

Weed Hosts Represent an Important Reservoir of Turnip Yellows Virus and a Possible Source of Virus Introduction into Oilseed Rape Crop

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling of Oilseed Rape and Weed Plants from Fields
2.2. Detection of TuYV by RT-PCR
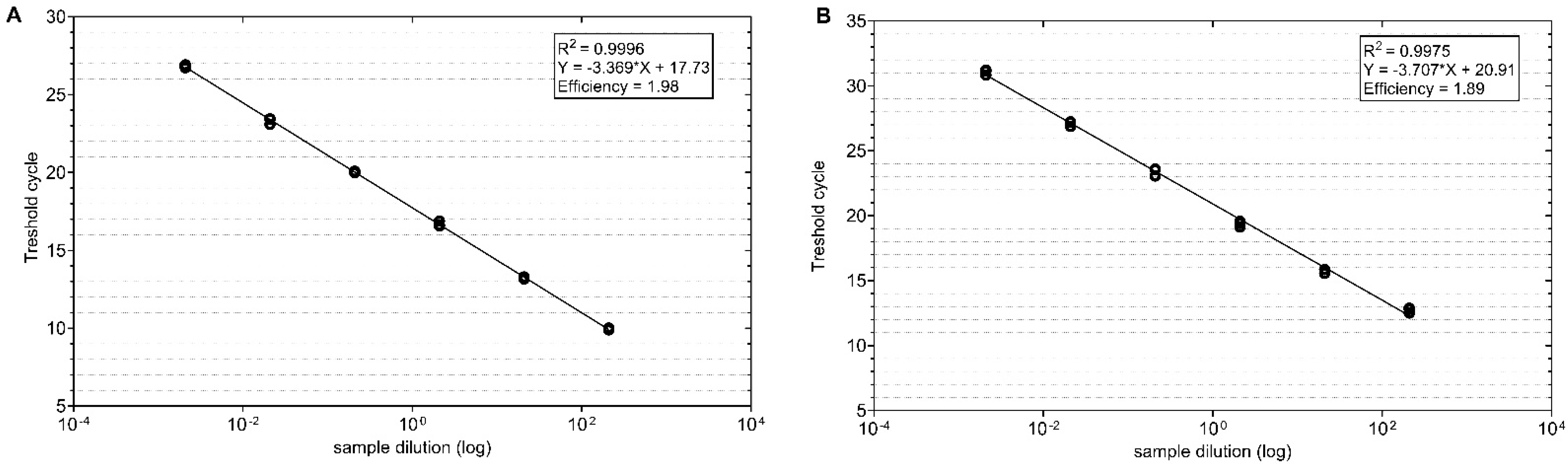
2.3. Analysis of TuYV Titre in Oilseed Rape and Weed Plants by RT-qPCR
2.4. Sequence Analysis of Readthrough Domain (RTD) Gene Sequence
2.5. High-Throughput Analysis of the PK2 P. rhoeas Sample
2.6. Multiple Sequence Alignment and Phylogenetic Analysis
3. Results and Discussion
3.1. Detection of TuYV in Oilseed Rape and Weed Hosts
3.2. TuYV Titre in Oilseed Rape and Weed Plants
3.3. The Diversity of TuYV Sequences from Crop and Non-Crop Host
3.4. Virome Analysis of a Common Poppy (P. rhoeas) Plant
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, J.A.; Tomlinson, J.A. Viruses infecting winter oilseed rape (Brassica-napus ssp oleifera). Ann. Appl. Biol. 1985, 107, 485–495. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Sharman, M.; Trebicki, P.; Maina, S.; Congdon, B.S. Virus diseases of cereal and oilseed crops in Australia: Current position and future challenges. Viruses 2021, 13, 2051. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, N.V.; Davies, J.M.L.; Wright, D.M. The incidence of three diseases of winter oilseed rape in England and Wales in the 1991/92 and 1992/93 growing season. Plant Pathol. 1994, 43, 1045–1049. [Google Scholar] [CrossRef]
- Duffus, J.E. Host relationship of beet western yellows virus strains. Phytopathology 1964, 54, 736–738. [Google Scholar]
- Graichen, K.; Rabenstein, F. European isolates of Beet western yellows virus (BWYV) from oilseed rape (Brassica napus L. ssp. napus) are non-pathogenic on sugar beet (Beta vulgaris L. var. altissima) but represent isolates of Turnip yellows virus (TuYV). J. Plant Dis. Prot. 1996, 103, 233–245. [Google Scholar]
- Wallis, R.L. Some host plants of the green peach aphid and beet western yellows virus in the Pacific Northwest. J. Econ. Entomol. 1967, 60, 904–907. [Google Scholar] [CrossRef]
- Goyal, G.; Harsimran, K.G.; McSorley, R. Common Weed Hosts of Insect-Transmitted Viruses of Florida Vegetable Crops; ENY-863; University of Florida Institute of Food and Agricultural Sciences: Gainesville, FL, USA, 2022; Available online: https://edis.ifas.ufl.edu/publication/in931 (accessed on 29 April 2022).
