
FMDV Leader Protein Interacts with the NACHT and LRR Domains of NLRP3 to Promote IL-1β Production

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Clinical Specimens, and Blood Samples
2.2. Cells and Cultures
2.3. Viruses and Infection
2.4. Reagents and Antibodies
2.5. Plasmids Construction
2.6. Lentivirus Production and Infection
2.7. Coimmunoprecipitation Assay (Co-IP)
2.8. Western Blot Analysis
2.9. Confocal Microscopy
2.10. RNA Extraction and RT-PCR
2.11. Luciferase Reporter Assays
2.12. ASC Oligomerization
2.13. Enzyme-Linked Immunosorbent Assay (ELISA)
2.14. Mature IL-1β Measurement:
2.15. Reagent Treatment
2.16. Statistical Analysis
3. Results
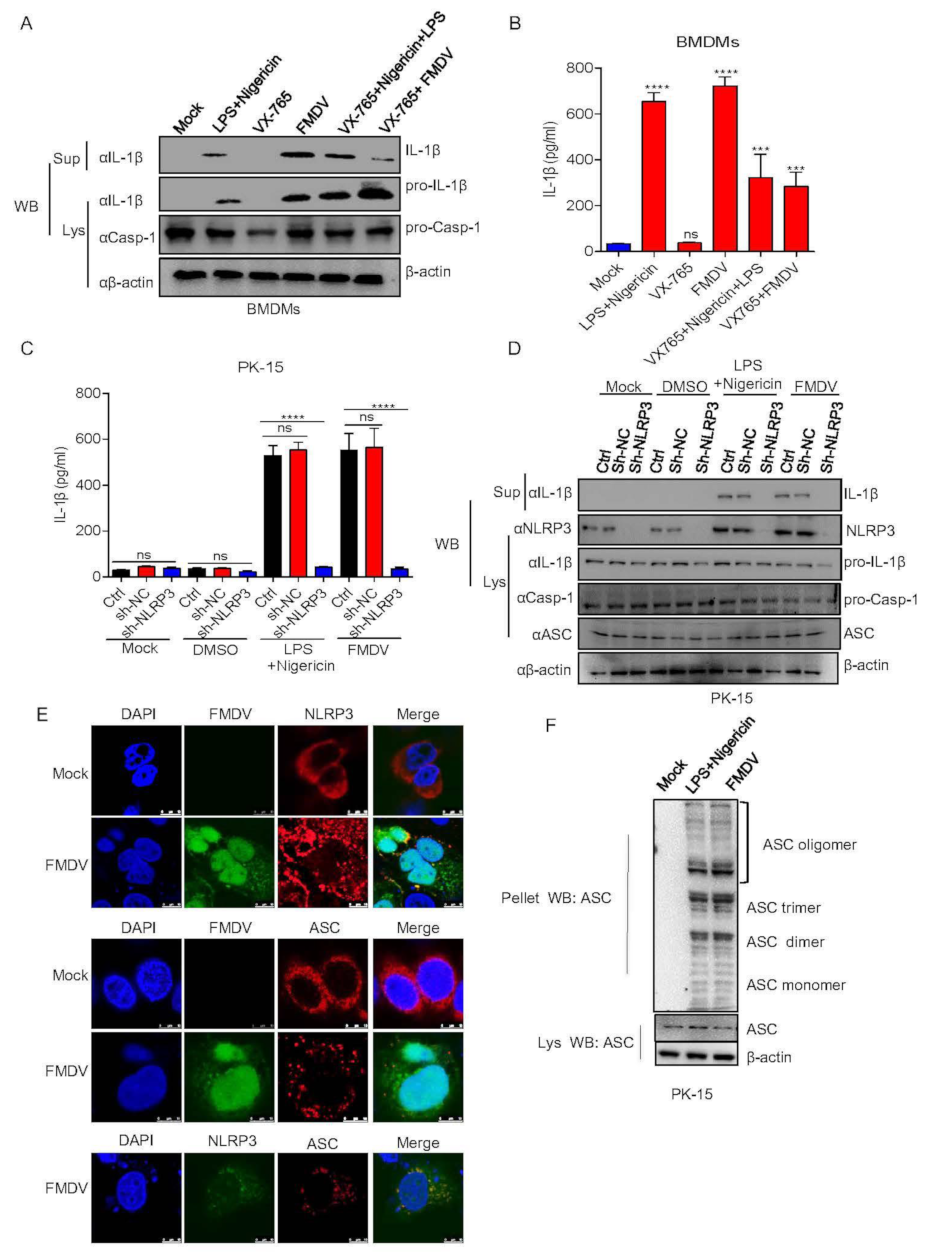
3.1. FMDV Infection Induces IL-1β Secretion in Pigs
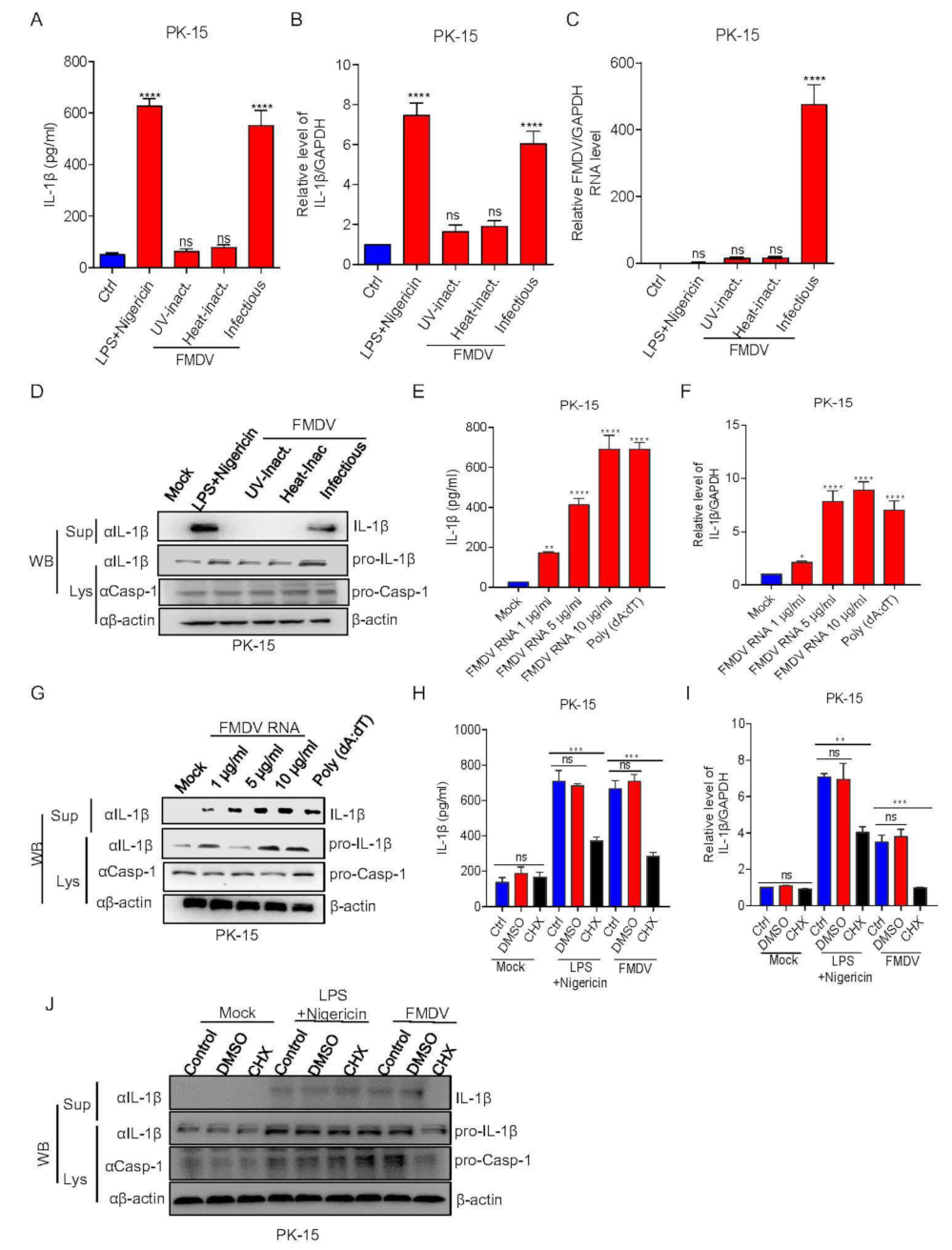
3.2. FMDV Activates the NLRP3 Inflammasome to Induce IL-1β Secretion in Pig Cells
3.3. FMDV Proteins Are Involved in NLRP3 Inflammasome Activation
3.4. FMDV Leader Protein Promotes NLRP3-Mediated IL-1β Production
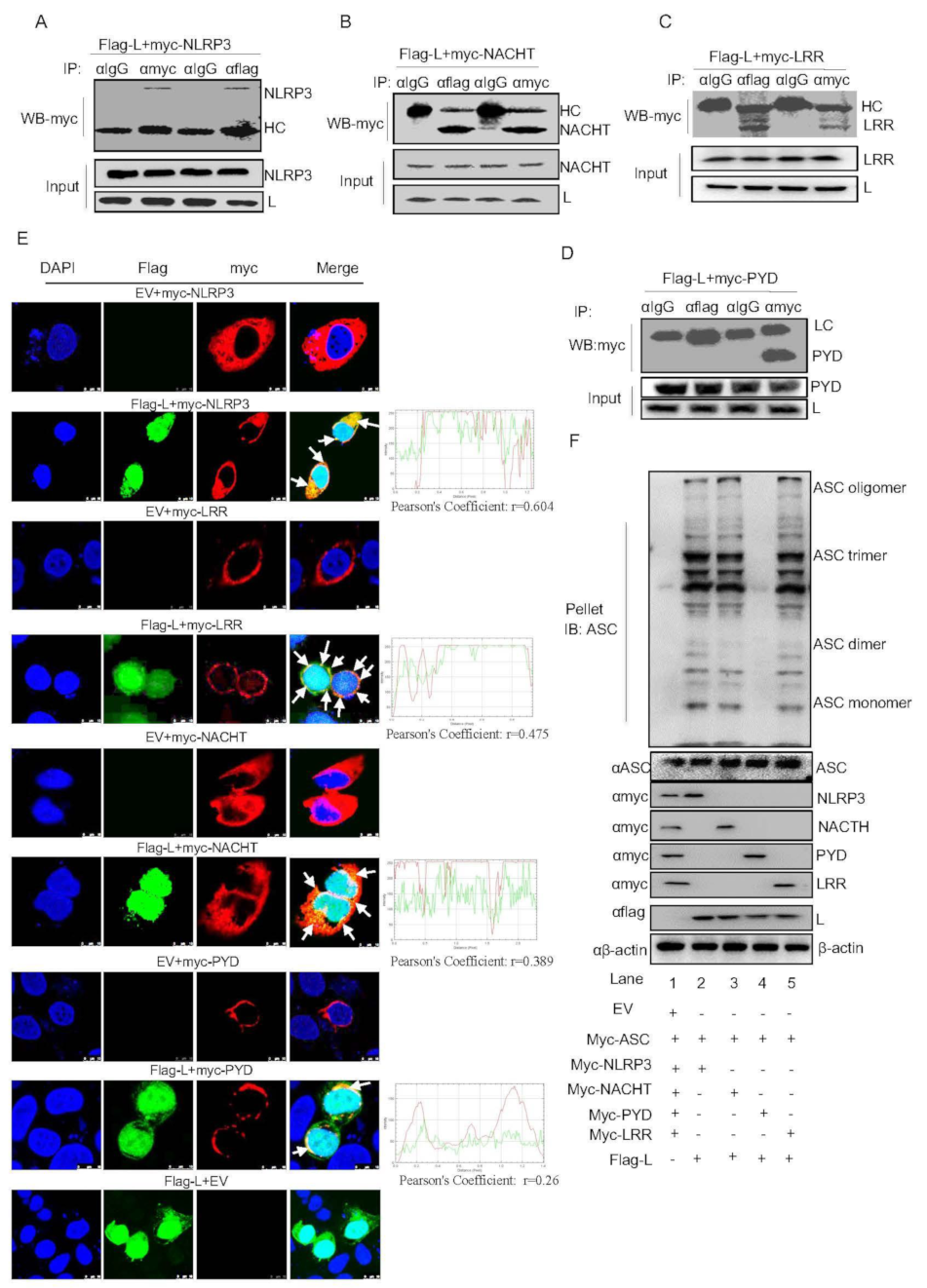
3.5. FMDV Lpro Interacts with the NACHT and LRR Domains of NLRP3 to Promote NLRP3 Inflammasome Assembly
3.6. Lpro Activated the NLRP3 Inflammasome through an Ion Channel
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.J.; Arzt, J.; Puckette, M.C.; Smoliga, G.R.; Pacheco, J.M.; Rodriguez, L.L. Mechanisms of foot-and-mouth disease virus tropism inferred from differential tissue gene expression. PLoS ONE 2013, 8, e64119. [Google Scholar] [CrossRef] [PubMed]
- Arzt, J.; Baxt, B.; Grubman, M.J.; Jackson, T.; Juleff, N.; Rhyan, J.; Rieder, E.; Waters, R.; Rodriguez, L.L. The pathogenesis of foot-and-mouth disease II: Viral pathways in swine, small ruminants, and wildlife; myotropism, chronic syndromes, and molecular virus-host interactions. Transbound Emerg. Dis. 2011, 58, 305–326. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Baranowski, E.; Escarmis, C.; Sobrino, F. Foot-and-mouth disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 297–308. [Google Scholar] [CrossRef]
