
Characterization of the Subclinical Infection of Porcine Deltacoronavirus in Grower Pigs under Experimental Conditions
 , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. The Study
2.2. Sample Collection, Processing, and Storage
2.3. Sample Processing for Assessment of virus Infectivity in Cell Culture
2.4. Inoculation of Fecal and Oral Fluid Specimens in Cell Culture
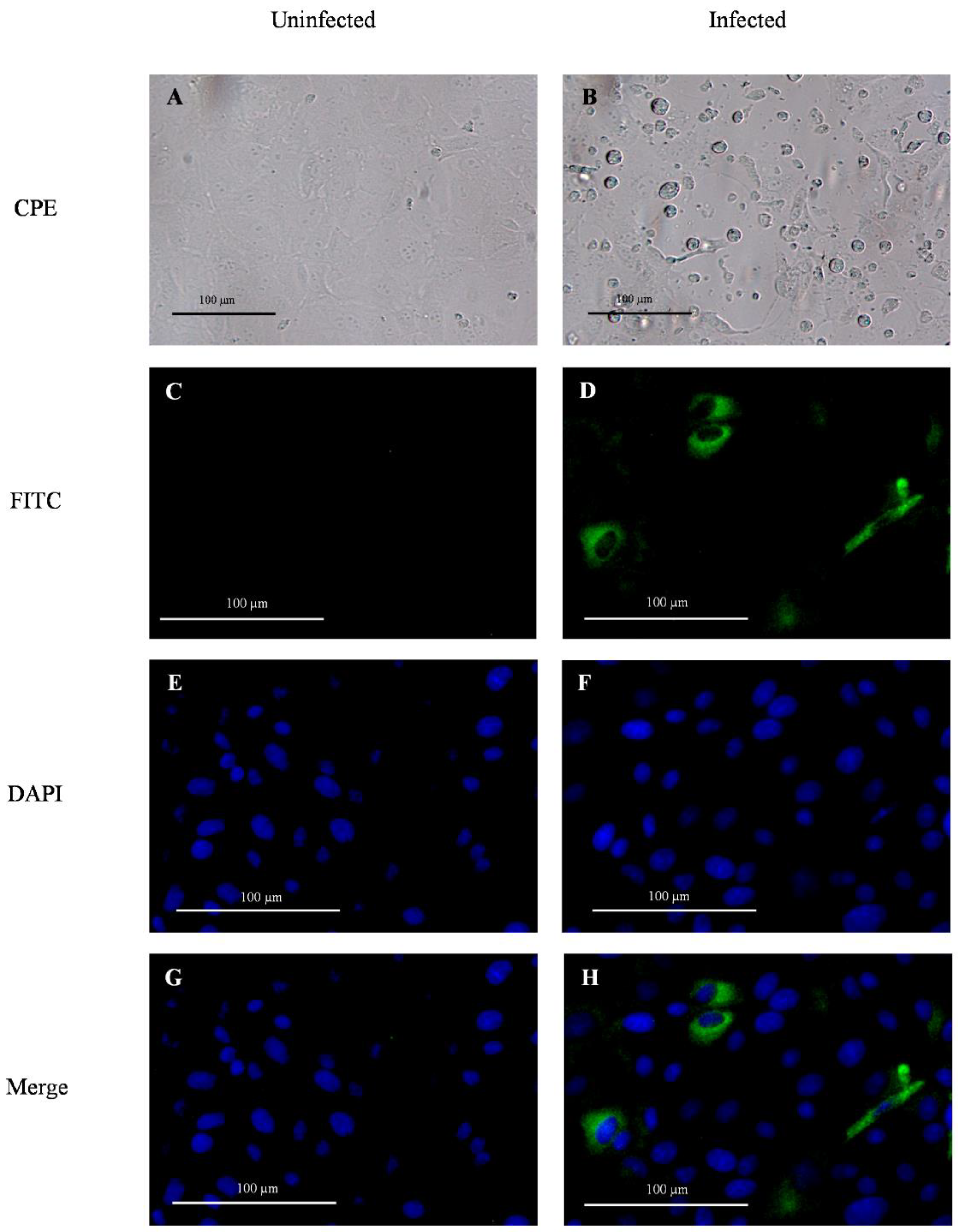
2.5. Immunofluorescence Assay
2.6. Quantitative Reverse Transcription PCR
2.7. Enzyme-Linked Immunosorbent Assay
2.8. Multiplex Porcine Cytokine and Chemokine Immunoassay
2.9. Data Analysis
3. Results
