
Serological Positivity against Selected Flaviviruses and Alphaviruses in Free-Ranging Bats and Birds from Costa Rica Evidence Exposure to Arboviruses Seldom Reported Locally in Humans

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Mayer, S.V.; Tesh, R.B.; Vasilakis, N. The emergence of arthropod-borne viral diseases: A global prospective on Dengue, Chikungunya and Zika Fevers. Acta Trop. 2016, 166, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.I.; Clark, D.C.; Pesavento, P.; Lerche, N.W.; Luciw, P.A.; Reisen, W.K.; Brault, A.C. Comparative pathogenesis of epidemic and enzootic Chikungunya viruses in a pregnant Rhesus Macaque model. Am. J. Trop. Med. Hyg. 2010, 83, 1249–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilakis, N.; Weaver, S.C. Flavivirus transmission focusing on Zika. Curr. Opin. Virol. 2017, 22, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Unni, S.K. Chikungunya virus: Host pathogen interaction. Rev. Med. Virol. 2011, 21, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Chen, R.; Weaver, S.C. Interspecies transmission and chikungunya virus emergence. Curr. Opin. Virol. 2016, 16, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krutikov, M.; Manson, J. Chikungunya virus infection: An update on joint manifestations and management. Rambam. Maimonides. Med. J. 2016, 7, e0033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuno, G.; Mackenzie, J.S.; Junglen, S.; Hubálek, Z.; Plyusnin, A.; Gubler, D.J. Vertebrate reservoirs of arboviruses: Myth, synonym of amplifier, or reality? Viruses 2017, 9, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, P.S.; Doyle, M.M.; Smart, K.M.; Young, C.C.W.; Drape, G.W.; Johnson, C.K. Predicting wildlife reservoirs and global vulnerability to zoonotic Flaviviruses. Nat. Commun. 2018, 9, 5425. [Google Scholar] [CrossRef]
- Guzmán-Terán, C.; Calderón-Rangel, A.; Rodriguez-Morales, A.; Mattar, S. Venezuelan equine encephalitis virus: The problem is not over for tropical America. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 19. [Google Scholar] [CrossRef]
- Ministerio de Salud. Análisis de la Situación de Salud: Dengue 2017. Available online: https://www.ministeriodesalud.go.cr/index.php/vigilancia-de-la-salud/analisis-de-situacion-de-salud/3280-dengue-2017/file (accessed on 15 September 2017).
- Ministerio de Salud. Análisis de la Situación de Salud: Zika 2017. Available online: https://www.ministeriodesalud.go.cr/index.php/vigilancia-de-la-salud/analisis-de-situacion-de-salud/3278-zika-2017/file (accessed on 15 September 2017).
- Ministerio de Salud. Análisis de la Situación de Salud: Chikungunya 2017. Available online: https://www.ministeriodesalud.go.cr/index.php/vigilancia-de-la-salud/analisis-de-situacion-de-salud/3279-chikungunya-2017/file (accessed on 15 September 2017).
