

Interspecies Transmission from Pigs to Ferrets of Antigenically Distinct Swine H1 Influenza A Viruses with Reduced Reactivity to Candidate Vaccine Virus Antisera as Measures of Relative Zoonotic Risk

 , , ,
, , ,
Abstract
1. Introduction
2. Material and Methods
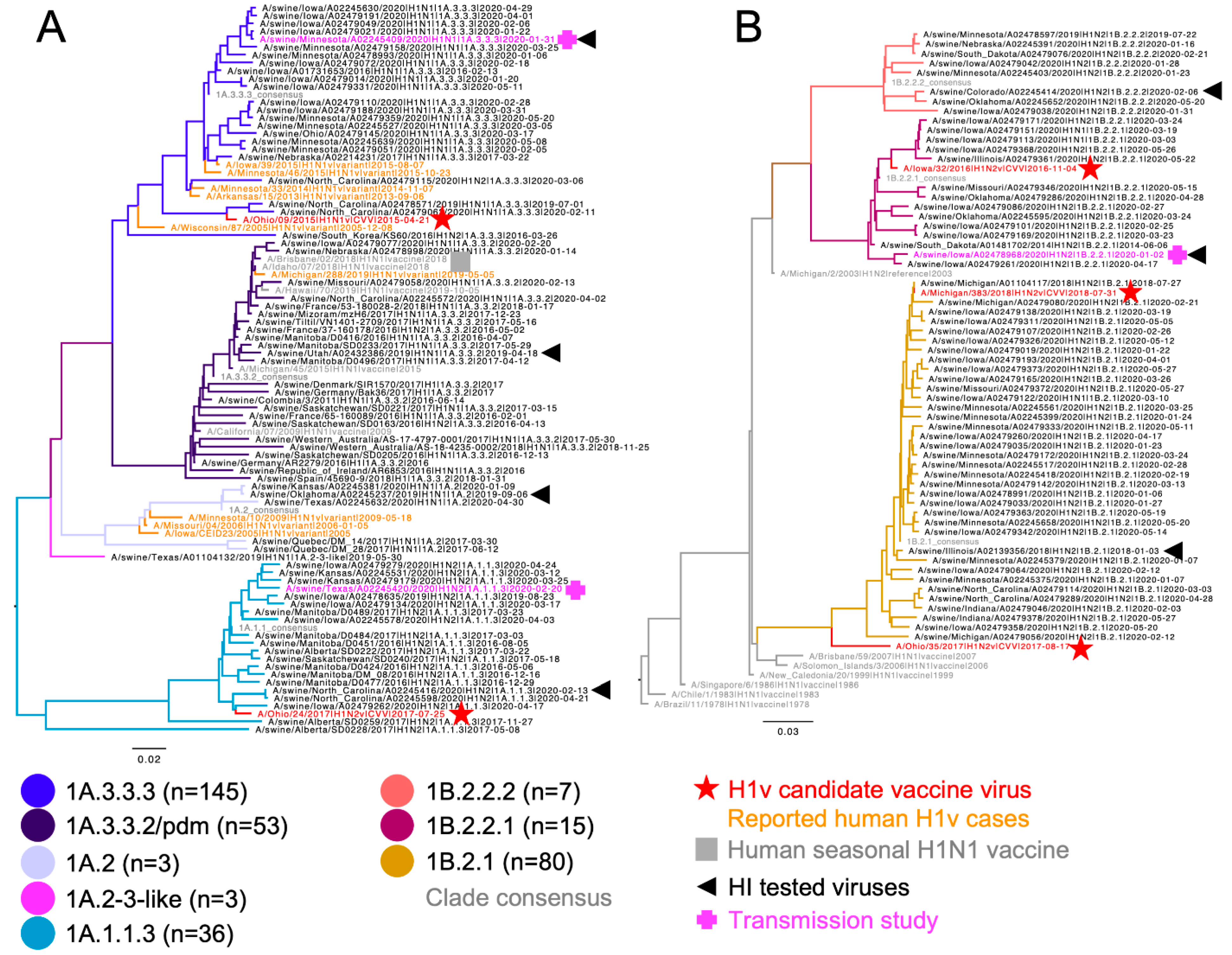
2.1. Genetic Analysis and Strain Selection
2.2. Viruses and Ferret Antisera
2.3. Hemagglutination Inhibition
2.4. Swine-to-Ferret Transmission Study Design
2.5. Virus Replication and Shedding
2.6. Pathology Examination
2.7. Microbiological Assays
2.8. Data Analysis
3. Results
3.1. Genetic and Phylogenetic Characterization of US Swine H1 Hemagglutinin
3.2. Dominant U.S. Swine H1 Strains Drifted from Human Seasonal H1 or CVV
3.3. Swine-to-Ferret Transmission
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lambert, L.C.; Fauci, A.S. Influenza vaccines for the future. N. Engl. J. Med. 2010, 363, 2036–2044. [Google Scholar] [CrossRef] [PubMed]
- Girard, M.P.; Cherian, T.; Pervikov, Y.; Kieny, M.P. A review of vaccine research and development: Human acute respiratory infections. Vaccine 2005, 23, 5708–5724. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Estimates of deaths associated with seasonal influenza—United States, 1976–2007. Morb. Mortal. Wkly. Rep. 2010, 59, 1057–1062. [Google Scholar]
- Zeller, M.A.; Anderson, T.K.; Walia, R.W.; Vincent, A.L.; Gauger, P.C. ISU FLUture: A veterinary diagnostic laboratory web-based platform to monitor the temporal genetic patterns of Influenza A virus in swine. BMC Bioinform. 2018, 19, 397. [Google Scholar] [CrossRef] [PubMed]
- Arendsee, Z.W.; Chang, J.; Hufnagel, D.E.; Markin, A.; Janas-Martindale, A.; Vincent, A.L.; Anderson, T.K. octoFLUshow: An Interactive Tool Describing Spatial and Temporal Trends in the Genetic Diversity of Influenza A Virus in U.S. Swine. Microbiol. Resour. Announc. 2021, 10, e01081-21. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.I.; Vincent, A.L. Reverse zoonosis of influenza to swine: New perspectives on the human-animal interface. Trends Microbiol. 2015, 23, 142–153. