
Novel 3′ Proximal Replication Elements in Umbravirus Genomes





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Construction of Umbravirus Phylogenetic Trees
2.2. Oligonucleotide-Mediated Site-Directed Mutagenesis of PEMV2 Constructs and In Vitro Transcription of PEMV2 RNA
2.3. High-Throughput SHAPE (Selective 2′ Hydroxyl Acylation Analyzed by Primer Extension) Structure Probing
2.4. Transfection of Protoplasts and RNA Gel Blots for Viral RNA Accumulation
2.5. In Vitro Translation of PEMV2 gRNA/sgRNA Transcripts in WGE
2.6. In Vivo Luciferase Translation Assays
2.7. Trans-Replication Assay
2.8. RNA Structure Drawing
3. Results
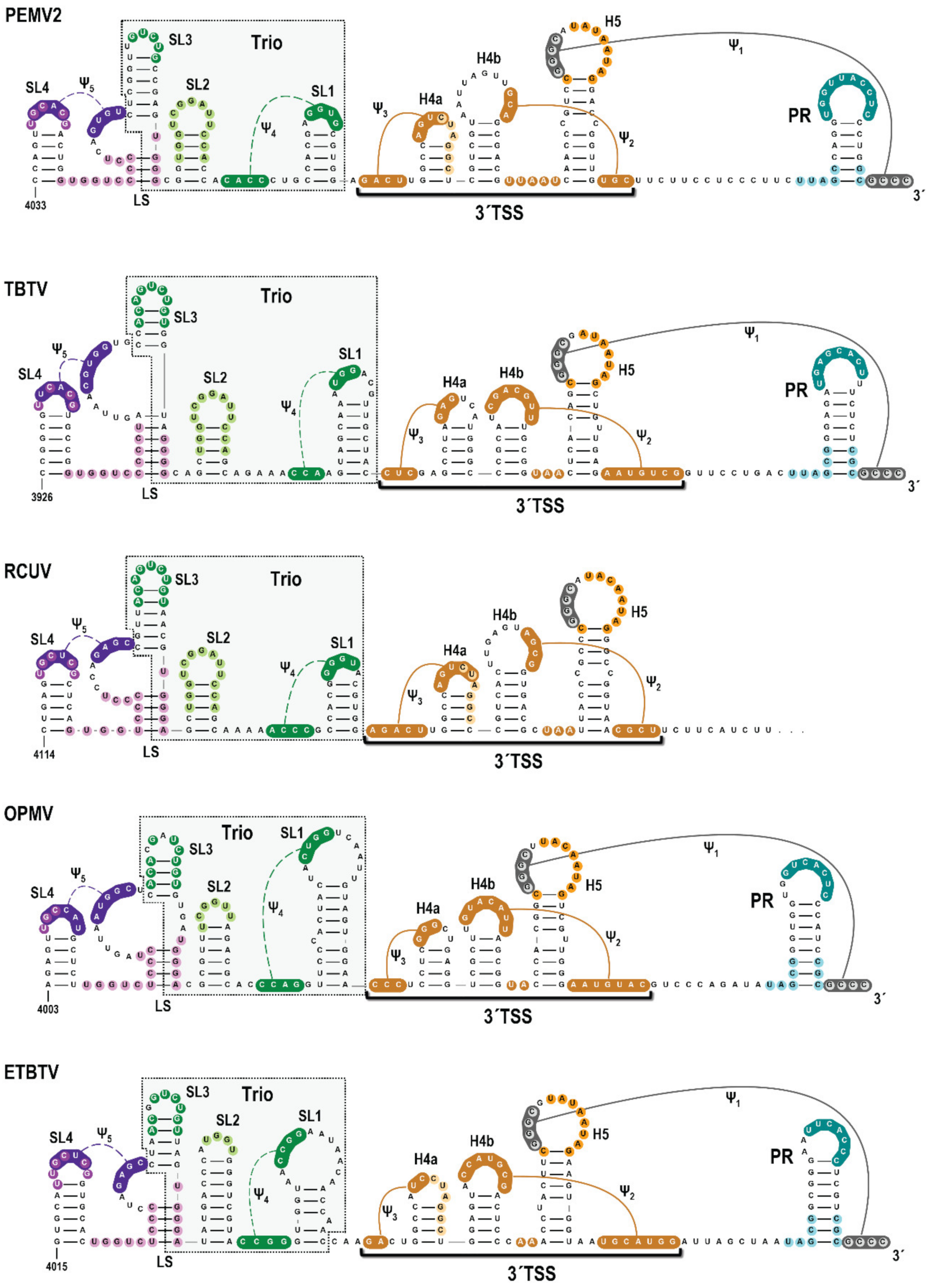
3.1. A Subset of Umbravirus 3′ Terminal Regions Contain Highly Conserved Elements
3.2. High-Throughput SHAPE Structure Probing of the 3′UTR of PEMV2 gRNA
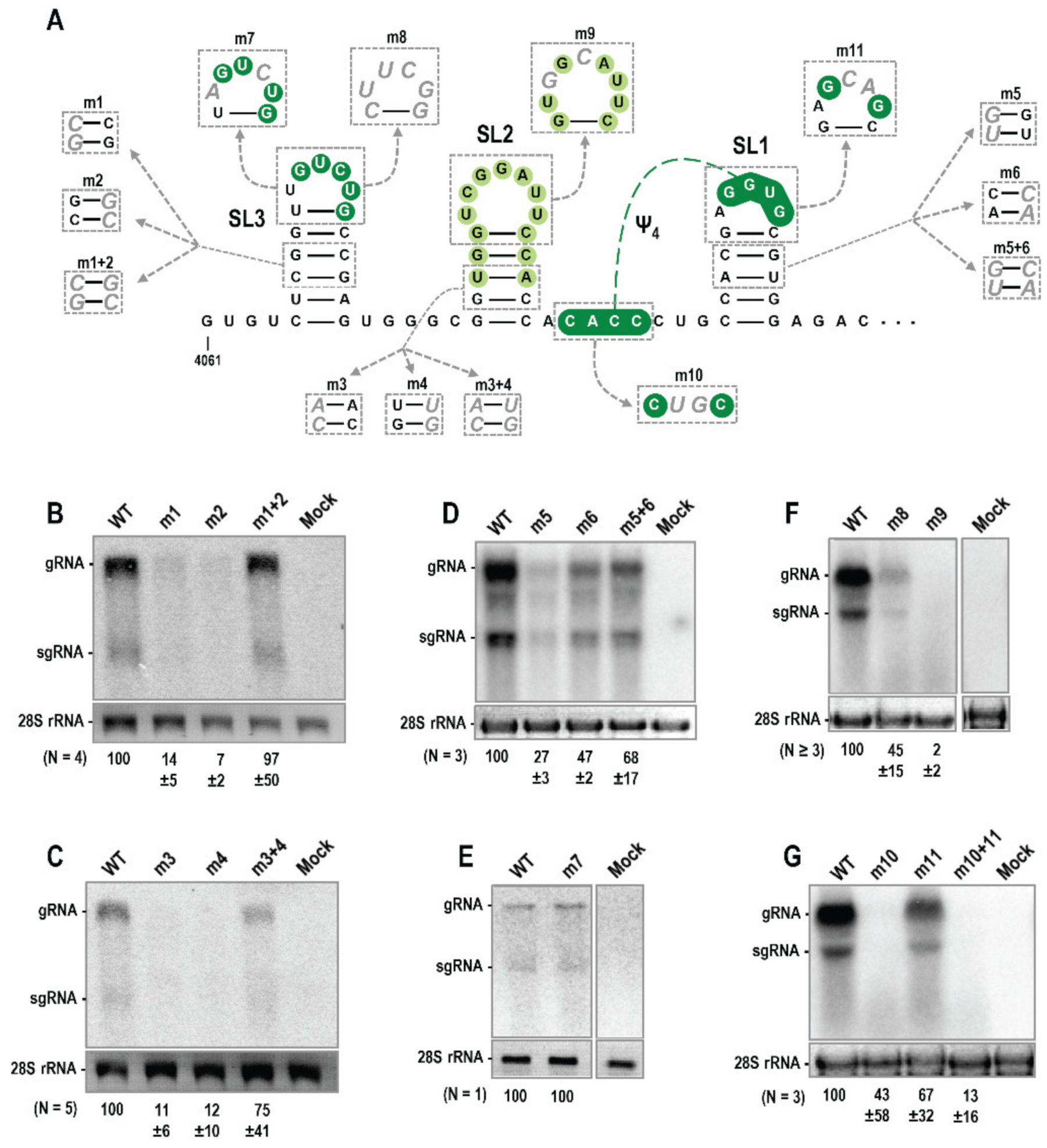
3.3. Trio Hairpins Are Critical for PEMV2 Viral Fitness



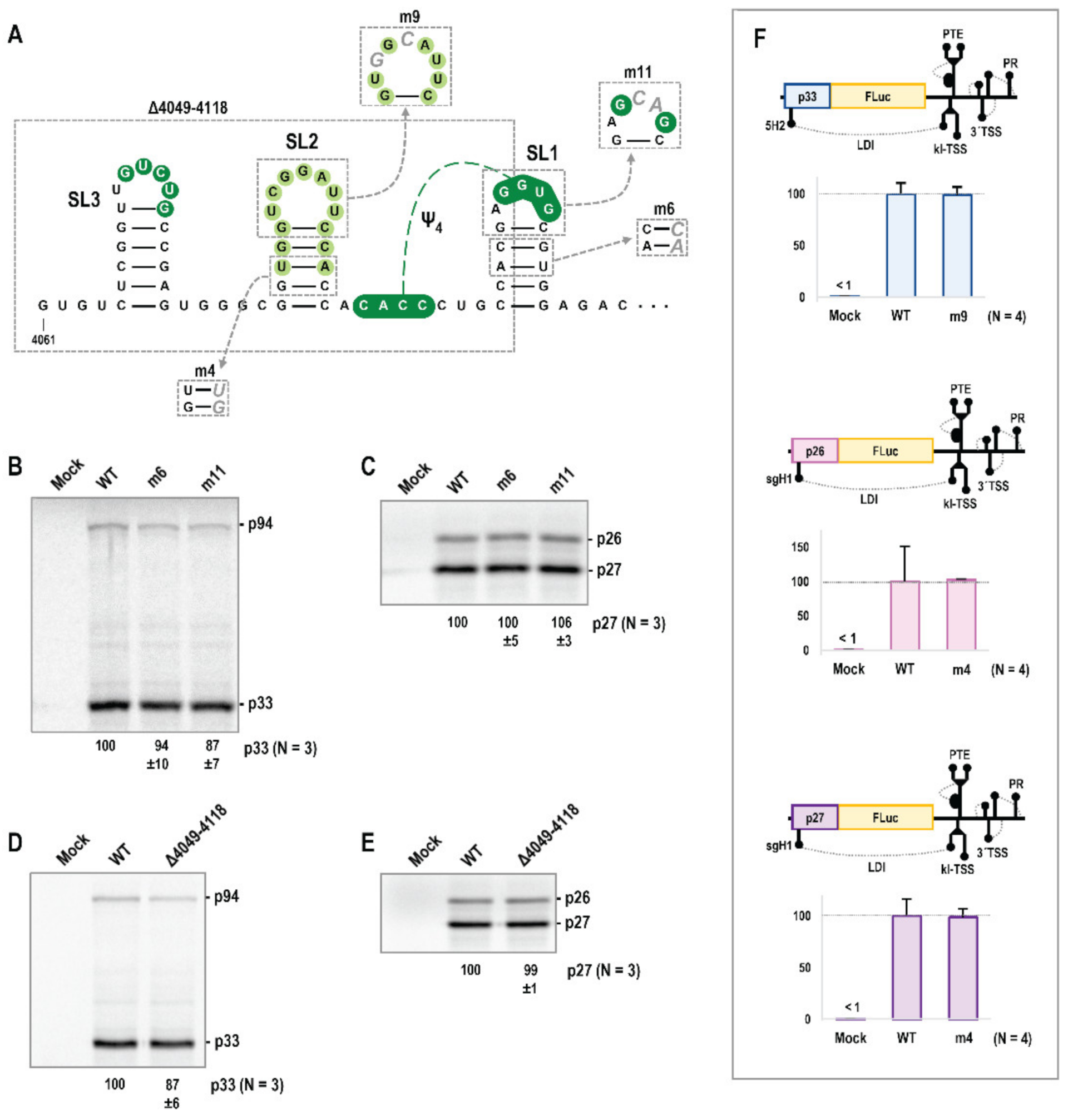
3.4. Alterations in Trio Hairpins Do Not Affect PEMV2 Translation
3.5. Trio Stem-Loops Are Important for PEMV2 Replication
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newburn, L.R.; White, K.A. Cis-acting RNA elements in positive-strand RNA plant virus genomes. Virology 2015, 479–480, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Na, H.; White, K.A. Structure and prevalence of replication silencer-3′ terminus RNA interactions in Tombusviridae. Virology 2006, 345, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.E. 3′UTRs of carmoviruses. Virus Res. 2015, 206, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.E.; Miller, W.A. 3′ cap-independent translation enhancers of plant viruses. Annu. Rev. Microbiol. 2013, 67, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Miras, M.; Miller, W.A.; Truniger, V.; Aranda, M.A. Non-canonical Translation in Plant RNA Viruses. Front. Plant Sci. 2017, 8, 494. [Google Scholar] [CrossRef]
- Pathak, K.B.; Pogany, J.; Xu, K.; White, K.A.; Nagy, P.D. Defining the roles of cis-acting RNA elements in tombusvirus replicase assembly in vitro. J. Virol. 2012, 86, 156–171. [Google Scholar] [CrossRef]