- Medlin, S.; Deardorff, E.R.; Hanley, C.S.; Vergneau-Grosset, C.; Siudak-Campfield, A.; Dallwig, R.; Travassos da Rosa, A.; Tesh, R.B.; Martin, M.P.; Weaver, S.C.; et al. Serosurvey of selected aboviral pathogens in free-ranging, two-toed sloths (Choloepus hoffmani) and three-toed sloths (Bradypus variegatus) in Costa Rica, 2005–2007. J. Wildl. Dis. 2016, 52, 883–892. [Google Scholar] [CrossRef] [Green Version]
- Berger, S. Yellow Fever: Global Status, 2019th ed.; Gideon E-Book Series; Gideon Informatics Inc.: Los Angeles, CA, USA, 2019; pp. 70–71. [Google Scholar]
- Jiménez, C.; Romero, M.; Piche, M.; Baldi, M.; Alfaro, A.; Chaves, A.; Morales, J.; León, B.; Hutter, S.; Corrales-Aguilar, E. Arboviral encephalitis in Costa Rican horses: 2009–2016. Int. J. Infect. Dis. 2016, 53, 153. [Google Scholar] [CrossRef] [Green Version]
- León, B.; Jiménez, C.; González, R.; Ramirez-Carvajal, L. First complete coding sequence of a venezuelan equine encephalitis virus strain isolated from an equine encephalitis case in Costa Rica. Microbiol. Resour. Announc. 2019, 5, e00672-19. [Google Scholar] [CrossRef] [Green Version]
- Carrara, S.; Gonzales, M.; Ferro, C.; Tamayo, M.; Aronson, J.; Paessler, S.; Anishchenko, M.; Boshell, J.; Weaver, S.C. Venezuelan Equine Encephalitis Virus Infection of Spiny Rats. Emerg. Infect. Dis. 2005, 11, 663–669. [Google Scholar] [CrossRef]
- Soto-Garita, C.; Somogyi, T.; Vicente-Santos, A.; Corrales-Aguilar, E. Molecular characterization of two major dengue outbreaks in Costa Rica. Am. J. Trop. Med. Hyg. 2016, 95, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Franck, P.T.; Johnson, K.M. An outbreak of Venezuelan equine encephalomeylitis in Central America. Evidence for exogenous source of a virulent virus subtype. Am. J. Epidemiol. 1971, 94, 487–495. [Google Scholar] [CrossRef]
- León, B.; Käsbohrer, A.; Hutter, S.E.; Baldi, M.; Firth, C.L.; Romero-Zúñiga, J.J.; Jiménez, C. National seroprevalence and risk factors for eastern equine encephalitis and Venezuelan equine encephalitis in Costa Rica. J. Equine Vet. Sci. 2020, 92, 103140. [Google Scholar] [CrossRef]
- Rodríguez-Herrera, B.; Ramírez-Fernández, J.D.; Villalobos-Chaves, D.; Sánchez, R. Actualización de la Lista de Especies de Mamíferos Vivientes de Costa Rica. Mastozool. Neotrop. 2014, 21, 275–289. [Google Scholar]
- Garrigues, R.; Camacho-Varela, P.; Montoya, M.; O’Donnell, P.; Ramírez-Alán, O.; Zook, J. Lista Oficial de las Aves de Costa Rica–Actualización 2018. Comité de Especies Raras y Registros Ornitológicos de Costa Rica (Comité Científico), Asociación Ornitológica de Costa Rica. Zeledonia 2018, 22, 52–58. [Google Scholar]
- Contigiani, M.A.; Diaz, L.A.; Spinsanti, L.I. General aspects on arboviruses. In Arthropod Borne Diseases, 1st ed.; Brisola-Marcondes, C., Ed.; Springer: Cham, Switzerland, 2017; pp. 61–71. [Google Scholar]
- Moreira-Soto, A.; Corrales-Aguilar, E. Bats and flaviviruses. In Bats and Viruses: Current Research and Future Trends, 1st ed.; Corrales-Aguilar, E., Schwemmle, M., Eds.; Caister Academic Press: Poole, UK, 2020; pp. 7–26. [Google Scholar]
- Borchert, R.; Meyer, S.A.; Felger, R.S.; Porter-Bolland, L. Environmental control of flowering periodicity in Costa Rican and Mexican tropical dry forests. Glob. Ecol. Biogeogr. 2004, 13, 409–425. [Google Scholar] [CrossRef]
- Instituto Meteorológico Nacional. Clima en Costa Rica: Pacífico Norte. Available online: https://www.imn.ac.cr/clima-en-costa-rica (accessed on 15 September 2017).