- Carrillo, C.; Tulman, E.R.; Delhon, G.; Lu, Z.; Carreno, A.; Vagnozzi, A.; Kutish, G.F.; Rock, D.L. Comparative genomics of foot-and-mouth disease virus. J. Virol. 2005, 79, 6487–6504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Sun, S.Q.; Guo, H.C. Biological function of Foot-and-mouth disease virus non-structural proteins and non-coding elements. Virol. J. 2016, 13, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Li, D.; Yang, W.; Wang, Y.; Li, L.; Zheng, H. Foot-and-Mouth Disease Virus VP3 Protein Acts as a Critical Proinflammatory Factor by Promoting Toll-Like Receptor 4-Mediated Signaling. J. Virol. 2021, 95, e0112021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ahmed, R.; Paton, D.; Bashiruddin, J.B. Cytokine mRNA responses in bovine epithelia during foot-and-mouth disease virus infection. Vet. J. 2009, 179, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Zhi, X.; Zhang, Y.; Sun, S.; Zhang, Z.; Dong, H.; Luo, X.; Wei, Y.; Lu, Z.; Dou, Y.; Wu, R.; et al. NLRP3 inflammasome activation by Foot-and-mouth disease virus infection mainly induced by viral RNA and non-structural protein 2B. RNA Biol. 2020, 17, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, R.; Stichling, N.; Koelen, J.; Kuryk, L.; Lipiec, A.; Greber, U.F. Innate immunity to adenovirus. Hum. Gene Ther. 2014, 25, 265–284. [Google Scholar] [CrossRef] [Green Version]
- Aarreberg, L.D.; Wilkins, C.; Ramos, H.J.; Green, R.; Davis, M.A.; Chow, K.; Gale, M., Jr. Interleukin-1beta Signaling in Dendritic Cells Induces Antiviral Interferon Responses. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suthar, M.S.; Aguirre, S.; Fernandez-Sesma, A. Innate immune sensing of flaviviruses. PLoS Pathog. 2013, 9, e1003541. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Cao, R.; Hu, H.M. TLR and NLRP3 inflammasome-dependent innate immune responses to tumor-derived autophagosomes (DRibbles). Cell Death Dis. 2016, 7, e2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinarello, C.A. Immunological and inflammatory functions of the interleukin-1 family. Annu. Rev. Immunol. 2009, 27, 519–550. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, Z.M.; Wu, X.; Zhang, L.; Cao, Y.; Zhou, P. Distinct Molecular Mechanisms Underlying Potassium Efflux for NLRP3 Inflammasome Activation. Front. Immunol. 2020, 11, 609441. [Google Scholar] [CrossRef] [PubMed]
- Jager, E.; Murthy, S.; Schmidt, C.; Hahn, M.; Strobel, S.; Peters, A.; Staubert, C.; Sungur, P.; Venus, T.; Geisler, M.; et al. Calcium-sensing receptor-mediated NLRP3 inflammasome response to calciprotein particles drives inflammation in rheumatoid arthritis. Nat. Commun. 2020, 11, 4243. [Google Scholar] [CrossRef] [PubMed]
- Chevriaux, A.; Pilot, T.; Derangere, V.; Simonin, H.; Martine, P.; Chalmin, F.; Ghiringhelli, F.; Rebe, C. Cathepsin B Is Required for NLRP3 Inflammasome Activation in Macrophages, Through NLRP3 Interaction. Front. Cell Dev. Biol. 2020, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Petrilli, V.; Van Bruggen, R.; Steele, C.; Mossman, B.T.; Tschopp, J. Innate immune activation through Nalp3 inflammasome sensing of asbestos and silica. Science 2008, 320, 674–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Wang, Y.; Li, X.; Zhan, X.; Tang, M.; Fina, M.; Su, L.; Pratt, D.; Bu, C.H.; Hildebrand, S.; et al. NLRP3 activation and mitosis are mutually exclusive events coordinated by NEK7, a new inflammasome component. Nat. Immunol. 2016, 17, 250–258. [Google Scholar] [CrossRef]
