

Seroprevalence of Getah virus in Pigs in Eastern China Determined with a Recombinant E2 Protein-Based Indirect ELISA


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Serum Samples
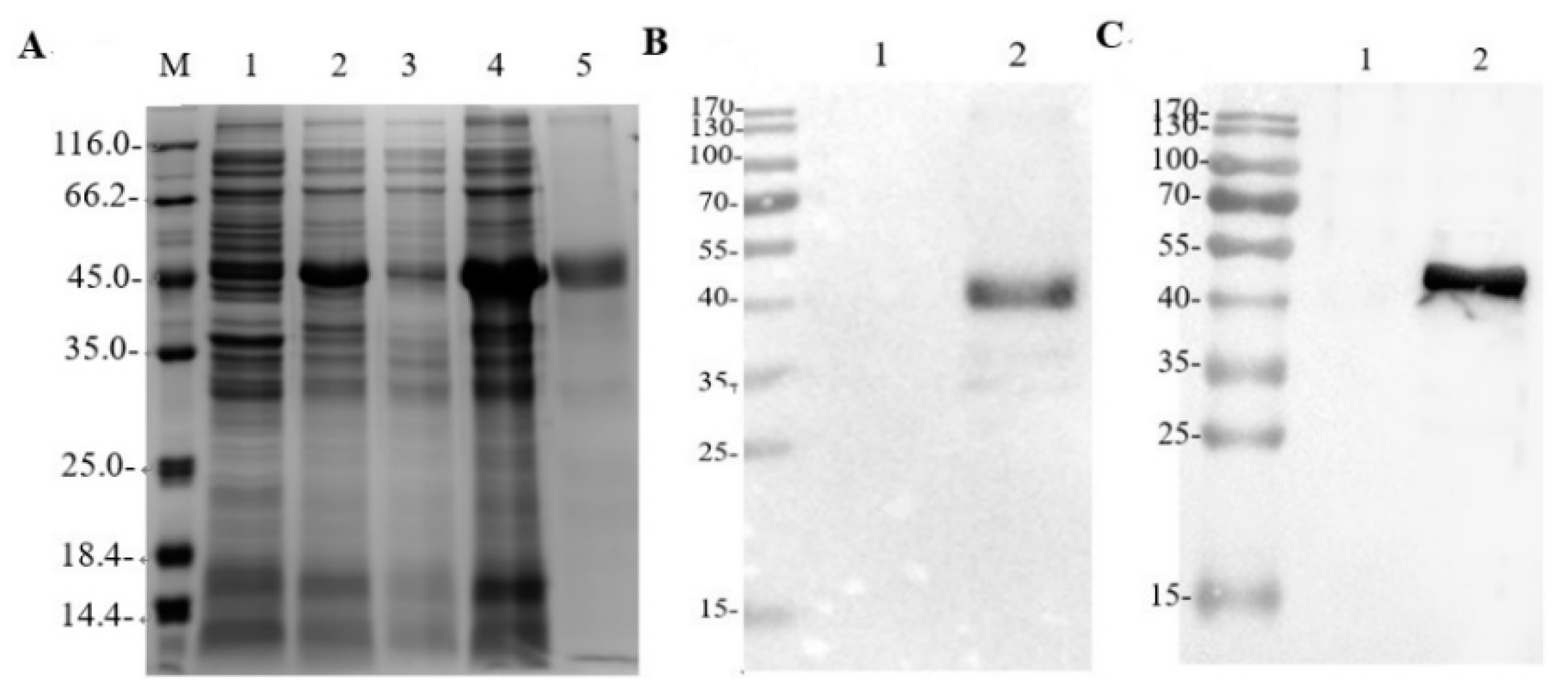
2.2. Expression and Purification of rE2 Protein
2.3. ELISA Development
2.4. Specificity Analysis
2.5. Sensitivity Analysis
2.6. Reproducibility Assay for the Indirect ELISA
2.7. Coincidence Test of the Indirect ELISA
2.8. Statistical Analysis
3. Results
3.1. Expression and Purification of rE2 Protein
3.2. ELISA Optimization with rE2 Protein
3.3. ROC Curve Analysis
3.4. Specificity Analysis of the Indirect ELISA
3.5. Sensitivity Analysis Test of the Indirect ELISA
3.6. Reproducibility of the Indirect ELISA
3.7. Coincidence of the Indirect ELISA
3.8. Relationship between Swine GETV-Antibody-Positive Rate and Province in Eastern China
3.9. Relationship between Swine GETV-Antibody-Positive Rate and Season in Eastern China
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fukunaga, Y.; Kumanomido, T.; Kamada, M. Getah virus as an equine pathogen. Vet. Clin. N. Am. Equine Pract. 2000, 16, 605–617. [Google Scholar] [CrossRef]
- Feng, Y.; Fu, S.; Zhang, H.; Li, M.; Zhou, T.; Wang, J.; Liang, G. Distribution of mosquitoes and mosquito-borne viruses along the ChinaMyanmar border in Yunnan Province. Jpn. J. Infect. Dis. 2012, 65, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Guryev, Y.L.; Gromashevsky, V.L.; Prilipov, A.G.; Lvov, S.D. Analysis of the genome of two Getah virus strains (LEIV 16275 Mar and LEIV 17741 MPR) isolated from mosquitoes in the North-Eastern Asia. Vopr. Virusol. 2008, 53, 27–31. [Google Scholar]
- Li, Y.; Liu, H.; Fu, S.H.; Li, X.L.; Guo, X.F.; Li, M.H.; Liang, G.D. From discovery to spread: The evolution and phylogeny of Getah virus. Infect. Genet. Evol. 2017, 55, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, M.; Banna, H.; Tsujimura, K.; Kobayashi, M.; Kikuchi, T.; Yamanaka, T.; Kondo, T. Getah virus infection among racehorses, Japan. Emerg. Infect. Dis. 2015, 21, 883–885. [Google Scholar] [CrossRef]
- Seo, H.J.; Kim, H.C.; Klein, T.A.; Park, J.Y.; Cho, Y.S.; Cho, I.S.; Yeh, J.Y. Characterization of recent Getah virus isolates from South Korea. Acta Virol. 2012, 56, 265–267. [Google Scholar] [CrossRef][Green Version]