- Le, M.T.; Kasprzak, W.K.; Kim, T.; Gao, F.; Young, M.Y.; Yuan, X.; Shapiro, B.A.; Seog, J.; Simon, A.E. Folding behavior of a T-shaped, ribosome-binding translation enhancer implicated in a wide-spread conformational switch. Elife 2017, 6, e22883. [Google Scholar] [CrossRef]
- Li, Z.; Nagy, P.D. Diverse roles of host RNA binding proteins in RNA virus replication. RNA Biol. 2011, 8, 305–315. [Google Scholar] [CrossRef]
- White, K.A.; Nagy, P.D. Advances in the molecular biology of tombusviruses: Gene expression, genome replication, and recombination. Prog. Nucleic Acid Res. Mol. Biol. 2004, 78, 187–226. [Google Scholar]
- Taliansky, M.E.; Robinson, D.J. Molecular biology of umbraviruses: Phantom warriors. J. Gen. Virol. 2003, 84, 1951–1960. [Google Scholar] [CrossRef]
- Gao, F.; Simon, A.E. Multiple Cis-acting elements modulate programmed -1 ribosomal frameshifting in Pea enation mosaic virus. Nucleic Acids Res. 2016, 44, 878–895. [Google Scholar] [CrossRef] [PubMed]
- May, J.P.; Johnson, P.Z.; Ilyas, M.; Gao, F.; Simon, A.E. The multifunctional long-distance movement protein of Pea enation mosaic virus 2 protects viral and host transcripts from nonsense-mediated decay. MBio 2020, 11, e00204-20. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Roberts, I.M.; Palukaitis, P.; Taliansky, M. Host-specific cell-to-cell and long-distance movements of cucumber mosaic virus are facilitated by the movement protein of groundnut rosette virus. Virology 1999, 260, 98–108. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ryabov, E.V.; Oparka, K.J.; Santa Cruz, S.; Robinson, D.J.; Taliansky, M.E. Intracellular location of two groundnut rosette umbravirus proteins delivered by PVX and TMV vectors. Virology 1998, 242, 303–313. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Demler, S.A.; Rucker, D.G.; de Zoeten, G.A. The chimeric nature of the genome of pea enation mosaic virus: The independent replication of RNA 2. J. Gen. Virol. 1993, 74 Pt 1, 1–14. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Robinson, D.J.; Taliansky, M. Umbravirus-encoded proteins both stabilize heterologous viral RNA and mediate its systemic movement in some plant species. Virology 2001, 288, 391–400. [Google Scholar] [CrossRef]