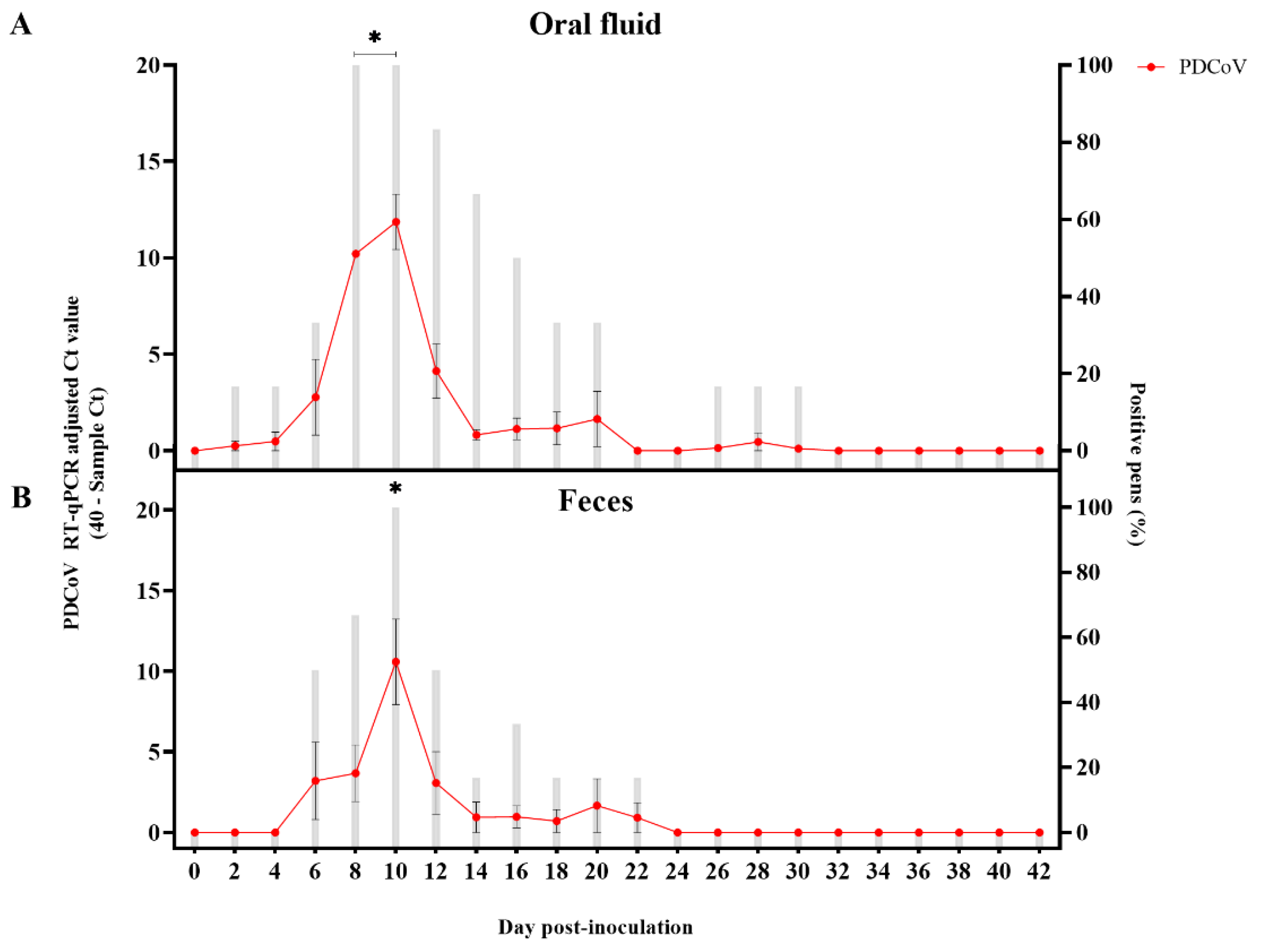
3.1. Absence of Noticeable Clinical Signs and Viremia after PDCoV Inoculation but Active Virus Replication and Shedding Detected in Feces and Oral Fluids
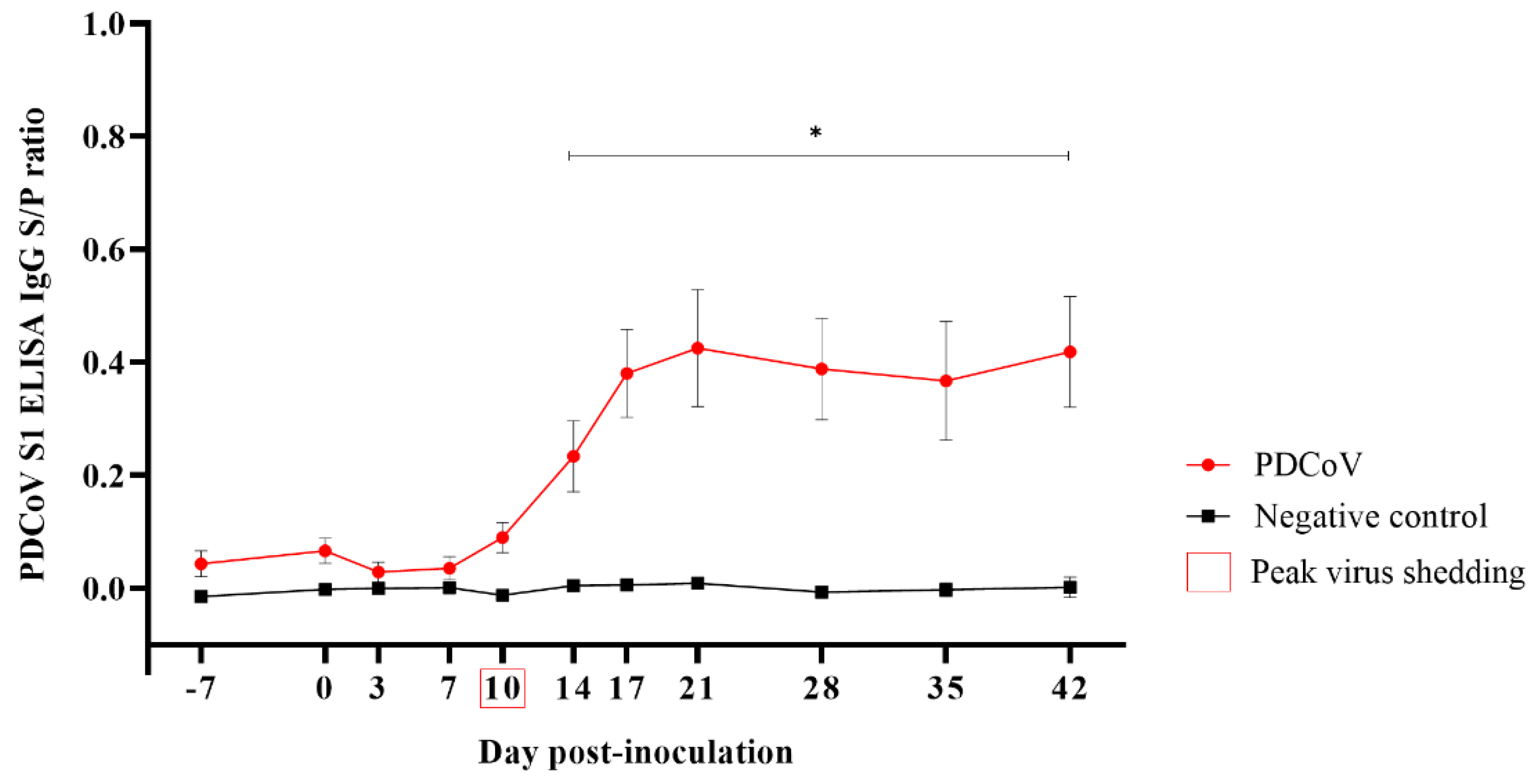
3.2. Specific IgG Seroconversion Detected in Serum Two Weeks after PDCoV Inoculation