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Smith, G.J.; Pybus, O.G.; Zhu, H.; Bhatt, S.; Poon, L.L.; Riley, S.; Bahl, J.; Ma, S.K.; Cheung, C.L.; et al. Long-term evolution and transmission dynamics of swine influenza A virus. Nature 2011, 473, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Rajao, D.S.; Gauger, P.C.; Anderson, T.K.; Lewis, N.S.; Abente, E.J.; Killian, M.L.; Perez, D.R.; Sutton, T.C.; Zhang, J.; Vincent, A.L. Novel Reassortant Human-Like H3N2 and H3N1 Influenza A Viruses Detected in Pigs Are Virulent and Antigenically Distinct from Swine Viruses Endemic to the United States. J. Virol. 2015, 89, 11213–11222. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Noda, T.; Kawaoka, Y. Emergence and pandemic potential of swine-origin H1N1 influenza virus. Nature 2009, 459, 931–939. [Google Scholar] [CrossRef]
- Anderson, T.K.; Macken, C.A.; Lewis, N.S.; Scheuermann, R.H.; Van Reeth, K.; Brown, I.H.; Swenson, S.L.; Simon, G.; Saito, T.; Berhane, Y.; et al. A Phylogeny-Based Global Nomenclature System and Automated Annotation Tool for H1 Hemagglutinin Genes from Swine Influenza A Viruses. Msphere 2016, 1, e00275-16. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, D.; Anderson, T.K.; Kimble, J.B.; Chang, J.; Lopes, S.; Souza, C.K.; Pekosz, A.; Shaw-Saliba, K.; Rothman, R.E.; Chen, K.-F. Antigenic characterization and pandemic risk assessment of North American H1 influenza A viruses circulating in swine. Spectrum 2022. [Google Scholar]
- Nelson, M.I.; Stratton, J.; Killian, M.L.; Janas-Martindale, A.; Vincent, A.L. Continual Reintroduction of Human Pandemic H1N1 Influenza A Viruses into Swine in the United States, 2009 to 2014. J. Virol. 2015, 89, 6218–6226. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.C.F.; Bahl, J.; Joseph, U.; Butt, K.M.; Peck, H.A.; Koay, E.S.C.; Oon, L.L.E.; Barr, I.G.; Vijaykrishna, D.; Smith, G.J.D. Phylodynamics of H1N1/2009 influenza reveals the transition from host adaptation to immune-driven selection. Nat. Commun. 2015, 6, 7952. [Google Scholar] [CrossRef] [PubMed]
- Vijaykrishna, D.; Poon, L.L.; Zhu, H.C.; Ma, S.K.; Li, O.T.; Cheung, C.L.; Smith, G.J.; Peiris, J.S.; Guan, Y. Reassortment of pandemic H1N1/2009 influenza A virus in swine. Science 2010, 328, 1529. [Google Scholar] [CrossRef] [PubMed]
- WHO. Summary of Status of Development and Availability of Variant Influenza A(H1) Candidate Vaccine Viruses and Potency Testing Reagents. 2020. Available online: https://www.who.int/influenza/vaccines/virus/candidates_reagents/summary_a_h1v_cvv_sh2021_20200930.pdf?ua=1 (accessed on 15 June 2020).