- He, Y.; Zeng, M.Y.; Yang, D.; Motro, B.; Nunez, G. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature 2016, 530, 354–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Lu, L.; Li, L. NEK7: A novel promising therapy target for NLRP3-related inflammatory diseases. Acta Biochim. Biophys. Sin. (Shanghai) 2016, 48, 966–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, T.S.; de Sa, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Goncalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.C.; Scull, M.A.; Moore, C.B.; Holl, E.K.; McElvania-TeKippe, E.; Taxman, D.J.; Guthrie, E.H.; Pickles, R.J.; Ting, J.P. The NLRP3 inflammasome mediates in vivo innate immunity to influenza A virus through recognition of viral RNA. Immunity 2009, 30, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negash, A.A.; Olson, R.M.; Griffin, S.; Gale, M., Jr. Modulation of calcium signaling pathway by hepatitis C virus core protein stimulates NLRP3 inflammasome activation. PLoS Pathog. 2019, 15, e1007593. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xiao, F.; Wan, P.; Pan, P.; Zhang, Y.; Liu, F.; Wu, K.; Liu, Y.; Wu, J. EV71 3D Protein Binds with NLRP3 and Enhances the Assembly of Inflammasome Complex. PLoS Pathog. 2017, 13, e1006123. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, G.; De, W.; Luo, Z.; Pan, P.; Tian, M.; Wang, Y.; Xiao, F.; Li, A.; Wu, K.; et al. Zika virus infection induces host inflammatory responses by facilitating NLRP3 inflammasome assembly and interleukin-1beta secretion. Nat. Commun. 2018, 9, 106. [Google Scholar] [CrossRef]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol 2016, 5, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID50 for Quantitation of Virus Infectivity. Virol Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yanagi, Y.; Ichinohe, T. Encephalomyocarditis virus viroporin 2B activates NLRP3 inflammasome. PLoS Pathog. 2012, 8, e1002857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Xu, Y.; Li, H.; Tao, W.; Xiang, Y.; Huang, B.; Niu, J.; Zhong, J.; Meng, G. HCV genomic RNA activates the NLRP3 inflammasome in human myeloid cells. PLoS ONE 2014, 9, e84953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, H.D.; Smith, A.D.; Chen, C.; Urban, J.F., Jr. An in-depth comparison of the porcine, murine and human inflammasomes; lessons from the porcine genome and transcriptome. Vet. Microbiol. 2017, 202, 2–15. [Google Scholar] [CrossRef]