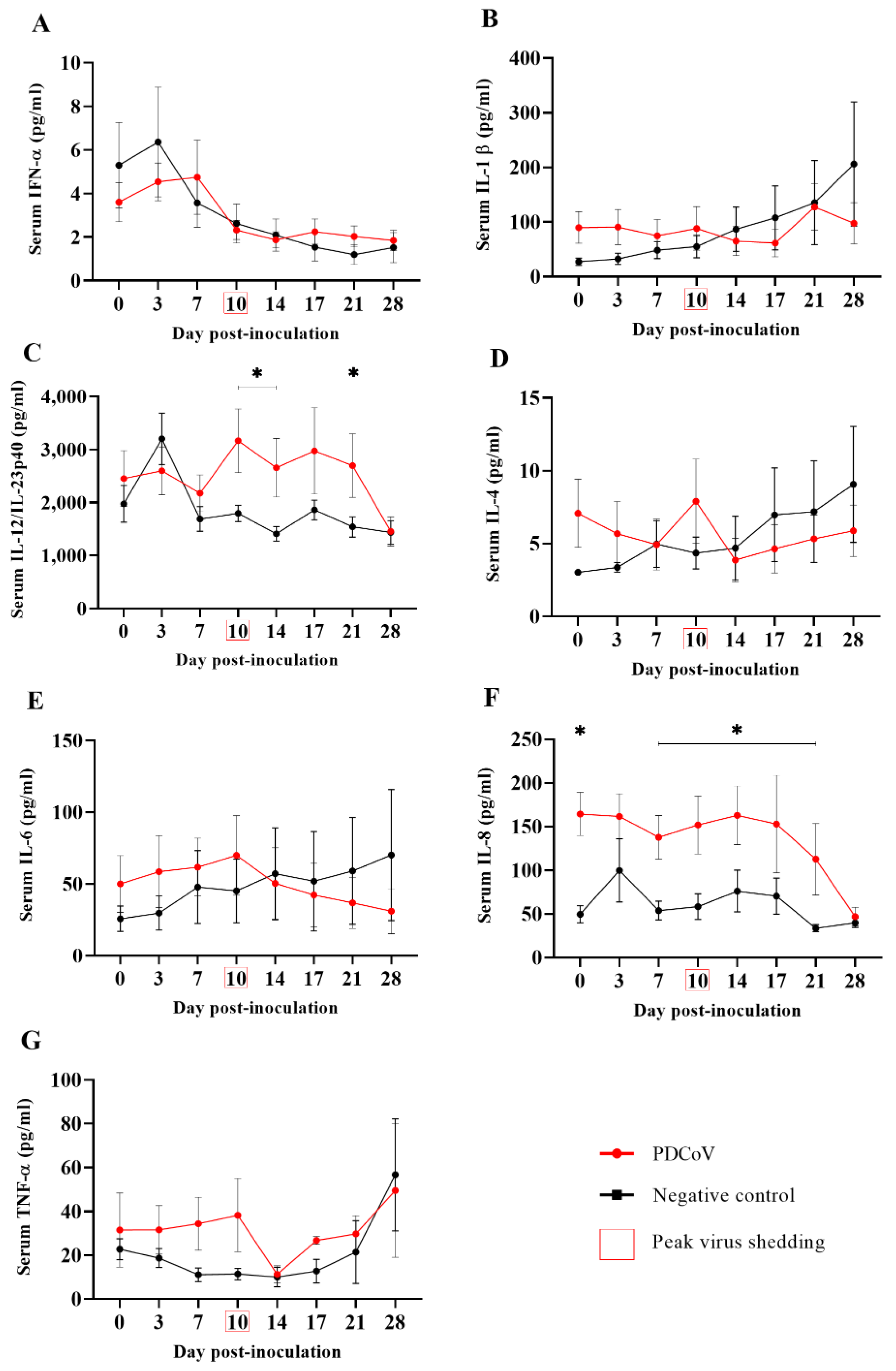
3.3. Increased Serum Levels of Proinflammatory Cytokine IL-12 in Response to PDCoV Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taxonomy History—Taxonomy—ICTV. Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=202101870 (accessed on 2 June 2022).