- Gao, F.; Kasprzak, W.K.; Szarko, C.; Shapiro, B.A.; Simon, A.E. The 3′ untranslated region of Pea Enation Mosaic Virus contains two T-shaped, ribosome-binding, cap-independent translation enhancers. J. Virol. 2014, 88, 11696–11712. [Google Scholar] [CrossRef]
- Du, Z.; Alekhina, O.M.; Vassilenko, K.S.; Simon, A.E. Concerted action of two 3′ cap-independent translation enhancers increases the competitive strength of translated viral genomes. Nucleic Acids Res. 2017, 45, 9558–9572. [Google Scholar] [CrossRef][Green Version]
- Gao, F.; Gulay, S.P.; Kasprzak, W.; Dinman, J.D.; Shapiro, B.A.; Simon, A.E. The kissing-loop T-shaped structure translational enhancer of Pea enation mosaic virus can bind simultaneously to ribosomes and a 5’ proximal hairpin. J. Virol. 2013, 87, 11987–12002. [Google Scholar] [CrossRef]
- Wang, Z.; Treder, K.; Miller, W.A. Structure of a viral cap-independent translation element that functions via high affinity binding to the eIF4E subunit of eIF4F. J. Biol. Chem. 2009, 284, 14189–14202. [Google Scholar] [CrossRef]
- Wang, Z.; Parisien, M.; Scheets, K.; Miller, W.A. The cap-binding translation initiation factor, eIF4E, binds a pseudoknot in a viral cap-independent translation element. Structure 2011, 19, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Simon, A.E. Differential use of 3′CITEs by the subgenomic RNA of Pea enation mosaic virus 2. Virology 2017, 510, 194–204. [Google Scholar] [CrossRef] [PubMed]
- McCormack, J.C.; Yuan, X.; Yingling, Y.G.; Kasprzak, W.; Zamora, R.E.; Shapiro, B.A.; Simon, A.E. Structural domains within the 3′ untranslated region of Turnip crinkle virus. J. Virol. 2008, 82, 8706–8720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Simon, A.E. A multifunctional turnip crinkle virus replication enhancer revealed by in vivo functional SELEX. J. Mol. Biol. 2003, 326, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Simon, A.E. Requirement of a 3′-terminal stem-loop in in vitro transcription by an RNA-dependent RNA polymerase. J. Mol. Biol. 1995, 254, 6–14. [Google Scholar] [CrossRef]