- Garrigues, R.; Dean, R. The Birds of Costa Rica: A Field Guide, 1st ed.; Cornell University Press: New York, NY, USA, 2007; pp. 1–440. [Google Scholar]
- York, H.A.; Rodríguez-Herrera, B.; Laval, R.K.; Timm, R.M.; Lindsay, K.E. Field key to the bats of Costa Rica and Nicaragua. J. Mammal. 2019, 100, 1726–1749. [Google Scholar] [CrossRef]
- American Veterinary Medical Association. Guidelines for the Euthanasia of Animals: 2013 Edition. Available online: https://www.avma.org/sites/default/files/resources/euthanasia.pdf (accessed on 12 November 2020).
- Scaramozzino, N.; Crance, J.M.; Jouan, A.; Debriel, D.A.; Stoll, F.; Garin, D. Comparison of flavivirus universal primer pairs and development of a rapid, highly sensitive heminested reverse transcription-PCR assay for detection of flaviviruses targeted to a conserved region of the NS5 gene sequences. J. Clin. Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef] [Green Version]
- Grywna, K.; Kupfer, B.; Panning, M.; Drexler, J.F.; Emmerich, P.; Drosten, C.; Kümmerer, B.M. Detection of All Species of the Genus Alphavirus by Reverse Transcription-PCR with Diagnostic Sensitivity. J. Clin. Microbiol. 2010, 48, 3386–3387. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Santos, A.; Moreira-Soto, A.; Soto-Garita, C.; Chaverri, L.G.; Chaves, A.; Drexler, J.F.; Morales, J.A.; Alfaro-Alarcón, A.; Rodríguez-Herrera, B.; Corrales-Aguilar, E. Neotropical bats that co-habit with humans’ function as dead-end hosts for dengue virus. PLoS Negl. Trop. Dis. 2017, 11, e0005537. [Google Scholar] [CrossRef] [PubMed]
- Goupil, B.A.; McNulty, M.A.; Martin, M.J.; McCracken, M.K.; Christofferson, R.C.; Mores, N.C. Novel lesions of bones and joints associated with chikungunya virus infection in two mouse models of disease: New insights into disease pathogenesis. PLoS ONE 2016, 11, e0155243. [Google Scholar] [CrossRef]