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Huang, Y.; Lau, S.K.P.; Yuen, K.Y. Coronavirus Genomics and Bioinformatics Analysis. Viruses 2010, 2, 1804–1820. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.W.; Leung, C.Y.H.; Gilbert, M.; Joyner, P.H.; Ng, E.M.; Tse, T.M.; Guan, Y.; Peiris, J.S.M.; Poon, L.L.M. Avian Coronavirus in Wild Aquatic Birds. J. Virol. 2011, 85, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, T.; Dara, R.; Misener, M.; Pasma, T.; Moser, L.; Poljak, Z. Herd-level Prevalence and Incidence of Porcine Epidemic Diarrhoea Virus (PEDV) and Porcine Deltacoronavirus (PDCoV) in Swine Herds in Ontario, Canada. Transbound Emerg. Dis. 2018, 65, 1197. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rivera, C.; Ramírez-Mendoza, H.; Mendoza-Elvira, S.; Segura-Velázquez, R.; Sánchez-Betancourt, J.I. First Report and Phylogenetic Analysis of Porcine Deltacoronavirus in Mexico. Transbound Emerg. Dis. 2019, 66, 1436. [Google Scholar] [CrossRef]
- Vicente-Huaman, J.; Gómez-Quispe, O.E. Evaluation of a Porcine Deltacoronavirus Eradication Program in a Full-Cycle Pig Farm in Peru. J. Adv. Vet. Anim. Res. 2021, 8, 300–306. [Google Scholar] [CrossRef]
- Wang, L.; Byrum, B.; Zhang, Y. Detection and Genetic Characterization of Deltacoronavirus in Pigs, Ohio, USA, 2014. Emerg. Infect. Dis. 2014, 20, 1227. [Google Scholar] [CrossRef]
- Song, D.; Zhou, X.; Peng, Q.; Chen, Y.; Zhang, F.; Huang, T.; Zhang, T.; Li, A.; Huang, D.; Wu, Q.; et al. Newly Emerged Porcine Deltacoronavirus Associated with Diarrhoea in Swine in China: Identification, Prevalence and Full-Length Genome Sequence Analysis. Transbound Emerg. Dis. 2015, 62, 575–580. [Google Scholar] [CrossRef]
- Zhang, F.; Luo, S.; Gu, J.; Li, Z.; Li, K.; Yuan, W.; Ye, Y.; Li, H.; Ding, Z.; Song, D.; et al. Prevalence and Phylogenetic Analysis of Porcine Diarrhea Associated Viruses in Southern China from 2012 to 2018. Bmc Vet. Res. 2019, 15, 1–9. [Google Scholar] [CrossRef]
- Zhai, S.L.; Wei, W.K.; Li, X.P.; Wen, X.H.; Zhou, X.; Zhang, H.; Lv, D.H.; Li, F.; Wang, D. Occurrence and Sequence Analysis of Porcine Deltacoronaviruses in Southern China Susanna Lau. Virol. J. 2016, 13, 1–11. [Google Scholar] [CrossRef]
- Lee, S.; Lee, C. Complete Genome Characterization of Korean Porcine Deltacoronavirus Strain KOR/KNU14-04/2014. Genome Announc. 2014, 2, e01191-14. [Google Scholar] [CrossRef] [PubMed]
- Saeng-chuto, K.; Lorsirigool, A.; Temeeyasen, G.; Vui, D.T.; Stott, C.J.; Madapong, A.; Tripipat, T.; Wegner, M.; Intrakamhaeng, M.; Chongcharoen, W.; et al. Different Lineage of Porcine Deltacoronavirus in Thailand, Vietnam and Lao PDR in 2015. Transbound Emerg. Dis. 2017, 64, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Shibahara, T.; Imai, N.; Yamamoto, T.; Ohashi, S. Genetic Characterization and Pathogenicity of Japanese Porcine Deltacoronavirus. Infect. Genet. Evol. 2018, 61, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.; Hutchings, L. A Transmissible Gastroenteritis in Pigs. J. Am. Vet. Med. Assoc. 1946, 108, 257–259. [Google Scholar]
- Coussement, W.; Ducatelle, R.; Debouck, P.; Hoorens, J. Pathology of Experimental CV777 Coronavirus Enteritis in Piglets. I. Histological and Histochemical Study. Vet. Pathol. 1982, 19, 46–56. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Liang, X.; Lou, F.; Oglesbee, M.; Krakowka, S.; Li, J. Origin, Evolution, and Virulence of Porcine Deltacoronaviruses in the United States. mBio 2015, 6, e00064-15. [Google Scholar] [CrossRef]