- Stupina, V.; Simon, A.E. Analysis in vivo of turnip crinkle virus satellite RNA C variants with mutations in the 3′-terminal minus-strand promoter. Virology 1997, 238, 470–477. [Google Scholar] [CrossRef][Green Version]
- Carpenter, C.D.; Simon, A.E. Analysis of sequences and predicted structures required for viral satellite RNA accumulation by in vivo genetic selection. Nucleic Acids Res. 1998, 26, 2426–2432. [Google Scholar] [CrossRef]
- Kuhlmann, M.M.; Chattopadhyay, M.; Stupina, V.A.; Gao, F.; Simon, A.E. An RNA element that facilitates programmed ribosomal readthrough in Turnip crinkle virus adopts multiple conformations. J. Virol. 2016, 90, 8575–8591. [Google Scholar] [CrossRef]
- Cimino, P.A.; Nicholson, B.L.; Wu, B.; Xu, W.; White, K.A. Multifaceted regulation of translational readthrough by RNA replication elements in a tombusvirus. PLoS Pathog. 2011, 7, e1002423. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, J.; Simon, A.E. Repression and derepression of minus-strand synthesis in a plus-strand RNA virus replicon. J. Virol. 2004, 78, 7619–7633. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, G.; McCormack, J.C.; Simon, A.E. Evolution of virus-derived sequences for high-level replication of a subviral RNA. Virology 2006, 351, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Naismith, J.H. An efficient one-step site-directed deletion, insertion, single and multiple-site plasmid mutagenesis protocol. BMC Biotechnol. 2008, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Shi, K.; Simon, A.E. A local, interactive network of 3′ RNA elements supports translation and replication of Turnip crinkle virus. J. Virol. 2012, 86, 4065–4081. [Google Scholar] [CrossRef]
- Ilyas, M.; Du, Z.; Simon, A.E. Opium poppy mosaic virus has an XRN-resistant, translated subgenomic RNA and a BTE 3′ CITE. J. Virol. 2021, 95, e02109-20. [Google Scholar] [CrossRef]