- Yu, J.; Liu, X.; Ke, C.; Wu, Q.; Lu, W.; Qin, Z.; He, X.; Liu, Y.; Deng, J.; Xu, S.; et al. Effective suckling C57BL/6, kunming, and BALB/c mouse models with remarkable neurological manifestation for zika virus infection. Viruses 2017, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Sarathy, V.V.; Milligan, G.N.; Bourne, N.; Barrett, A.D. Mouse models of dengue virus infection for vaccine testing. Vaccine 2015, 33, 7051–7060. [Google Scholar] [CrossRef] [Green Version]
- Hubálek, Z.; Rudolf, I.; Nowotny, N. Arboviruses pathogenic for domestic and wild animals. Adv. Virus Res. 2014, 89, 201–275. [Google Scholar] [PubMed]
- Senne, D.A.; Pedersen, J.C.; Hutto, D.L.; Taylor, W.D.; Schmitt, B.J.; Panigrahy, B. Pathogenicity of West Nile virus in chickens. Avian. Dis. 2000, 44, 642–649. [Google Scholar] [CrossRef]
- Farfán-Ale, J.A.; Blitvich, B.J.; Marlenee, N.L.; Loroño-Pino, M.A.; Puerto-Manzano, F.; García-Rejón, J.E.; Rosado-Paredes, E.P.; Flores-Flores, L.F.; Ortega-Salazar, A.; Chávez-Medina, J.; et al. Antibodies to West Nile virus in asymptomatic mammals, birds, and reptiles in the Yucatan Peninsula of Mexico. Am. J. Trop. Med. Hyg. 2006, 74, 908–914. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.; Coffey, L.L.; Burkett-Cadena, N.; Day, J.F. Reemergence of St. Louis Encephalitis Virus in the Americas. Emerg. Infect. Dis. 2018, 24, 2150–2157. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Tommy Lam, T.T.Y.; Heberlein Larson, L.A.; Smole, S.C.; Auguste, A.J.; Hennigan, S.; Halpin, R.A.; Fedorova, N.; Puri, V.; Stockwell, T.B.; et al. Large-scale complete-genome sequencing and phylodynamic analysis of eastern equine encephalitis virus reveals source-sink transmission dynamics in the United States. J. Virol. 2018, 92, e00074-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelen, V.P.; Downs, J.A.; Unnasch, T.; Stark, L. A risk index model for predicting eastern equine encephalitis virus transmission to horses in Florida. Appl. Geogr. 2014, 1, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, A.S.; Austin, K.; Gardner, C.L.; Zuiani, A.; Reed, D.S.; Trobaugh, D.W.; Sun, C.; Basore, K.; Williamson, L.E.; Crowe, J.E., Jr.; et al. Protective antibodies against Eastern equine encephalitis virus bind to epitopes in domains A and B of the E2 glycoprotein. Nat. Microbiol. 