- Rattanatumhi, K.; Prasertsincharoen, N.; Naimon, N.; Kuwata, R.; Shimoda, H.; Ishijima, K.; Yonemitsu, K.; Minami, S.; Kuroda, Y.; Tatemoto, K.; et al. A serological survey and characterization of Getah virus in domestic pigs in Thailand, 2017–2018. Transbound. Emerg. Dis. 2022, 69, 913–918. [Google Scholar] [CrossRef]
- Broen, C.; Timoney, P. Getah virus infection of Indian horses. Trop. Anim. Health Prod. 1988, 30, 241–252. [Google Scholar] [CrossRef]
- Bannai, H.; Ochi, A.; Nemoto, M.; Tsujimura, K.; Yamanaka, T.; Kondo, T. A 2015 outbreak of Getah virus infection occurring among Japanese racehorses sequentially to an outbreak in 2014 at the same site. BMC Vet. Res. 2016, 12, 98. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Turtle, L.; Solomon, T. Japanese encephalitis virus infection. Handb. Clin. Neurol. 2014, 123, 561–576. [Google Scholar]
- Kumanomido, T.; Wada, R.; Kanemaru, T.; Kamada, M.; Hirasawa, K.; Akiyama, Y. Clinical and virological observations on swine experimentally infected with Getah virus. Vet. Microbiol. 1988, 16, 295–301. [Google Scholar] [CrossRef]
- Yang, T.; Li, R.; Hu, Y.; Yang, L.; Zhao, D.; Du, L.; Li, J.; Ge, M.; Yu, X. An outbreak of Getah virus infection among pigs in China. Transbound. Emerg. Dis. 2018, 65, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, S.; Guo, X.; Li, X.; Li, M.; Wang, L.; Liang, G. Serological survey of Getah virus in domestic animals in Yunnan province, China. Vector-Borne Zoonotic Dis. 2019, 19, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Mukhopadhyay, S.; Merits, A.; Bolling, B.; Nasar, F.; Coffey, L.L.; Powers, A.; Weaver, S.C. ICTV virus taxonomy profile: Togaviridae. J. Gen. Virol. 2018, 99, 761–762. [Google Scholar] [CrossRef] [PubMed]
- Fong, R.H.; Banik, S.S.R.; Mattia, K.; Barnes, T.; Tucker, D.; Liss, N.; Lu, K.; Selvarajar, S.; Srinivasan, S.; Mabila, M.; et al. Exposure of epitope residues on the outer face of the Chikungunya virus envelope trimer determines antibody neutralizing efficacy. J. Virol. 2014, 88, 14364–14379. [Google Scholar] [CrossRef] [PubMed]
- Weger-Lucarelli, J.; Aliota, M.T.; Wlodarchak, N.; Kamlangdee, A.; Swanson, R.; Osorio, J.E. Dissecting the role of E2 protein domains in Alphavirus pathogenicity. J. Virol. 2016, 90, 2418–2433. [Google Scholar] [CrossRef]
- Wang, N.; Zhai, X.; Li, X.; Wang, Y.; He, W.; Jiang, Z.; Veit, M.; Su, S. Attenuation of Getah virus by a single amino acid substitution at residue 253 of the E2 protein that might be part of a new heparan sulfate binding site on Alphaviruses. J. Virol. 2022, 96, e01751-21. [Google Scholar] [CrossRef]