- Stevens, M.; Smith, H.G.; Hallsworth, P.B. The host-range of Beet yellowing viruses among common arable weed species. Plant Pathol. 1994, 43, 579–588. [Google Scholar] [CrossRef]
- Stevens, M.; McGrann, G.; Clark, B.; Authority, H. Turnip yellows virus (syn Beet western yellows virus): An emerging threat to European oilseed rape production? HGCA Res. Rev. 2008, 69, 1–37. [Google Scholar]
- Freeman, A.; Aftab, M. Effective management of viruses in pulse crops in south eastern Australia should include management of weeds. Australas. Plant Pathol. 2011, 40, 430–441. [Google Scholar] [CrossRef]
- Chalupníková, J.; Kundu, J.K.; Singh, K.; Bartakova, P.; Beoni, E. Wheat streak mosaic virus: Incidence in field crops, potential reservoir within grass species and uptake in winter wheat cultivars. J. Integr. Agric. 2017, 16, 523–531. [Google Scholar] [CrossRef]
- Rashidi, M.; Cruzado, K.R.; Hutchinson, S.J.P.; Bosque-Pérez, A.N.; Marshall, M.J.; Rashed, A. Grassy weeds and corn as potential sources of barley yellow dwarf virus spread into winter wheat. Plant Dis. 2021, 105, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Yazdkhasti, E.; Hopkins, J.R.; Kvarnheden, A. Reservoirs of plant virus disease: Occurrence of wheat dwarf virus and barley/cereal yellow dwarf viruses in Sweden. Plant Pathol. 2021, 70, 1552–1561. [Google Scholar] [CrossRef]
- Souza, T.A.; Macedo, M.A.; Albuquerque, L.C.; Inoue-Nagata, K.A. Host range and natural infection of tomato chlorosis virus in weeds collected in Central Brazil. Trop. Plant Pathol. 2020, 45, 84–90. [Google Scholar] [CrossRef]
- Korbecka-Glinka, G.; Przyby´s, M.; Feledyn-Szewczyk, B.A. Survey of five plant viruses in weeds and tobacco in Poland. Agronomy 2021, 11, 1667. [Google Scholar] [CrossRef]
- Hancinsky, R.; Mihalik, D.; Mrkvova, M.; Candresse, T.; Glasa, M. Plant viruses infecting Solanaceae family members in the cultivated and wild environments: A review. Plants 2020, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- Maliano, M.R.; Macedo, M.A.; Rojas, M.R.; Gilbertson, R.L. Weed-infecting viruses in a tropical agroecosystem present different threats to crops and evolutionary histories. PLoS ONE 2021, 16, e0250066. [Google Scholar] [CrossRef]
- Chen, G.; Pan, H.; Xie, W.; Wang, S.; Wu, Q.; Fang, Y.; Shi, X.; Zhang, Y. Virus infection of a weed increases vector attraction to and vector fitness on the weed. Sci. Rep. 2013, 3, 2253. [Google Scholar] [CrossRef]
- Sõmera, M.; Fargette, D.; Hébrard, E.; Sarmiento, C. ICTV Report Consortium: ICTV Virus Taxonomy Profile: Solemoviridae. J. Gen. Virol. 2021, 102, 001707. [Google Scholar]
- Stevens, M.; Freeman, B.; Liu, H.Y.; Herrbach, E.; Lemaire, O. Beet poleroviruses: Close friends or distant relatives? Mol. Plant Pathol. 2005, 6, 1–9. [Google Scholar] [CrossRef]
- Pfeffer, S.; Dunoyer, P.; Heim, F.; Richards, K.E.; Jonard, G.; Ziegler-Graff, V. P0 of Beet western yellows virus is a suppressor of posttranscriptional gene silencing. J. Virol. 2002, 76, 6815–6824. [Google Scholar] [CrossRef]
- Bortolamiol, D.; Pazhouhandeh, M.; Marrocco, K.; Genschik, P.; Ziegler-Graff, V. The Polerovirus F box protein P0 targets ARGONAUTE1 to suppress RNA silencing. Curr. Biol. 2007, 17, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Reutenauer, A.; Ziegler-Graff, V.; Lot, H.; Scheidecker, D.; Guilley, H.; Richards, K.; Jonard, G. Identification of Beet western yellows virus luteovirus genes implicated in viral replication and particle morphogenesis. Virology 1993, 195, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.R.; Keese, P.K.; Young, M.J.; Waterhouse, P.M.; Gerlach, W.L. Evolution and molecular biology of luteoviruses. Annu. Rev. Phytopathol. 1990, 28, 341–363. [Google Scholar] [CrossRef]