- Pelegrin, P.; Barroso-Gutierrez, C.; Surprenant, A. P2X7 receptor differentially couples to distinct release pathways for IL-1beta in mouse macrophage. J. Immunol. 2008, 180, 7147–7157. [Google Scholar] [CrossRef] [Green Version]
- Hafner-Bratkovic, I.; Susjan, P.; Lainscek, D.; Tapia-Abellan, A.; Cerovic, K.; Kadunc, L.; Angosto-Bazarra, D.; Pelegrin, P.; Jerala, R. NLRP3 lacking the leucine-rich repeat domain can be fully activated via the canonical inflammasome pathway. Nat. Commun. 2018, 9, 5182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boal-Carvalho, I.; Mazel-Sanchez, B.; Silva, F.; Garnier, L.; Yildiz, S.; Bonifacio, J.P.; Niu, C.; Williams, N.; Francois, P.; Schwerk, N.; et al. Influenza A viruses limit NLRP3-NEK7-complex formation and pyroptosis in human macrophages. EMBO Rep. 2020, 21, e50421. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.K.M.; Ma, X.; Abdullah, S.W.; Zheng, H. Activation and Inhibition of the NLRP3 Inflammasome by RNA Viruses. J. Inflamm. Res. 2021, 14, 1145–1163. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.J.; Mishra, R.; Schneider, K.S.; Medard, G.; Wettmarshausen, J.; Dittlein, D.C.; Shi, H.; Gorka, O.; Koenig, P.A.; Fromm, S.; et al. K(+) Efflux-Independent NLRP3 Inflammasome Activation by Small Molecules Targeting Mitochondria. Immunity 2016, 45, 761–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.S.; Subramanian, N.; Kim, A.I.; Aksentijevich, I.; Goldbach-Mansky, R.; Sacks, D.B.; Germain, R.N.; Kastner, D.L.; Chae, J.J. The calcium-sensing receptor regulates the NLRP3 inflammasome through Ca2+ and cAMP. Nature 2012, 492, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Minutoli, L.; Puzzolo, D.; Rinaldi, M.; Irrera, N.; Marini, H.; Arcoraci, V.; Bitto, A.; Crea, G.; Pisani, A.; Squadrito, F.; et al. ROS-Mediated NLRP3 Inflammasome Activation in Brain, Heart, Kidney, and Testis Ischemia/Reperfusion Injury. Oxid. Med. Cell. Longev. 