2019, 4, 187–197. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Laboratory Diagnosis for the Infection for Yellow Fever Virus. Available online: https://www.paho.org/hq/index.php?option=com_docman&task=doc_view&gid=43571&Itemid=270&lang=en (accessed on 15 April 2018).
- Thompson, N.N.; Auguste, A.J.; Travassos da Rosa, A.P.; Carrington, C.V.; Blitvich, B.J.; Chadee, D.D.; Tesh, R.B.; Weaver, S.C.; Adesiyun, A.A. Seroepidemiology of selected alphaviruses and flaviviruses in bats in Trinidad. Zoonoses Public Health 2014, 62, 53–60. [Google Scholar] [CrossRef]
- Bittar, C.; Machado, R.R.G.; Comelis, M.T.; Bueno, L.M.; Morielle-Versute, E.; Beguelini, M.R.; de Souza, R.P.; Nogueira, M.L.; Rahal, P. Lack of serological and molecular evidence of arbovirus infections in bats from Brazil. PLoS ONE 2018, 13, e0207010. [Google Scholar] [CrossRef]
- Guzmán, C.; Calderón, A.; Oviedo, T.; Mattar, S.; Catañeda, J.; Rodríguez, V.; Moraes-Figueiredo, L.T. Mollecular an celular evidence if natural Venezuelan equine encephalitis virus infection in frugivorous bats in Colombia. Vet. World 2020, 13, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.; Pontier, D.; Filippi-Codaccioni, O.; Pons, J.; Postigo-Hidalgo, I.; Duhayer, J.; Brünink, S.; Drexler, J.F. Venezuelan equine encephalitis complex alphavirus in bats, French Guiana. Emerg. Infect. Dis. 2021, 27, 1141–1145. [Google Scholar] [CrossRef]
- Herbold, J.; Heuschele, W.; Berry, R.; Parsons, M. Reservoir of St. Louis encephalitis virus in Ohio bats. Am. J. Vet. Res. 1983, 44, 1889–1893. [Google Scholar]
- Stone, D.; Lyons, A.; Huang, Y.J.; Vanlandingham, D.; Higgs, S.; Blitvich, B.; Adesiyun, A.; Santana, S.; Leiser-Miller, L.; Cheetham, S. Serological evidence of widespread exposure of Grenada fruit bats to chikungunya virus. Zoonoses Public Health 2018, 65, 505–511. [Google Scholar] [CrossRef]