- Bannai, H.; Nemoto, M.; Tsujimura, K.; Yamanaka, T.; Kokado, H. Development of an enzyme-linked immunosorbent assay for Getah virus infection in horses using recombinant E2 protein as an antigen. J. Virol. Methods 2019, 271, 113681. [Google Scholar] [CrossRef]
- Bannai, H.; Nemoto, M.; Tsujimura, K.; Ohta, M. Establishment of an enzyme-linked immunosorbent assay for Getah virus infection in horses using a 20-mer synthetic peptide for the E2 glycoprotein as an antigen. Arch. Virol. 2020, 165, 377–385. [Google Scholar] [CrossRef]
- Hameed, M.; Wahaab, A.; Shan, T.; Wang, X.; Khan, S.; Di, D.; Xiqian, L.; Zhang, J.J.; Anwar, M.N.; Nawaz, M.; et al. A Metagenomic Analysis of Mosquito Virome Collected from Different Animal Farms at Yunnan-Myanmar Border of China. Front. Microbiol. 2021, 11, 591478. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Guan, Z.; Yang, Y.; Zhang, J.; Sun, Q.; Li, B.; Qiu, Y.; Liu, K.; Shao, D.; et al. Rapid Differential Detection of Japanese Encephalitis Virus and Getah Virus in Pigs or Mosquitos by a Duplex TaqMan Real-Time RT-PCR Assay. Front. Vet. Sci. 2022, 9, 839443. [Google Scholar] [CrossRef]
- Shan, Y.; Liu, Y.; Liu, Z.; Li, G.; Chen, C.; Luo, H.; Chen, Y.; Guo, N.; Shi, X.; Zhang, X.; et al. Development and application of an indirect enzyme-linked immunosorbent assay using recombinant S1 for serological testing of porcine epidemic diarrhea virus. Can. J. Microbiol. 2019, 65, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Peng, J.Y.; Cheng, Y.H.; Chang, Y.C.; Wu, Y.T.; Tsai, P.S.; Chiou, H.Y.; Jeng, C.R.; Chang, H.W. Development and comparison of enzyme-linked immunosorbent assays based on recombinant trimeric full-length and truncated spike proteins for detecting antibodies against porcine epidemic diarrhea virus. BMC Vet. Res. 2019, 15, 421. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Shin, M.K.; Cha, S.B.; Yoo, H.S. Development of a novel enzyme-linked immunosorbent assay to detect anti-IgG against swine hepatitis E virus. J. Vet. Sci. 2013, 14, 467–472. [Google Scholar] [CrossRef]
- Xing, C.; Jiang, J.; Lu, Z.; Mi, S.; He, B.; Tu, C.; Liu, X.; Gong, W. Isolation and characterization of Getah virus from pigs in Guangdong province of China. Transbound. Emerg. Dis. 2020, 67, 2253–2279. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jose, J.; Xiang, Y.; Kuhn, R.; Rossmann, M. Structural changes of envelope proteins during alphavirus fusion. Nature. 2010, 468, 705–708. [Google Scholar] [CrossRef]