- Brault, V.; van den Heuvel, J.F.; Verbeek, M.; Ziegler-Graff, V.; Reutenauer, A.; Herrbach, E.; Garaud, J.C.; Guilley, H.; Richards, K.; Jonard, G. Aphid transmission of beet western yellows luteovirus requires the minor capsid read-through protein P74. EMBO J. 1995, 14, 650–659. [Google Scholar] [CrossRef]
- Van den Heuvel, J.F.J.M.; Bruyère, A.; Hogenhout, A.; Ziegler-Graff, V.; Brault, V.; Verbeek, M.; van der Wilk, F.; Richards, K. The N-terminal region of the luteovirus readthrough domain determines virus binding to Buchnera GroEL and is essential for virus persistence in the aphid. J. Virol. 1997, 71, 7258–7265. [Google Scholar] [CrossRef] [PubMed]
- Peter, K.A.; Gildow, F.; Palukaitis, P.; Gray, S.M. The C terminus of the polerovirus P5 readthrough domain limits virus infection to the phloem. J. Virol. 2009, 83, 5419–5429. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Firth, A.E.; Miller, W.A.; Scheidecker, D.; Brault, V.; Reinbold, C.; Rakotondrafara, A.M.; Chung, B.Y.W.; Ziegler-Graff, V. Discovery of a small non-AUG-initiated ORF in poleroviruses and luteoviruses that is required for long-distance movement. PLoS Pathog. 2015, 11, e1004868. [Google Scholar] [CrossRef] [PubMed]
- Filardo, F.; Nancarrow, N.; Kehoe, M.; McTaggart, A.R.; Congdon, B.; Kumari, S.; Aftab, M.; Trębicki, P.; Rodoni, B.; Thomas, J.; et al. Genetic diversity and recombination between turnip yellows virus strains in Australia. Arch. Virol. 2021, 166, 813–829. [Google Scholar] [CrossRef]
- Brault, V.; Périgon, S.; Reinbold, C.; Erdinger, M.; Scheidecker, D.; Herrbach, E.; Richards, K.; Ziegler-Graff, V. The polerovirus minor capsid protein determines vector specificity and intestinal tropism in the aphid. J. Virol. 2005, 79, 9685–9693. [Google Scholar] [CrossRef]
- Hanzlik, K.; Gerowitt, B. Occurrence and distribution of important weed species in German winter oilseed rape fields. J. Plant Dis. Prot. 2012, 119, 107–120. [Google Scholar] [CrossRef]
- Krähmer, H.; Andreasen, C.; Economou-Antonaka, G.; Holec, J.; Kalivas, D.; Kolářová, M.; Novák, R.; Panozzog, S.; Pinke, G.; Salonen, J.; et al. Weed surveys and weed mapping in Europe: State of the art and future tasks. Crop Prot. 2020, 129, 105010. [Google Scholar] [CrossRef]
- Abraham, A.D.; Menzel, W.; Lesemann, D.E.; Varrelmann, M.; Vetten, H.J. Chickpea chlorotic stunt virus: A new polerovirus infecting cool-season food legumes in ethiopia. Phytopathology 2006, 96, 437–446. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Singh, K.; Slavíková, L.; Kumar, J. Reakční Směs pro Kvantifikaci Viru Žloutenky Vodnice v Kulturních a Nekulturních Vzorcích Pomocí qPCR (Reaction Mixture for Quantification of Turnip Yellows Virus in Cultivated and Uncultivated Plant Samples by qPCR); Užitný Vzor (Utility Model) no. 35269; Industrial Property Office: Prague, Czech Republic, 2021. [Google Scholar]
- Jarošová, J.; Kundu, J.K. Validation of reference genes as internal control for studying viral infections in cereals by quantitative real-time RT-PCR. BMC Plant Biol. 2010, 10, 146. [Google Scholar] [CrossRef] [PubMed]
- Dráb, T.; Svobodová, E.; Ripl, J.; Jarošová, J.; Rabenstein, F.; Melcher, U.; Kundu, J.K. SYBR Green I based RT-qPCR assays for the detection of RNA viruses of cereals and grasses. Crop Pasture Sci. 2014, 65, 1323–1328. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Walkey, D.G.A.; Pink, D.A.C. Studies on resistance to Beet western yellows virus in lettuce (Lactuca sativa) and the occurrence of field sources of the virus. Plant Pathol. 1990, 39, 141–155. [Google Scholar] [CrossRef]