- Trapp, A.L.; Sanger, V.L.; Stalnaker, E. Lesions of the Small Intestinal Mucosa in Transmissible Gastroenteritis-Infected Germfree Pigs. Am. J. Vet. Res. 1966, 27, 1695–1702. [Google Scholar]
- Stevenson, G.W.; Hoang, H.; Schwartz, K.J.; Burrough, E.R.; Sun, D.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; et al. Emergence of Porcine Epidemic Diarrhea Virus in the United States: Clinical Signs, Lesions, and Viral Genomic Sequences. J. Vet. Diagn. Investig. 2013, 25, 649–654. [Google Scholar] [CrossRef]
- Vitosh-Sillman, S.; Loy, J.D.; Brodersen, B.; Kelling, C.; Doster, A.; Topliff, C.; Nelson, E.; Bai, J.; Schirtzinger, E.; Poulsen, E.; et al. Experimental Infection of Conventional Nursing Pigs and Their Dams with Porcine Deltacoronavirus. J. Vet. Diagn. Investig. 2016, 28, 486–497. [Google Scholar] [CrossRef]
- Chen, Q.; Gauger, P.; Stafne, M.; Thomas, J.; Arruda, P.; Burrough, E.; Madson, D.; Brodie, J.; Magstadt, D.; Derscheid, R.; et al. Pathogenicity and Pathogenesis of a United States Porcine Deltacoronavirus Cell Culture Isolate in 5-Day-Old Neonatal Piglets. Virology 2015, 482, 51–59. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Liu, Y.; Da, S.; Shi, H.; Zhang, X.; Liu, J.; Cao, L.; Zhu, X.; Wang, X.; et al. Pathogenicity of Porcine Deltacoronavirus (PDCoV) Strain NH and Immunization of Pregnant Sows with an Inactivated PDCoV Vaccine Protects 5-day-old Neonatal Piglets from Virulent Challenge. Transbound Emerg. Dis. 2020, 67, 572. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Hu, H.; Eyerly, B.; Lu, Z.; Chepngeno, J.; Saif, L.J. Pathogenicity of 2 Porcine Deltacoronavirus Strains in Gnotobiotic Pigs. Emerg. Infect. Dis. 2015, 21, 650. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hayes, J.; Sarver, C.; Byrum, B.; Zhang, Y. Porcine Deltacoronavirus: Histological Lesions and Genetic Characterization. Arch. Virol. 2016, 161, 171. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Raymond, L.; Jiang, Y.; Collins, J.; Rossow, K.; Rovira, A. Rapid Detection, Complete Genome Sequencing, and Phylogenetic Analysis of Porcine Deltacoronavirus. Emerg. Infect. Dis. 2014, 20, 1347. [Google Scholar] [CrossRef]
- Li, G.; Chen, Q.; Harmon, K.M.; Yoon, K.J.; Schwartz, K.J.; Hoogland, M.J.; Gauger, P.C.; Main, R.G.; Zhang, J. Full-Length Genome Sequence of Porcine Deltacoronavirus Strain USA/IA/2014/8734. Genome Announc. 2014, 2, 278–292. [Google Scholar] [CrossRef] [PubMed]
- Thachil, A.; Gerber, P.F.; Xiao, C.T.; Huang, Y.W.; Opriessnig, T. Development and Application of an ELISA for the Detection of Porcine Deltacoronavirus IgG Antibodies. PLoS ONE 2015, 10, e0124363. [Google Scholar] [CrossRef]
- Zhang, J. Porcine Deltacoronavirus: Overview of Infection Dynamics, Diagnostic Methods, Prevalence and Genetic Evolution. Virus Res. 2016, 226, 71–84. [Google Scholar] [CrossRef]
- Hu, H.; Jung, K.; Vlasova, A.N.; Saif, L.J. Experimental Infection of Gnotobiotic Pigs with the Cell-Culture-Adapted Porcine Deltacoronavirus Strain OH-FD22. Arch. Virol. 2016, 161, 3421–3434. [Google Scholar] [CrossRef]
- Gimenez-Lirola, L.G.; Zhang, J.; Carrillo-Avila, J.A.; Chen, Q.; Magtoto, R.; Poonsuk, K.; Baum, D.H.; Piñeyro, P.; Zimmerman, J. Reactivity of Porcine Epidemic Diarrhea Virus Structural Proteins to Antibodies against Porcine Enteric Coronaviruses: Diagnostic Implications. J. Clin. Microbiol. 2017, 55, 1426. [Google Scholar] [CrossRef]
- Prickett, J.; Simer, R.; Christopher-Hennings, J.; Yoon, K.J.; Evans, R.; Zimmerman, J. Detection of Porcine Reproductive and Respiratory Syndrome Virus Infection in Porcine Oral Fluid Samples: A Longitudinal Study under Experimental Conditions. J. Vet. Diagn. Investig. 2008, 20, 156–163. [Google Scholar] [CrossRef]