- Voss, J.; Vaney, M.; Duquerroy, S.; Vonrhein, C.; Girard-Blanc, C.; Crublet, E.; Thompson, A.; Bricogne, G.; Rey, F. Glycoprotein organization of Chikungunya virus particles revealed by X-ray crystallography. Nature 2010, 468, 709–712. [Google Scholar] [CrossRef]
- Cho, B.; Jeon, B.; Kim, J.; Noh, J.; Kim, J.; Park, M.; Park, S. Expression and evaluation of Chikungunya virus E1 and E2 envelope proteins for serodiagnosis of Chikungunya virus infection. Yonsei Med. J. 2008, 49, 828–835. [Google Scholar] [CrossRef]
- Verma, P.; Bhatnagar, S.; Kumar, P.; Chattree, V.; Parida, M.; Hoti, S.; Ali, S.; Rao, D. Analysis of antibody response (IgM, IgG, IgG3) to Chikungunya virus using a panel of peptides derived from envelope protein for serodiagnosis. Clin. Chem. Lab. Med. 2014, 52, 297–307. [Google Scholar] [PubMed]
- Qian, S.; Jia, X.; Gao, Z.; Weida, Z.; Xu, Q.; Li, Z. Isolation and identification of porcine deltacoronavirus and alteration of immunoglobulin transport receptors in the intestinal mucosa of PDCoV-infected piglets. Viruses 2020, 12, 79–84. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Serum Dilution | Protein Coating Concentration (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| 196.000 | 98.000 | 49.000 | 24.500 * | 12.250 | 6.130 | 3.030 | ||
| 1:25 | P | 2.871 ± 0.021 | 2.870 ± 0.028 | 2.651 ± 0.019 | 2.885 ± 0.061 | 2.716 ± 0.068 | 2.823 ± 0.027 | 2.863 ± 0.068 |
| N | 0.279 ± 0.006 | 0.287 ± 0.003 | 0.273 ± 0.004 | 0.302 ± 0.008 | 0.288 ± 0.002 | 0.379 ± 0.004 | 0.463 ± 0.006 | |
| P/N | 10.290 | 10.004 | 9.710 | 9.568 | 9.418 | 7.456 | 6.189 | |
| 1:50 | P | 2.546 ± 0.043 | 2.432 ± 0.033 | 2.111 ± 0.011 | 2.064 ± 0.159 | 1.886 ± 0.086 | 1.774 ± 0.023 | 1.712 ± 0.024 |
| N | 0.173 ± 0.005 | 0.175 ± 0.004 | 0.181 ± 0.003 | 0.183 ± 0.003 | 0.194 ± 0.001 | 0.198 ± 0.002 | 0.241 ± 0.003 | |
| P/N | 14.717 | 13.897 | 11.663 | 11.279 | 9.722 | 8.966 | 7.106 | |
| 1:100 * | P | 1.871 ± 0.069 | 1.870 ± 0.034 | 1.651 ± 0.047 | 1.885 ± 0.031 | 1.716 ± 0.032 | 1.823±0.057 | 1.863 ± 0.022 |
| N | 0.086 ± 0.012 | 0.091 ± 0.003 | 0.087 ± 0.005 | 0.073 ± 0.001 | 0.098 ± 0.011 | 0.130 ± 0.009 | 0.143 ± 0.002 | |
| P/N | 21.684 | 20.605 | 19.013 | 25.822 * | 17.579 | 14.043 | 13.000 | |
| 1:200 | P | 1.704 ± 0.064 | 1.741 ± 0.032 | 1.555 ± 0.052 | 1.726 ± 0.017 | 1.579 ± 0.056 | 1.774 ± 0.024 | 1.725 ± 0.011 |
| N | 0.083 ± 0.012 | 0.088 ± 0.010 | 0.082 ± 0.009 | 0.108 ± 0.004 | 0.111 ± 0.006 | 0.144 ± 0.007 | 0.142 ± 0.005 | |