- Pallett, D.W.; Thurston, M.I.; Cortina-Borja, M.; Edwards, M.L.; Alexander, M.; Mitchell, E.; Raybould, A.F.; Cooper, J.I. The incidence of viruses in wild Brassica rapa ssp. sylvestris in southern England. Ann. Appl. Biol. 2002, 141, 163–170. [Google Scholar] [CrossRef]
- Thurston, M.I.; Pallett, D.W.; Cortina-Borja, M.; Edwards, M.L.; Raybould, A.F.; Cooper, J.I. The incidence of viruses in wild Brassica nigra in Dorset (UK). Ann. Appl. Biol. 2001, 139, 277–284. [Google Scholar] [CrossRef]
- Coutts, B.A.; Hawkes, J.R.; Jones, R.A.C. Occurrence of Beet western yellows virus and its aphid vectors in over-summering broad-leafed weeds and volunteer crop plants in the grainbelt region of south-western Australia. Aust. J. Agric. Res. 2006, 57, 975–982. [Google Scholar] [CrossRef]
- Kolářová, M.; Tyšer, J.; Soukup, J. Survey about the weed occurrence on arable land in the Czech Republic. Sci. Agric. Bohem. 2013, 44, 63–69. [Google Scholar] [CrossRef]
- Jursík, M.; Holec, J.; Soukup, J.; Venclová, V. Competitive relationships between sugar beet and weeds in dependence on time of weed control. Plant Soil Environ. 2008, 54, 108–116. [Google Scholar] [CrossRef]
- Latham, L.J.; Smith, L.J.; Jones, R.A.C. Incidence of three viruses in vegetable brassica plantings and associated wild radish weeds in south-west Australia. Australas. Plant Pathol. 2003, 32, 387–391. [Google Scholar] [CrossRef]
- Primot, S.; Valantin-Morison, M.; Makowski, D. Predicting the risk of weed infestation in winter oilseed rape crops. Weed Res. 2005, 46, 22–33. [Google Scholar] [CrossRef]
- Maree, H.J.; Fox, A.; Al Rwahnih, M.; Boonham, N.; Candresse, T. Application of HTS for routine plant virus diagnostics: State of the art and challenges. Front. Plant Sci. 2018, 9, 1082. [Google Scholar] [CrossRef]
- Mrkvová, M.; Hančinský, R.; Predajňa, L.; Alaxin, P.; Achs, A.; Tomašechová, J.; Šoltys, K.; Mihálik, D.; Olmos, A.; Ruiz-García, A.B.; et al. High-throughput sequencing discloses the Cucumber mosaic virus (CMV) diversity in Slovakia and reveals new hosts of CMV from the Papaveraceae family. Plants 2022, 11, 1665. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slavíková, L.; Ibrahim, E.; Alquicer, G.; Tomašechová, J.; Šoltys, K.; Glasa, M.; Kundu, J.K. Weed Hosts Represent an Important Reservoir of Turnip Yellows Virus and a Possible Source of Virus Introduction into Oilseed Rape Crop. Viruses 2022, 14, 2511. https://doi.org/10.3390/v14112511
Slavíková L, Ibrahim E, Alquicer G, Tomašechová J, Šoltys K, Glasa M, Kundu JK. Weed Hosts Represent an Important Reservoir of Turnip Yellows Virus and a Possible Source of Virus Introduction into Oilseed Rape Crop. Viruses. 2022; 14(11):2511. https://doi.org/10.3390/v14112511
Chicago/Turabian StyleSlavíková, Lucie, Emad Ibrahim, Glenda Alquicer, Jana Tomašechová, Katarína Šoltys, Miroslav Glasa, and Jiban Kumar Kundu. 2022. "Weed Hosts Represent an Important Reservoir of Turnip Yellows Virus and a Possible Source of Virus Introduction into Oilseed Rape Crop" Viruses 14, no. 11: 2511. https://doi.org/10.3390/v14112511
APA StyleSlavíková, L., Ibrahim, E., Alquicer, G., Tomašechová, J., Šoltys, K., Glasa, M., & Kundu, J. K. (2022). Weed Hosts Represent an Important Reservoir of Turnip Yellows Virus and a Possible Source of Virus Introduction into Oilseed Rape Crop. Viruses, 14(11), 2511. https://doi.org/10.3390/v14112511