- Pilipski, J.D.; Pilipski, L.M.; Risley, L.S. West Nile virus antibodies in bats from New Jersey and New York. J. Wildl. Dis. 2004, 40, 335–337. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.; Bunning, M.; Gordy, P.; Panella, N.; Blitvich, B.; Bowen, R. Experimental and natural infection of North American bats with West Nile virus. Am. J. Trop. Med. Hyg. 2005, 73, 467–469. [Google Scholar] [CrossRef]
- Fagre, A.C.; Kading, R.C. Can bats serve as reservoirs for Arboviruses? Viruses 2019, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Thompson, N.N.; Auguste, A.J.; Coombs, D.; Blitvich, B.J.; Carrington, C.V.; da Rosa, A.P.; Wang, E.; Chadee, D.D.; Drebot, M.A.; Tesh, R.B.; et al. Serological evidence of flaviviruses and alphaviruses in livestock and wildlife in Trinidad. Vector Borne Zoonotic Dis. 2012, 12, 969–978. [Google Scholar] [CrossRef] [Green Version]
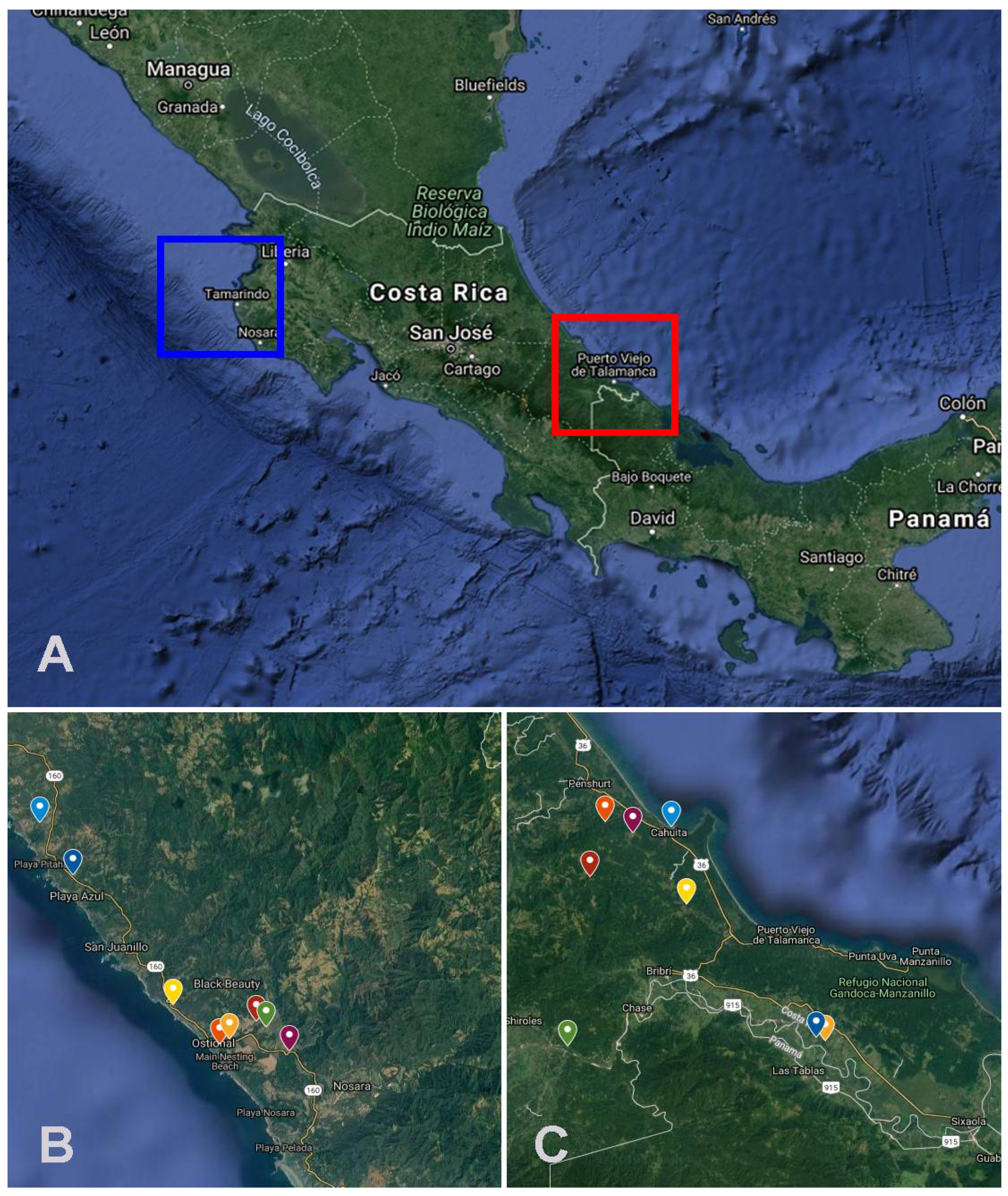

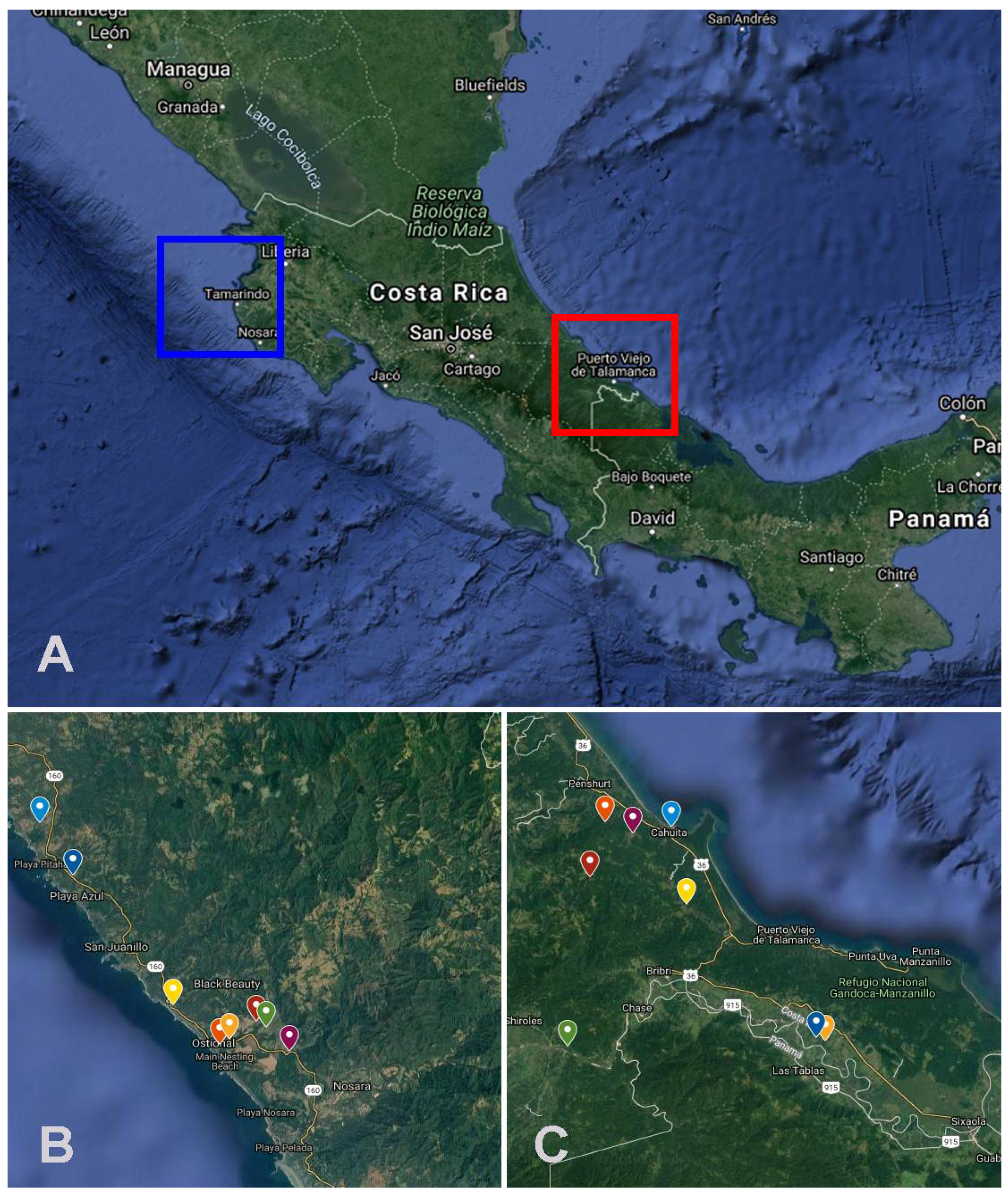{kind=link}
| Species | Sex | Positive Results in PRNT 1:10 (Putative Cross-Reactions) |
|---|---|---|
| Glossophaga soricina | Female | DENV-1/DENV-3 |
| Artibeus jamaicensis | Female | DENV-1 DENV-2 |
| Artibeus phaeotis | Female | DENV-1 DENV-2 DENV-4 |
| Artibeus phaeotis | Female | DENV-1 DENV-2 DENV-4 |
| Phyllostomus discolor | Male | DENV-1 EEEV |
| Phyllostomus discolor | Female | DENV-1 DENV-2 DENV-3 EEEV |
| Noctilio albiventris | Male | DENV-1 EEEV |
| Artibeus lituratus | Female | DENV-1/DENV-2/ DENV-3 |
| Artibeus jamaicensis | Male | DENV-1 DENV-2 EEEV |
| Artibeus phaeotis | Male | DENV-1 DENV-2 WNV SLEV |
| Carollia perspicillata | Male | DENV-1 DENV-2 |
| Artibeus watsoni | Male | DENV-1 DENV-2 DENV-3 DENV-4 |
| Artibeus jamaicensis | Female | DENV-1 SLEV |
| Carollia perspicillata | Male | DENV-1 DENV-2 |
| Glossophaga soricina | Female | DENV-1 DENV-2 DENV-3 DENV-4 |
| Artibeus jamaicensis | Female | DENV-1 DENV-2 YFV WNV |
| Artibeus jamaicensis | Female | SLEV, EEEV |
| Uroderma convexum | Female | DENV-1 DENV-2 DENV-3 SLEV |
| Uroderma convexum | Female | DENV-1 DENV-2 SLEV |
| Carollia perspicillata | Female | DENV-1 DENV-2 |
| Artibeus jamaicensis | Male | DENV-1 DENV-2 |
