system in inflammation, immunity and aging. Curr. Pharm. Des. 2012, 18, 963–970. [Google Scholar] [CrossRef]
- Rautou, P.-E.; Tatsumi, K.; Antoniak, S.; Owens III, A.P.; Sparkenbaugh, E.; Holle, L.A.; Wolberg, A.S.; Kopec, A.K.; Pawlinski, R.; Luyendyk, J.P. Hepatocyte tissue factor contributes to the hypercoagulable state in a mouse model of chronic liver injury. J. Hepatol. 2016, 64, 53–59. [Google Scholar] [CrossRef]
- Shim, K.Y.; Eom, Y.W.; Kim, M.Y.; Kang, S.H.; Baik, S.K. Role of the renin-angiotensin system in hepatic fibrosis and portal hypertension. Korean J. Intern. Med. 2018, 33, 453. [Google Scholar] [CrossRef]
- Elsharkawy, A.M.; Mann, D.A. Nuclear factor-κB and the hepatic inflammation-fibrosis-cancer axis. Hepatology 2007, 46, 590–597. [Google Scholar] [CrossRef]
- Rehan, M.; Ahmed, F.; Howladar, S.M.; Refai, M.Y.; Baeissa, H.M.; Zughaibi, T.A.; Kedwa, K.M.; Jamal, M.S. A Computational Approach Identified Andrographolide as a Potential Drug for Suppressing COVID-19-Induced Cytokine Storm. Front. Immunol. 2021, 12, 648250. [Google Scholar] [CrossRef]
- Okajima, K. Prevention of Endothelial Cell Injury by Activated Protein C: The Molecular Mechanism(s) and Therapeutic Implications. Curr. Vasc. Pharmacol. 2004, 2, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, T.D.; Cordioli, R.L.; Campos Guerra, J.C.; Caldin da Silva, B.; dos Reis Rodrigues, R.; De Souza, G.M.; Midega, T.D.; Campos, N.S.; Carneiro, B.V.; Campos, F.N.D. Coagulation profile of COVID-19 patients admitted to the ICU: An exploratory study. PLoS ONE 2020, 15, e0243604. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, E.S.; Jamieson, A.M. Unique transcriptional changes in coagulation cascade genes in SARS-CoV-2-infected lung 3 epithelial cells: A potential factor in COVID-19 coagulopathies 4. bioRxiv 2020. [Google Scholar] [CrossRef]
- Stanne, T.M.; Pedersén, A.; Gisslén, M.; Jern, C. Low admission protein C levels is a risk factor for disease worsening and mortality in hospitalized patients with COVID-19. Thromb. Res. 2021, 204, 13–15. [Google Scholar] [CrossRef]
- Panigada, M.; Bottino, N.; Tagliabue, P.; Grasselli, G.; Novembrino, C.; Chantarangkul, V.; Pesenti, A.; Peyvandi, F.; Tripodi, A. Hypercoagulability of COVID-19 patients in intensive care unit: A report of thromboelastography findings and other parameters of hemostasis. J. Thromb. Haemost. 2020, 18, 1738–1742. [Google Scholar] [CrossRef]
- Tabatabai, A.; Rabin, J.; Menaker, J.; Madathil, R.; Galvagno, S.; Menne, A.; Chow, J.H.; Grazioli, A.; Herr, D.; Tanaka, K. Factor VIII and functional protein C activity in critically ill patients with coronavirus disease 2019: A case series. Aa Pract. 2020, 14, e01236. [Google Scholar] [CrossRef]
- Ahn, J.H.; Kim, J.; Hong, S.P.; Choi, S.Y.; Yang, M.J.; Ju, Y.S.; Kim, Y.T.; Kim, H.M.; Rahman, M.T.; Chung, M.K. Nasal ciliated cells are primary targets for SARS-CoV-2 replication in early stage of COVID-19. J. Clin. Investig. 2021, 131, e148517. [Google Scholar] [CrossRef]