2016, 2016, 2183026. [Google Scholar] [CrossRef] [PubMed]
- Rovira-Llopis, S.; Apostolova, N.; Banuls, C.; Muntane, J.; Rocha, M.; Victor, V.M. Mitochondria, the NLRP3 Inflammasome, and Sirtuins in Type 2 Diabetes: New Therapeutic Targets. Antioxid Redox Signal. 2018, 29, 749–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, N.; Li, C.C.; Di, B.; Xu, L.L. Recent advances in the NEK7-licensed NLRP3 inflammasome activation: Mechanisms, role in diseases and related inhibitors. J. Autoimmun. 2020, 113, 102515. [Google Scholar] [CrossRef]
- Sharif, H.; Wang, L.; Wang, W.L.; Magupalli, V.G.; Andreeva, L.; Qiao, Q.; Hauenstein, A.V.; Wu, Z.; Nunez, G.; Mao, Y.; et al. Structural mechanism for NEK7-licensed activation of NLRP3 inflammasome. Nature 2019, 570, 338–343. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sense Primers (5′-3′) | Anti-Sense Primers (5′-3′) |
|---|---|---|
| hGAPDH-qRT | GAGTCAACGGATTTGGTCGT | GACAAGCTTCCCGTTCTCAG |
| P-GAPDH-qRT | ACATGGCCTCCAAGGAGTAAGA | GATCGAGTTGGGGCTGTGACT |
| mGAPDH-qRT | CCATGTTCGTCATGGGTGTGAACCA | GCCAGTAGAGGCAGGGATGATGTTC |
| h-p65-qRT | TGAACCGAAACTCTGGCAGCTG | CATCAGCTTGCGAAAAGGAGCC |
| h-IL-1β-qRT | GCAAGGGCTTCAGGCAGGCCGCG | GGTCATTCTCCTGGAAGGTCTGTGGGC |
| FMDV-qRT | ACTGGGTTTTACAAACCTGTGA | GCGAGTCCTGCCACGGA |
| m-IL-1β-qRT | GCACTACAGGCTCCGAGATGAAC | TTGTCGTTGCTTGGTTCTCCTTGT |
| P-caspase 1-qRT | GAAATACTCCGAAGAAGTCCCAGA | GACCCCTTGCTTCTCACCAC |
| P-PYCARD-qRT | TCAAGGGTCACAGACGTGGA | TTTGGTGGGGTTGGTGTG |
| h-NLRP3-qRT | AAGGGCCATGGACTATTTCC | GACTCCACCCGATGACAGTT |
| h-ASC-qRT | AACCCAAGCAAGATGCGGAAG | 5′-TTAGGGCCTGGAGGAGCAAG-3′ |
| h-Casp-1-qRT | TCCAATAATGCAAGTCAAGCC | GCTGTACCCCAGATTTTGTAGCA |