- Magtoto, R.; Poonsuk, K.; Baum, D.; Zhang, J.; Chen, Q.; Ji, J.; Piñeyro, P.; Zimmerman, J.; Giménez-Lirola, L.G. Evaluation of the Serologic Cross-Reactivity between Transmissible Gastroenteritis Coronavirus and Porcine Respiratory Coronavirus Using Commercial Blocking Enzyme-Linked Immunosorbent Assay Kits. mSphere 2019, 4, e00017-19. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Jung, K.; Vlasova, A.N.; Chepngeno, J.; Lu, Z.; Wang, Q.; Saif, L.J. Isolation and Characterization of Porcine Deltacoronavirus from Pigs with Diarrhea in the United States. J. Clin. Microbiol. 2015, 53, 1537–1548. [Google Scholar] [CrossRef]
- Xu, Z.; Zhong, H.; Huang, S.; Zhou, Q.; Du, Y.; Chen, L.; Xue, C.; Cao, Y. Porcine Deltacoronavirus Induces TLR3, IL-12, IFN-α, IFN-β and PKR MRNA Expression in Infected Peyer’s Patches in Vivo. Vet. Microbiol. 2019, 228, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Miyazaki, A.; Hu, H.; Saif, L.J. Susceptibility of Porcine IPEC-J2 Intestinal Epithelial Cells to Infection with Porcine Deltacoronavirus (PDCoV) and Serum Cytokine Responses of Gnotobiotic Pigs to Acute Infection with IPEC-J2 Cell Culture-Passaged PDCoV. Vet. Microbiol. 2018, 221, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Saeng-chuto, K.; Madapong, A.; Kaeoket, K.; Piñeyro, P.E.; Tantituvanont, A.; Nilubol, D. Coinfection of Porcine Deltacoronavirus and Porcine Epidemic Diarrhea Virus Increases Disease Severity, Cell Trophism and Earlier Upregulation of IFN-α and IL12. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Zhao, D.; Gao, X.; Zhou, P.; Zhang, L.; Zhang, Y.; Wang, Y.; Liu, X. Evaluation of the Immune Response in Conventionally Weaned Pigs Infected with Porcine Deltacoronavirus. Arch. Virol. 2020, 165, 1653–1658. [Google Scholar] [CrossRef]
- Moon, H.W.; Norman, J.O.; Lambert, G. Age Dependent Resistance to Transmissible Gastroenteritis of Swine (TGE) I. Clinical Signs and Some Mucosal Dimensions in Small Intestine. Can. J. Comp. Med. 1973, 37, 157. [Google Scholar]
- Jung, K.; Saif, L.J. Porcine Epidemic Diarrhea Virus Infection: Etiology, Epidemiology, Pathogenesis and Immunoprophylaxis. Vet. J. 2015, 204, 134–143. [Google Scholar] [CrossRef]
- Zimmerman, J.; Karriker, L.; Ramirez, A.; Schwartz, K.; Stevenson, G.; Zhang, J. (Eds.) Diseases of Swine, 11th ed.; Wiley: Hoboken, NJ, USA, 2019; pp. 494–508. [Google Scholar]
- Bjustrom-Kraft, J.; Woodard, K.; Giménez-Lirola, L.; Rotolo, M.; Wang, C.; Sun, Y.; Lasley, P.; Zhang, J.; Baum, D.; Gauger, P.; et al. Porcine Epidemic Diarrhea Virus (PEDV) Detection and Antibody Response in Commercial Growing Pigs. Bmc Vet. Res. 2016, 12, 1–8. [Google Scholar] [CrossRef]
- Mora-Díaz, J.C.; Temeeyasen, G.; Magtoto, R.; Rauh, R.; Nelson, W.; Carrillo-Ávila, J.A.; Zimmerman, J.; Piñeyro, P.; Giménez-Lirola, L. Infection and Immune Response to Porcine Hemagglutinating Encephalomyelitis Virus in Grower Pigs. Vet. Microbiol. 2021, 253, 108958. [Google Scholar] [CrossRef]
- Annamalai, T.; Saif, L.J.; Lu, Z.; Jung, K. Age-Dependent Variation in Innate Immune Responses to Porcine Epidemic Diarrhea Virus Infection in Suckling versus Weaned Pigs. Vet. Immunol. Immunopathol. 2015, 168, 193. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPI a | Pen 1 | Pen 2 | Pen 3 | Pen 4 | Pen 5 | Pen 6 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oral Fluid/Feces | Oral Fluid/Feces | Oral Fluid/Feces | Oral Fluid/Feces | Oral Fluid/Feces | Oral Fluid/Feces | |||||||||||||