- Yuan, J.; Fan, D.; Xue, Z.; Qu, J.; Su, J. Co-expression of mitochondrial genes and ACE2 in cornea involved in COVID-19. Investig. Ophthalmol. Vis. Sci. 2020, 61, 13. [Google Scholar] [CrossRef]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Chen, L.; Li, J.; Wang, X.; Wang, F. The landscape of lung bronchoalveolar immune cells in COVID-19 revealed by single-cell RNA sequencing. MedRxiv 2020. [Google Scholar] [CrossRef]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L. Severe COVID-19 is marked by a dysregulated myeloid cell compartment. Cell 2020, 182, 1419–1440.e1423. [Google Scholar] [CrossRef]
- Willis Fox, O.; Preston, R.J. Molecular basis of protease-activated receptor 1 signaling diversity. J. Thromb. Haemost. 2020, 18, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Xiong, W.-F. Advances in the relationship between coronavirus infection and coagulation function. Chin. Med. J. 2020, 133, 1489. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Soh, U.J.; Paing, M.M.; Arora, P.; Trejo, J. Caveolae are required for protease-selective signaling by protease-activated receptor–1. Proc. Natl. Acad. Sci. USA 2009, 106, 6393–6397. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, J.; Assmann, J.; Schwaninger, M. Thrombomodulin–a new target for treating stroke at the crossroad of coagulation and inflammation. Curr. Med. Chem. 2014, 21, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Kishi, Y.; Kondo, T.; Xiao, S.; Yosef, N.; Gaublomme, J.; Wu, C.; Wang, C.; Chihara, N.; Regev, A.; Joller, N. Protein C receptor (PROCR) is a negative regulator of Th17 pathogenicity. J. Exp. Med. 2016, 213, 2489–2501. [Google Scholar] [CrossRef]
- Schoergenhofer, C.; Schwameis, M.; Gelbenegger, G.; Buchtele, N.; Thaler, B.; Mussbacher, M.; Schabbauer, G.; Wojta, J.; Jilma-Stohlawetz, P.; Jilma, B. Inhibition of protease-activated receptor (PAR1) reduces activation of the endothelium, coagulation, fibrinolysis and inflammation during human endotoxemia. Thromb. Haemost. 2018, 118, 1176–1184. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Liver | PBMC | Nasal Epithelium | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Protein Product | LogFC | COVID-19% | Healthy % | Adj. p-Value | LogFC | COVID-19% | Healthy % | Adj. p-Value | LogFC | COVID-19% | Healthy % | Adj. p-Value |
| ACE2 | Angiotensin-converting enzyme 2 | 2.868 | 0.434 | 0.068 | 1.54 × 10−14 | 2.943 | 0.046 | 0.006 | 0.013 | 0.429 | 3.336 | 2.583 | 0.000161 |
| AGTR1 | Angiotensin II receptor type 1 | 1.937 | 6.534 | 1.862 | 2.349 × 10−98 | - | - | - | - | - | - | - | - |
| BAX | BCL2-associated X protein | −4.150 | 0.958 | 14.268 | 2.285 × 10−168 | - | - | - | - | 0.211 | 6.778 | 5.587 | 0.009326 |
| BCL2 | BCL2 apoptosis regulator | 2.11 | 12.903 | 4.323 | 7.662 × 10−231 | 0.211 | 20.762 | 19.053 | 8.444 × 10−10 | - | - | - | - |
| CAV1 | Caveolin 1 | −27.18 | 0 | 6.134 | 2.662 × 10−80 | 2.685 | 0.889 | 0.144 | 3.332 × 10−31 | - | - | - | - |
| F2 | Prothrombin | −1.685 | 12.572 | 29.613 | 1.753 × 10−167 | - | - | - | - | - | - | - | |
| F2R | Coagulation factor II Thrombin receptor | −2.533 | 1.751 | 9.005 | 9.583 × 10−78 | −1.127 | 7.090 | 14.326 | 1.136 × 10−118 | - | - | - | - |