- Robertson, J.S.; Nicolson, C.; Harvey, R.; Johnson, R.; Major, D.; Guilfoyle, K.; Roseby, S.; Newman, R.; Collin, R.; Wallis, C.; et al. The development of vaccine viruses against pandemic A(H1N1) influenza. Vaccine 2011, 29, 1836–1843. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.K.; Chang, J.; Arendsee, Z.W.; Venkatesh, D.; Souza, C.K.; Kimble, J.B.; Lewis, N.S.; Davis, C.T.; Vincent, A.L. Swine Influenza A Viruses and the Tangled Relationship with Humans. Cold Spring Harb Perspect. Med. 2021, 11, a038737. [Google Scholar] [CrossRef]
- Shu, Y.; McCauley, J. GISAID: Global initiative on sharing all influenza data—from vision to reality. Eurosurveill 2017, 22, 30494. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Chang, J.; Anderson, T.K.; Zeller, M.A.; Gauger, P.C.; Vincent, A.L. octoFLU: Automated Classification for the Evolutionary Origin of Influenza A Virus Gene Sequences Detected in US Swine. Microbiol. Resour. Announc. 2019, 8, e00673-19. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kaplan, B.S.; Kimble, J.B.; Chang, J.; Anderson, T.K.; Gauger, P.C.; Janas-Martindale, A.; Killian, M.L.; Bowman, A.S.; Vincent, A.L. Aerosol Transmission from Infected Swine to Ferrets of an H3N2 Virus Collected from an Agricultural Fair and Associated with Human Variant Infections. J. Virol. 2020, 94, e01009-20. [Google Scholar] [CrossRef] [PubMed]
- Gauger, P.C.; Vincent, A.L.; Loving, C.L.; Henningson, J.N.; Lager, K.M.; Janke, B.H.; Kehrli, M.E., Jr.; Roth, J.A. Kinetics of lung lesion development and pro-inflammatory cytokine response in pigs with vaccine-associated enhanced respiratory disease induced by challenge with pandemic (2009) A/H1N1 influenza virus. Vet. Pathol. 2012, 49, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Gregory, V.; Collins, P.; Kloess, J.; Wharton, S.; Cattle, N.; Lackenby, A.; Daniels, R.; Hay, A. Neuraminidase receptor binding variants of human influenza A(H3N2) viruses resulting from substitution of aspartic acid 151 in the catalytic site: A role in virus attachment? J. Virol. 2010, 84, 6769–6781. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Opriessnig, T.; Yu, S.; Gallup, J.M.; Evans, R.B.; Fenaux, M.; Pallares, F.; Thacker, E.L.; Brockus, C.W.; Ackermann, M.R.; Thomas, P.; et al. Effect of vaccination with selective bacterins on conventional pigs infected with type 2 porcine circovirus. Vet. Pathol. 2003, 40, 521–529. [Google Scholar] [CrossRef]