| RT-qPCR b | CPE c | IFA d | RT-qPCR | CPE | IFA | RT-qPCR | CPE | IFA | RT-qPCR | CPE | IFA | RT-qPCR | CPE | IFA | RT-qPCR | CPE | IFA | |
| 0 | - e/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 2 | -/- | -/- | -/- | 38.5/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 4 | -/- | -/- | -/- | 37.1/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 6 | 35.1/37.4 | -/- | -/- | 28.2/25.0 | -/- | -/- | -/- | -/- | -/- | -/38.4 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 8 | 29.6/- | -/- | -/- | 29.7/37.5 | -/- | -/- | 29.8/38.7 | -/- | -/- | 30.0/31.3 | -/- | -/- | 29.5/30.5 | -/- | -/- | 30.1/- | -/- | -/- |
| 10 | 29.7/31.4 | -/- | -/- | 23.7/20.0 | -/- | -/- | 31.1/32.1 | -/- | -/- | 23.6/22.7 | -/- | -/- | 30.8/35.8 | -/- | -/- | 30.0/34.4 | -/- | -/- |
| 12 | 36.4/36.9 | -/- | -/- | 32.8/37.0 | -/- | -/- | 36.5/- | -/- | -/- | 30.9/27.7 | -/- | -/- | 38.5/- | -/- | -/- | -/- | -/- | -/- |
| 14 | 38.6/- | -/- | -/- | 39.0/- | -/- | -/- | 38.9/- | -/- | -/- | 38.6/34.3 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 16 | 38.2/- | -/- | -/- | 36.7/38.2 | -/- | -/- | 38.3/- | -/- | -/- | -/- | -/- | -/- | -/35.9 | -/- | -/- | -/- | -/- | -/- |
| 18 | -/- | -/- | -/- | 34.9/- | -/- | -/- | -/- | -/- | -/- | 38.1/35.7 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 20 | -/- | -/- | -/- | -/- | -/- | -/- | 38.9/- | -/- | -/- | 31.2/30.0 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 22 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/34.5 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 24 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 26 | -/- | -/- | -/- | 39.1/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 28 | -/- | -/- | -/- | 37.2/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 30 | -/- | -/- | -/- | 39.3/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 32 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 34 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 36 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 38 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 40 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
| 42 | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- | -/- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yen, L.; Mora-Díaz, J.C.; Rauh, R.; Nelson, W.; Castillo, G.; Ye, F.; Zhang, J.; Baum, D.; Zimmerman, J.; Nelli, R.; et al. Characterization of the Subclinical Infection of Porcine Deltacoronavirus in Grower Pigs under Experimental Conditions. Viruses 2022, 14, 2144. https://doi.org/10.3390/v14102144
Yen L, Mora-Díaz JC, Rauh R, Nelson W, Castillo G, Ye F, Zhang J, Baum D, Zimmerman J, Nelli R, et al. Characterization of the Subclinical Infection of Porcine Deltacoronavirus in Grower Pigs under Experimental Conditions. Viruses. 2022; 14(10):2144. https://doi.org/10.3390/v14102144
Chicago/Turabian StyleYen, Lu, Juan Carlos Mora-Díaz, Rolf Rauh, William Nelson, Gino Castillo, Fangshu Ye, Jianqiang Zhang, David Baum, Jeffrey Zimmerman, Rahul Nelli, and et al. 2022. "Characterization of the Subclinical Infection of Porcine Deltacoronavirus in Grower Pigs under Experimental Conditions" Viruses 14, no. 10: 2144. https://doi.org/10.3390/v14102144
APA StyleYen, L., Mora-Díaz, J. C., Rauh, R., Nelson, W., Castillo, G., Ye, F., Zhang, J., Baum, D., Zimmerman, J., Nelli, R., & Giménez-Lirola, L. (2022). Characterization of the Subclinical Infection of Porcine Deltacoronavirus in Grower Pigs under Experimental Conditions. Viruses, 14(10), 2144. https://doi.org/10.3390/v14102144