| Artibeus jamaicensis | Male | DENV-1 DENV-4 SLEV |
| Species | Sex | Exclusive Reaction in PRNT 1:10 |
|---|---|---|
| Uroderma convexum | Male | DENV-1 |
| Artibeus phaeotis | Male | DENV-1 |
| Desmodus rotundus | Female | DENV-1 |
| Sturnira parvidens | Female | DENV-1 |
| Artibeus jamaicensis | Male | DENV-1 |
| Chiroderma salvini | Male | DENV-1 |
| Ptenorotus mesoamericanus | Female | DENV-1 |
| Carollia perspicillata | Female | DENV-1 |
| Uroderma convexum | Male | DENV-1 |
| Glossophaga soricina | Male | DENV-1 |
| Loncophylla robusta | Female | DENV-1 |
| Myotis nigricans | Female | DENV-1 |
| Phyllostomus discolor | Male | DENV-1 |
| Artibeus jamaicensis | Male | DENV-1 |
| Glossophaga soricina | Male | DENV-1 |
| Carollia perspicillata | Female | DENV-1 |
| Carollia perspicillata | Male | DENV-1 |
| Artibeus jamaicensis | Female | DENV-1 |
| Uroderma convexum | Male | DENV-1 |
| Carollia castanea | Female | DENV-1 |
| Rhogeessa io | Female | DENV-1 |
| Uroderma convexum | Male | DENV-1 |
| Glossophaga soricina | Female | DENV-2 |
| Artibeus lituratus | Male | DENV-2 |
| Artibeus watsoni | Male | DENV-2 |
| Glossophaga soricina | Male | DENV-2 |
| Artibeus phaeotis | Female | DENV-3 |
| Desmodus rotundus | Female | DENV-4 |
| Carollia castanea | Female | WNV |
| Carollia perspicillata | Female | EEEV |
| Phyllostomus discolor | Male | EEEV |
| Phyllostomus discolor | Male | EEEV |
| Artibeus jamaicensis | Male | EEEV |
| Carollia perspicillata | Male | VEEV |
| Species | Sex | Cross-Reactions in PRNT 1:20 |
|---|---|---|
| Molossus sinaloe | Female | SLEV, DENV-3 ** |
| Molossus sinaloe | Female | WNV, DENV-3 ** |
| Molossus sinaloe | Female | SLEV, DENV-3 ** |
| Molossus sinaloe | Female | WNV, DENV-3 ** |
| Molossus sinaloe | Female | WNV *, DENV-3 ** |
| Molossus sinaloe | Male | WNV, SLEV, DENV-2 **, DENV-3 ** |
| Molossus sinaloe | Male | SLEV, DENV-3 ** |
| Molossus sinaloe | Male | SLEV, DENV-1 **, DENV-2 **, DENV4 ** |
| Uroderma bilobatum | Male | WNV, DENV-3 ** |
| Species | Sex | PRNT 1:10 Result |
|---|---|---|