| P-IL-1β-qRT | GACGGGCTTTTGTTCTGCTT | GGACATGGAGAAGCGATTTGT |
| P-p65-qRT | GGAACACGATGGCCACTTG | AAGAGGACATCGAGGTGTATTTCAC |
| P-Casp-1-qRT | GAAATACTCCGAAGAAGTCCCAGA | GACCCCTTGCTTCTCACCAC |
| P-NLRP3-qRT | CCTCTTTGGCCTTGTAAACC | TGGCTGGGCTCAATCTGTAG |
| P-ASC-qRT | GCTGGCTAGCATGGGGTGCACGCGTGAC | GCCGCTCGAGTCAGCTCTGCTCCAGGTCG |
| Genes | Sense Primers (5′-3′) | Anti-Sense Primers (5′-3′) |
|---|---|---|
| 2B | ATGCCCTTCTTCTTCTCCGA | TCACTTGTCATCATCGTCC |
| VP3 | ATGGGGATTTTCCCTGTGGCCTGTAG | TCACTTGTCATCATCGTCCTTATA |
| 3A | ATGATCTCAATTCCTTCCCAA | TTGGGAAGGAATTGAGATCAT |
| VP2 | ATGGATAAGAAAACCGAGGAG | CTCCTCGGTTTTCTTATCCAT |
| VP1 | ATGACCACTTCGACGGGCGAG | CTCGCCCGTCGAAGTGGTCAT |
| 3D | ATGGGGTTGATTGTGGACACC | GGTGTCCACAATCAACCCCAT |
| L | ATGAATACAACTGACTGTTTTATCGCT | TCATTTGAGCTTGCGTTGAACCTTGGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choudhury, S.M.; Ma, X.; Li, Y.; Nian, X.; Luo, Z.; Ma, Y.; Zhu, Z.; Yang, F.; Cao, W.; Zheng, H. FMDV Leader Protein Interacts with the NACHT and LRR Domains of NLRP3 to Promote IL-1β Production. Viruses 2022, 14, 22. https://doi.org/10.3390/v14010022
Choudhury SM, Ma X, Li Y, Nian X, Luo Z, Ma Y, Zhu Z, Yang F, Cao W, Zheng H. FMDV Leader Protein Interacts with the NACHT and LRR Domains of NLRP3 to Promote IL-1β Production. Viruses. 2022; 14(1):22. https://doi.org/10.3390/v14010022
Chicago/Turabian StyleChoudhury, Sk Mohiuddin, Xusheng Ma, Yuanyuan Li, Xiaofeng Nian, Zhikuan Luo, Yonghua Ma, Zixiang Zhu, Fan Yang, Weijun Cao, and Haixue Zheng. 2022. "FMDV Leader Protein Interacts with the NACHT and LRR Domains of NLRP3 to Promote IL-1β Production" Viruses 14, no. 1: 22. https://doi.org/10.3390/v14010022
APA StyleChoudhury, S. M., Ma, X., Li, Y., Nian, X., Luo, Z., Ma, Y., Zhu, Z., Yang, F., Cao, W., & Zheng, H. (2022). FMDV Leader Protein Interacts with the NACHT and LRR Domains of NLRP3 to Promote IL-1β Production. Viruses, 14(1), 22. https://doi.org/10.3390/v14010022