- Kilbourne, E.D. Influenza pandemics of the 20th century. Emerg. Infect. Dis. 2006, 12, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.I.; Lemey, P.; Tan, Y.; Vincent, A.; Lam, T.T.-Y.; Detmer, S.; Viboud, C.; Suchard, M.A.; Rambaut, A.; Holmes, E.C. Spatial dynamics of human-origin H1 influenza A virus in North American swine. PLoS Pathog. 2011, 7, e1002077. [Google Scholar] [CrossRef]
- Neveau, M.N.; Zeller, M.A.; Kaplan, B.S.; Souza, C.K.; Gauger, P.C.; Vincent, A.L.; Anderson, T.K.; Lowen, A.C. Genetic and Antigenic Characterization of an Expanding H3 Influenza A Virus Clade in U.S. Swine Visualized by Nextstrain. Msphere 2022, 7, e00994-21. [Google Scholar] [CrossRef]
- Zeller, M.A.; Chang, J.; Vincent, A.L.; Gauger, P.C.; Anderson, T.K. Spatial and temporal coevolution of N2 neuraminidase and H1 and H3 hemagglutinin genes of influenza A virus in US swine. Virus Evol. 2021, 7, veab090. [Google Scholar] [CrossRef]
- Trovao, N.S.; Nelson, M.I. When Pigs Fly: Pandemic influenza enters the 21st century. PLoS Pathog. 2020, 16, e1008259. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| 1A.1.1.3 Cons | A/OH/24/2017 CVV | A/sw/NC/A02245416/2020 | A/sw/TX/A02245420/2020 | 1A.3.3.2 Cons | A/ID/07/2018 | A/sw/UT/A02432386/2019 | 1A.3.3.3 Cons | A/OH/9/2015 CVV | A/sw/MN/A02245409/2020 | 1B.2.1 Cons | A/OH/35/2017 CVV | A/MI/383/2018 | A/sw/IL/A02139356/2018 | 1B.2.2.1 Cons | A/IA/32/2016 CVV | A/sw/IA/A02478968/2020 | 1B.2.2.2 Cons | A/sw/CO/A02245414/2020 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1A.1.1.3 consensus | 98.77 | 98.77 | 95.08 | 82.57 | 82.57 | 82.28 | 81.35 | 82.57 | 80.43 | 70.86 | 70.86 | 70.86 | 71.47 | 71.78 | 71.47 | 71.47 | 71.78 | 70.25 | |
| A/OH/24/2017 | 98.77 | 97.54 | 94.46 | 83.49 | 83.49 | 82.89 | 81.96 | 83.49 | 81.04 | 71.47 | 71.17 | 71.47 | 72.09 | 72.39 | 72.09 | 72.09 | 72.09 | 70.55 | |
| A/sw/NC/A02245416/2020 | 98.77 | 97.54 | 93.85 | 81.35 | 81.35 | 81.05 | 80.12 | 81.35 | 79.21 | 69.63 | 69.63 | 69.63 | 70.25 | 70.55 | 70.25 | 70.25 | 70.55 | 69.63 | |
| A/sw/TX/A02245420/2020 | 95.08 | 94.46 | 93.85 | 81.35 | 81.35 | 81.36 | 79.82 | 81.35 | 78.90 | 69.33 | 69.33 | 69.63 | 69.94 | 70.86 | 70.55 | 70.55 | 70.55 | 69.02 | |
| 1A.3.3.2 consensus | 82.57 | 83.49 | 81.35 | 81.35 | 99.69 | 97.26 | 87.77 | 86.85 | 86.54 | 70.03 | 70.34 | 70.64 | 70.64 | 70.95 | 70.03 | 70.64 | 70.95 | 69.42 | |
| A/ID/07/2018 | 82.57 | 83.49 | 81.35 | 81.35 | 99.69 | 97.57 | 88.07 | 87.16 | 86.85 | 70.03 | 70.34 | 70.64 | 70.64 | 70.95 | 70.03 | 70.64 | 70.95 | 69.42 | |
| A/sw/UT/A02432386/2019 | 82.28 | 82.89 | 81.05 | 81.36 | 97.26 | 97.57 | 88.70 | 86.56 | 87.48 | 70.35 | 70.66 | 70.96 | 70.96 | 71.27 | 70.35 | 70.96 | 70.66 | 69.13 | |
| 1A.3.3.3 consensus | 81.35 | 81.96 | 80.12 | 79.82 | 87.77 | 88.07 | 88.70 | 91.44 | 98.78 | 71.56 | 71.56 | 71.56 | 72.17 | 71.87 | 71.25 | 71.56 | 72.78 | 71.87 | |