- Wilkinson, K.A.; Merino, E.J.; Weeks, K.M. Selective 2’-hydroxyl acylation analyzed by primer extension (SHAPE): Quantitative RNA structure analysis at single nucleotide resolution. Nat. Protoc. 2006, 1, 1610–1616. [Google Scholar] [CrossRef]
- Karabiber, F.; McGinnis, J.L.; Favorov, O.V.; Weeks, K.M. QuShape: Rapid, accurate, and best-practices quantification of nucleic acid probing information, resolved by capillary electrophoresis. RNA 2013, 19, 63–73. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, G.; Guo, R.; Shapiro, B.A.; Simon, A.E. A pseudoknot in a preactive form of a viral RNA is part of a structural switch activating minus-strand synthesis. J. Virol. 2006, 80, 9181–9191. [Google Scholar] [CrossRef]
- McCormack, J.C.; Simon, A.E. Callus cultures of Arabidopsis. Curr. Protoc. Microbiol. 2006, 1, 16D-1. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Gao, F.; Kasprzak, W.; Stupina, V.A.; Shapiro, B.A.; Simon, A.E. A ribosome-binding, 3′ translational enhancer has a T-shaped structure and engages in a long-distance RNA-RNA interaction. J. Virol. 2012, 86, 9828–9842. [Google Scholar] [CrossRef] [PubMed]
- Stupina, V.A.; Yuan, X.; Meskauskas, A.; Dinman, J.D.; Simon, A.E. Ribosome binding to a 5’ translational enhancer is altered in the presence of the 3′ untranslated region in cap-independent translation of turnip crinkle virus. J. Virol. 2011, 85, 4638–4653. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Yim, H.S.; Ryu, J.Y.; Lee, H.S.; Lee, J.H.; Seen, D.S.; Kang, S.G. One-step sequence- and ligation-independent cloning as a rapid and versatile cloning method for functional genomics studies. Appl. Environ. Microbiol. 2012, 78, 5440–5443. [Google Scholar] [CrossRef] [PubMed]
- Mérai, Z.; Kerényi, Z.; Molnár, A.; Barta, E.; Válóczi, A.; Bisztray, G.; Havelda, Z.; Burgyán, J.; Silhavy, D. Aureusvirus P14 is an efficient RNA silencing suppressor that binds double-stranded RNAs without size specificity. J. Virol. 2005, 79, 7217–7226. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Glazebrook, J. Transformation of agrobacterium using electroporation. CSH Protoc. 2006, 2006, 7. [Google Scholar] [CrossRef]