| F3 | Tissue factor | 1.227 | 0.722 | 0.324 | 3.121 × 10−06 | 1.309 | 0.249 | 0.102 | 0.0004 | - | - | - | - |
| ICAM1 | Intercellular adhesion molecule 1 | −0.284 | 3.527 | 4.716 | 0.906 | 3.637 | 1.948 | 9.690 × 10−26 | 0.548 | 7.868 | 4.905 | 0.000189 | |
| NFKB1 | Nuclear factor kappa B subunit 1 | 1.812 | 16.648 | 6.015 | 9.235 × 10−209 | 0.121 | 16.039 | 14.650 | 0.0024 | −0.261 | 13.716 | 16.128 | 5.76 × 10−05 |
| PROC | Protein C | −2.192 | 5.487 | 19.480 | 9.039 × 10−149 | −1.040 | 0.085 | 0.174 | 0.0462 | −0.502 | 1.659 | 2.432 | 0.000246 |
| PROCR | Endothelial protein C receptor | 0.447 | 1.931 | 2.376 | 0.0012 | −1.097 | 0.302 | 0.643 | 5.658 × 10−06 | - | - | - | - |
| PROS1 | Protein S | 30.593 | 18.412 | 0.16 | 1.731 × 10−08 | 1.026 | 0.398 | 0.198 | 0.00034 | −0.985 | 6.894 | 12.993 | 7.8006 × 10−34 |
| RAC1 | Rac family small GTPase 1 | −30.680 | 0 | 42.942 | 0 | 0.259 | 28.828 | 24.845 | 4.315 × 10−21 | −0.767 | 34.803 | 38.897 | 3.00078 × 10−33 |
| RHOA | Rho family of small GTPases, | −30.570 | 0 | 41.165 | 0 | 0.113 | 54.260 | 51.530 | 7.641 × 10−09 | −0.841 | 42.300 | 46.841 | 1.98516 × 10−36 |
| S1PR1 | Sphingosine-1-phosphate receptor 1 | −2.686 | 0.685 | 4.306 | 3.276 × 10−39 | - | - | - | - | −1.108 | 0.132 | 0.254 | 0.024404 |
| SPHK1 | Sphingosine kinase 1 | - | - | - | - | 1.194 | 2.030 | 0.872 | 7.036 × 10−23 | - | - | - | - |
| TEK | TEK receptor Tyrosine kinase | 1.083 | 5.735 | 3.093 | 4.331 × 10−32 | - | - | - | - | - | - | - | - |
| THBD | Thrombomodulin | −1.636 | 0.597 | 1.828 | 3.007 × 10−11 | 1.881 | 2.392 | 0.661 | 3.457 × 10−52 | −1.323 | 0.907 | 1.605 | 1.18302 × 10−07 |
| TP53 | Tumor protein P53 | −1.013 | 1.926 | 4.272 | 1.501 × 10−13 | −0.193 | 6.076 | 6.850 | 0.00198 | −0.207 | 4.205 | 4.719 | 0.036667 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, B.R.S.; Jara, C.P.; Sidarta-Oliveira, D.; Velloso, L.A.; Velander, W.H.; Araújo, E.P. Downregulation of the Protein C Signaling System Is Associated with COVID-19 Hypercoagulability—A Single-Cell Transcriptomics Analysis. Viruses 2022, 14, 2753. https://doi.org/10.3390/v14122753
Silva BRS, Jara CP, Sidarta-Oliveira D, Velloso LA, Velander WH, Araújo EP. Downregulation of the Protein C Signaling System Is Associated with COVID-19 Hypercoagulability—A Single-Cell Transcriptomics Analysis. Viruses. 2022; 14(12):2753. https://doi.org/10.3390/v14122753
Chicago/Turabian StyleSilva, Bruna Rafaela Santos, Carlos Poblete Jara, Davi Sidarta-Oliveira, Licio A. Velloso, William H. Velander, and Eliana P. Araújo. 2022. "Downregulation of the Protein C Signaling System Is Associated with COVID-19 Hypercoagulability—A Single-Cell Transcriptomics Analysis" Viruses 14, no. 12: 2753. https://doi.org/10.3390/v14122753
APA StyleSilva, B. R. S., Jara, C. P., Sidarta-Oliveira, D., Velloso, L. A., Velander, W. H., & Araújo, E. P. (2022). Downregulation of the Protein C Signaling System Is Associated with COVID-19 Hypercoagulability—A Single-Cell Transcriptomics Analysis. Viruses, 14(12), 2753. https://doi.org/10.3390/v14122753