| A/OH/9/2015 | 82.57 | 83.49 | 81.35 | 81.35 | 86.85 | 87.16 | 86.56 | 91.44 | 90.83 | 70.34 | 69.42 | 70.95 | 70.64 | 70.34 | 69.73 | 70.34 | 70.64 | 69.11 | |
| A/sw/MN/A02245409/2020 | 80.43 | 81.04 | 79.21 | 78.90 | 86.54 | 86.85 | 87.48 | 98.78 | 90.83 | 71.56 | 71.87 | 71.56 | 71.87 | 72.17 | 71.56 | 71.87 | 73.09 | 72.17 | |
| 1B.2.1 Consensus | 70.86 | 71.47 | 69.63 | 69.33 | 70.03 | 70.03 | 70.35 | 71.56 | 70.34 | 71.56 | 93.56 | 98.77 | 99.39 | 85.58 | 84.97 | 85.58 | 86.50 | 84.05 | |
| A/OH/35/2017 | 70.86 | 71.17 | 69.63 | 69.33 | 70.34 | 70.34 | 70.66 | 71.56 | 69.42 | 71.87 | 93.56 | 93.25 | 93.56 | 85.89 | 85.58 | 85.28 | 85.58 | 84.05 | |
| A/MI/383/2018 | 70.86 | 71.47 | 69.63 | 69.63 | 70.64 | 70.64 | 70.96 | 71.56 | 70.95 | 71.56 | 98.77 | 93.25 | 98.16 | 85.89 | 85.28 | 85.89 | 86.20 | 83.74 | |
| A/sw/IL/A02139356/2018 | 71.47 | 72.09 | 70.25 | 69.94 | 70.64 | 70.64 | 70.96 | 72.17 | 70.64 | 71.87 | 99.39 | 93.56 | 98.16 | 86.20 | 85.58 | 86.20 | 87.12 | 84.66 | |
| 1B.2.2.1 consensus | 71.78 | 72.39 | 70.55 | 70.86 | 70.95 | 70.95 | 71.27 | 71.87 | 70.34 | 72.17 | 85.58 | 85.89 | 85.89 | 86.20 | 98.47 | 98.16 | 91.72 | 88.65 | |
| A/IA/32/2016 | 71.47 | 72.09 | 70.25 | 70.55 | 70.03 | 70.03 | 70.35 | 71.25 | 69.73 | 71.56 | 84.97 | 85.58 | 85.28 | 85.58 | 98.47 | 96.63 | 91.72 | 88.65 | |
| A/sw/IA/A02478968/2020 | 71.47 | 72.09 | 70.25 | 70.55 | 70.64 | 70.64 | 70.96 | 71.56 | 70.34 | 71.87 | 85.58 | 85.28 | 85.89 | 86.20 | 98.16 | 96.63 | 90.49 | 87.42 | |
| 1B.2.2.2 consensus | 71.78 | 72.09 | 70.55 | 70.55 | 70.95 | 70.95 | 70.66 | 72.78 | 70.64 | 73.09 | 86.50 | 85.58 | 86.20 | 87.12 | 91.72 | 91.72 | 90.49 | 96.63 | |
| A/sw/CO/A02245414/2020 | 70.25 | 70.55 | 69.63 | 69.02 | 69.42 | 69.42 | 69.13 | 71.87 | 69.11 | 72.17 | 84.05 | 84.05 | 83.74 | 84.66 | 88.65 | 88.65 | 87.42 | 96.63 |
| Strain | Lineage | IDCDC-RG59 A/Ohio/24/2017-CVV | IDCDC-RG48 A/Ohio/9/2015-CVV | A/Idaho/7/2018 * |
|---|---|---|---|---|
| A/Ohio/24/2017 | 1A.1.1.3 | 1280 | <10 | 640 |
| A/swine/North Carolina/A02245416/2020 | 1A.1.1.3 | 80 | <10 | <10 |
| A/swine/Oklahoma/A02245237/2019 | 1A.2 | 20 | 320 | 640 |
| A/Ohio/9/2015 | 1A.3.3.3 | <10 | 2560 | <10 |
| A/swine/Minnesota/102245409/2020 | 1A.3.3.3 | <10 | 80 | 40 |
| A/Idaho/7/2018 * | 1A.3.3.2 | <10 | <10 | 1280 |
| A/swine/Utah/A02432386/2019 | 1A.3.3.2 | 40 | 160 | 2560 |
| Strain | Lineage | A/Iowa/32/2016 | A/Ohio/5/2017 | A/Michigan/383/2018 |
|---|---|---|---|---|
| A/Iowa/32/2016 | 1B.2.2.1 | 640 | 20 | 10 |
| A/swine/Iowa/A02478968/2020 | 1B.2.2.1 | 80 | 10 | 40 |