| Aimophila ruficauda | Male | EEEV |
| Turdus grayi | Male | SLEV |
| Pitangus sulphuratus | Female | EEEV |
| Quiscalus mexicanus | Female | VEEV |
| Icterus pustulatus | Male | EEEV/VEEV |
| Icterus pustulatus | Male | EEEV |
| Campylorhynchus rufinucha | Female | WNV |
| Pitangus sulphuratus | Male | VEEV |
| Crotophaga sulcirostris | Male | EEEV |
| Myozetetes similis | Male | SLEV |
| Empidonax virescens * | Male | SLEV/EEEV/VEEV |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrantes Murillo, D.F.; Piche-Ovares, M.; Gamboa-Solano, J.C.; Romero, L.M.; Soto-Garita, C.; Alfaro-Alarcón, A.; Corrales-Aguilar, E. Serological Positivity against Selected Flaviviruses and Alphaviruses in Free-Ranging Bats and Birds from Costa Rica Evidence Exposure to Arboviruses Seldom Reported Locally in Humans. Viruses 2022, 14, 93. https://doi.org/10.3390/v14010093
Barrantes Murillo DF, Piche-Ovares M, Gamboa-Solano JC, Romero LM, Soto-Garita C, Alfaro-Alarcón A, Corrales-Aguilar E. Serological Positivity against Selected Flaviviruses and Alphaviruses in Free-Ranging Bats and Birds from Costa Rica Evidence Exposure to Arboviruses Seldom Reported Locally in Humans. Viruses. 2022; 14(1):93. https://doi.org/10.3390/v14010093
Chicago/Turabian StyleBarrantes Murillo, Daniel Felipe, Marta Piche-Ovares, José Carlos Gamboa-Solano, Luis Mario Romero, Claudio Soto-Garita, Alejandro Alfaro-Alarcón, and Eugenia Corrales-Aguilar. 2022. "Serological Positivity against Selected Flaviviruses and Alphaviruses in Free-Ranging Bats and Birds from Costa Rica Evidence Exposure to Arboviruses Seldom Reported Locally in Humans" Viruses 14, no. 1: 93. https://doi.org/10.3390/v14010093
APA StyleBarrantes Murillo, D. F., Piche-Ovares, M., Gamboa-Solano, J. C., Romero, L. M., Soto-Garita, C., Alfaro-Alarcón, A., & Corrales-Aguilar, E. (2022). Serological Positivity against Selected Flaviviruses and Alphaviruses in Free-Ranging Bats and Birds from Costa Rica Evidence Exposure to Arboviruses Seldom Reported Locally in Humans. Viruses, 14(1), 93. https://doi.org/10.3390/v14010093