| P/N | 20.421 | 19.871 | 18.868 | 16.014 | 14.221 | 12.343 | 12.115 | |
| Sample Number | Interassay CV (%) | Intra-Assay CV(%) | ||
|---|---|---|---|---|
| X ± SD | CV(%) | X ± SD | CV(%) | |
| 1 | 1.147 ± 0.038 | 3.29% | 1.140 ± 0.026 | 2.25% |
| 2 | 0.556 ± 0.032 | 5.78% | 0.554 ± 0.033 | 5.94% |
| 3 | 0.775 ± 0.029 | 3.79% | 0.785 ± 0.048 | 6.16% |
| 4 | 0.852 ± 0.043 | 5.06% | 0.857 ± 0.050 | 5.84% |
| 5 | 0.471 ± 0.019 | 3.99% | 0.471 ± 0.017 | 3.66% |
| 6 | 1.203 ± 0.070 | 5.81% | 1.195 ± 0.060 | 5.05% |
| IFA Results | ||||
|---|---|---|---|---|
| Positive | Negative | Total | ||
| Indirect ELISA results | Positive | 47 | 4 | 51 |
| Negative | 3 | 79 | 82 | |
| Total | 50 | 83 | 133 | |
| Variable | Category | No. Examined | No. Positive | Positive Rate | p-Value |
|---|---|---|---|---|---|
| Provinces | Shandong | 20 | 6 | 30.00% | 0.24 |
| Hebei | 20 | 7 | 35.00% | 0.24 | |
| Zhejiang | 20 | 9 | 45.00% | 0.19 | |
| Shanghai | 20 | 7 | 35.00% | 0.86 | |
| Jiangsu | 23 | 10 | 43.48% | 0.73 | |
| Guangdong | 30 | 11 | 36.67% | 0.43 | |
| Season | Spring | 66 | 16 | 24.24% | 0.03 |
| Autumn | 67 | 34 | 50.75% | 0.04 | |
| Total | 133 | 50 | 37.59% | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Q.; Xie, Y.; Guan, Z.; Zhang, Y.; Li, Y.; Yang, Y.; Zhang, J.; Li, Z.; Qiu, Y.; Li, B.; et al. Seroprevalence of Getah virus in Pigs in Eastern China Determined with a Recombinant E2 Protein-Based Indirect ELISA. Viruses 2022, 14, 2173. https://doi.org/10.3390/v14102173
Sun Q, Xie Y, Guan Z, Zhang Y, Li Y, Yang Y, Zhang J, Li Z, Qiu Y, Li B, et al. Seroprevalence of Getah virus in Pigs in Eastern China Determined with a Recombinant E2 Protein-Based Indirect ELISA. Viruses. 2022; 14(10):2173. https://doi.org/10.3390/v14102173
Chicago/Turabian StyleSun, Qing, Yixuan Xie, Zhixin Guan, Yan Zhang, Yuhao Li, Yang Yang, Junjie Zhang, Zongjie Li, Yafeng Qiu, Beibei Li, and et al. 2022. "Seroprevalence of Getah virus in Pigs in Eastern China Determined with a Recombinant E2 Protein-Based Indirect ELISA" Viruses 14, no. 10: 2173. https://doi.org/10.3390/v14102173
APA StyleSun, Q., Xie, Y., Guan, Z., Zhang, Y., Li, Y., Yang, Y., Zhang, J., Li, Z., Qiu, Y., Li, B., Liu, K., Shao, D., Wang, J., Ma, Z., Wei, J., & Li, P. (2022). Seroprevalence of Getah virus in Pigs in Eastern China Determined with a Recombinant E2 Protein-Based Indirect ELISA. Viruses, 14(10), 2173. https://doi.org/10.3390/v14102173