- Johnson, P.Z.; Kasprzak, W.K.; Shapiro, B.A.; Simon, A.E. RNA2Drawer: Geometrically strict drawing of nucleic acid structures with graphical structure editing and highlighting of complementary subsequences. RNA Biol. 2019, 16, 1667–1671. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Wilkinson, K.A.; Gorelick, R.J.; Vasa, S.M.; Guex, N.; Rein, A.; Mathews, D.H.; Giddings, M.C.; Weeks, K.M. High-throughput SHAPE analysis reveals structures in HIV-1 genomic RNA strongly conserved across distinct biological states. PLoS Biol. 2008, 6, e96. [Google Scholar] [CrossRef]
- Karabiber, F. A peak alignment algorithm with novel improvements in application to electropherogram analysis. J. Bioinform. Comput. Biol. 2013, 11, 1350011. [Google Scholar] [CrossRef]
- Nicholson, B.L.; White, K.A. Context-influenced cap-independent translation of Tombusvirus mRNAs in vitro. Virology 2008, 380, 203–212. [Google Scholar] [CrossRef][Green Version]
- Stupina, V.A.; Meskauskas, A.; McCormack, J.C.; Yingling, Y.G.; Shapiro, B.A.; Dinman, J.D.; Simon, A.E. The 3′ proximal translational enhancer of Turnip crinkle virus binds to 60S ribosomal subunits. RNA 2008, 14, 2379–2393. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, P.Z.; Reuning, H.M.; Bera, S.; Gao, F.; Du, Z.; Simon, A.E. Novel 3′ Proximal Replication Elements in Umbravirus Genomes. Viruses 2022, 14, 2615. https://doi.org/10.3390/v14122615
Johnson PZ, Reuning HM, Bera S, Gao F, Du Z, Simon AE. Novel 3′ Proximal Replication Elements in Umbravirus Genomes. Viruses. 2022; 14(12):2615. https://doi.org/10.3390/v14122615
Chicago/Turabian StyleJohnson, Philip Z., Hannah M. Reuning, Sayanta Bera, Feng Gao, Zhiyou Du, and Anne E. Simon. 2022. "Novel 3′ Proximal Replication Elements in Umbravirus Genomes" Viruses 14, no. 12: 2615. https://doi.org/10.3390/v14122615
APA StyleJohnson, P. Z., Reuning, H. M., Bera, S., Gao, F., Du, Z., & Simon, A. E. (2022). Novel 3′ Proximal Replication Elements in Umbravirus Genomes. Viruses, 14(12), 2615. https://doi.org/10.3390/v14122615