| A/swine/Colorado/A02245414/2020 | 1B.2.2.2 | 40 | 10 | 10 |
| A/Ohio/35/2017 | 1B.2.1 | 80 | 640 | 40 |
| A/Michigan/383/2018 | 1B.2.1 | 40 | 160 | 1280 |
| A/swine/Illinois/A02139356/2018 | 1B.2.1 | 20 | 320 | 1280 |
| 1 | Ferret Number | Change in Bodyweight (%) from 0–12 DPC | DPC with Nasal Shedding | Peak Nasal Titer * | 12 DPC HI Titer |
|---|---|---|---|---|---|
| 1A.1.1.3 | 1 | 4.61 | 5, 7 | 5.45 | 160 |
| 2 | 1.19 | 3, 5, 7 | 5.94 | 1280 | |
| 3 | 4.89 | 3, 5, 7 | 5.94 | 1280 | |
| 4 | 7.71 | 5 | 6.2 | 1280 | |
| 1A.3.3.3 | 5 | 21.36 | none | none | 80 |
| 6 | 16.83 | none | none | 160 | |
| 7 | 2.40 | 5, 7 | 5.87 | 320 | |
| 8 | −1.13 | 3, 5, 7 | 5.53 | 1280 | |
| 1B.2.2.1 | 9 | 6.74 | none | none | <10 |
| 10 | 9.84 | none | none | <10 | |
| 11 | 5.02 | 5, 7 | 4.95 | 160 | |
| 12 | 12.01 | none | none | <10 | |
| No virus | 13 | 7.25 | none | none | <10 |
| 14 | 2.35 | none | none | <10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimble, J.B.; Souza, C.K.; Anderson, T.K.; Arendsee, Z.W.; Hufnagel, D.E.; Young, K.M.; Lewis, N.S.; Davis, C.T.; Thor, S.; Vincent Baker, A.L. Interspecies Transmission from Pigs to Ferrets of Antigenically Distinct Swine H1 Influenza A Viruses with Reduced Reactivity to Candidate Vaccine Virus Antisera as Measures of Relative Zoonotic Risk. Viruses 2022, 14, 2398. https://doi.org/10.3390/v14112398
Kimble JB, Souza CK, Anderson TK, Arendsee ZW, Hufnagel DE, Young KM, Lewis NS, Davis CT, Thor S, Vincent Baker AL. Interspecies Transmission from Pigs to Ferrets of Antigenically Distinct Swine H1 Influenza A Viruses with Reduced Reactivity to Candidate Vaccine Virus Antisera as Measures of Relative Zoonotic Risk. Viruses. 2022; 14(11):2398. https://doi.org/10.3390/v14112398
Chicago/Turabian StyleKimble, J. Brian, Carine K. Souza, Tavis K. Anderson, Zebulun W. Arendsee, David E. Hufnagel, Katharine M. Young, Nicola S. Lewis, C. Todd Davis, Sharmi Thor, and Amy L. Vincent Baker. 2022. "Interspecies Transmission from Pigs to Ferrets of Antigenically Distinct Swine H1 Influenza A Viruses with Reduced Reactivity to Candidate Vaccine Virus Antisera as Measures of Relative Zoonotic Risk" Viruses 14, no. 11: 2398. https://doi.org/10.3390/v14112398
APA StyleKimble, J. B., Souza, C. K., Anderson, T. K., Arendsee, Z. W., Hufnagel, D. E., Young, K. M., Lewis, N. S., Davis, C. T., Thor, S., & Vincent Baker, A. L. (2022). Interspecies Transmission from Pigs to Ferrets of Antigenically Distinct Swine H1 Influenza A Viruses with Reduced Reactivity to Candidate Vaccine Virus Antisera as Measures of Relative Zoonotic Risk. Viruses, 14(11), 2398. https://doi.org/10.3390/v14112398