
Bacterial Toxins from Staphylococcus aureus and Bordetella bronchiseptica Predispose the Horse’s Respiratory Tract to Equine Herpesvirus Type 1 Infection

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Equine Respiratory Explant Model
2.2. Pretreatment with Bacterial Toxins (Hla and ACT)
2.3. Assessment of Mucosal Morphology and Viability
2.4. EHV-1 Inoculation
2.5. Evaluation of EHV-1 Infection by Immunofluorescent Stainings and Confocal Microscopy
2.6. Statistical Analysis
3. Results
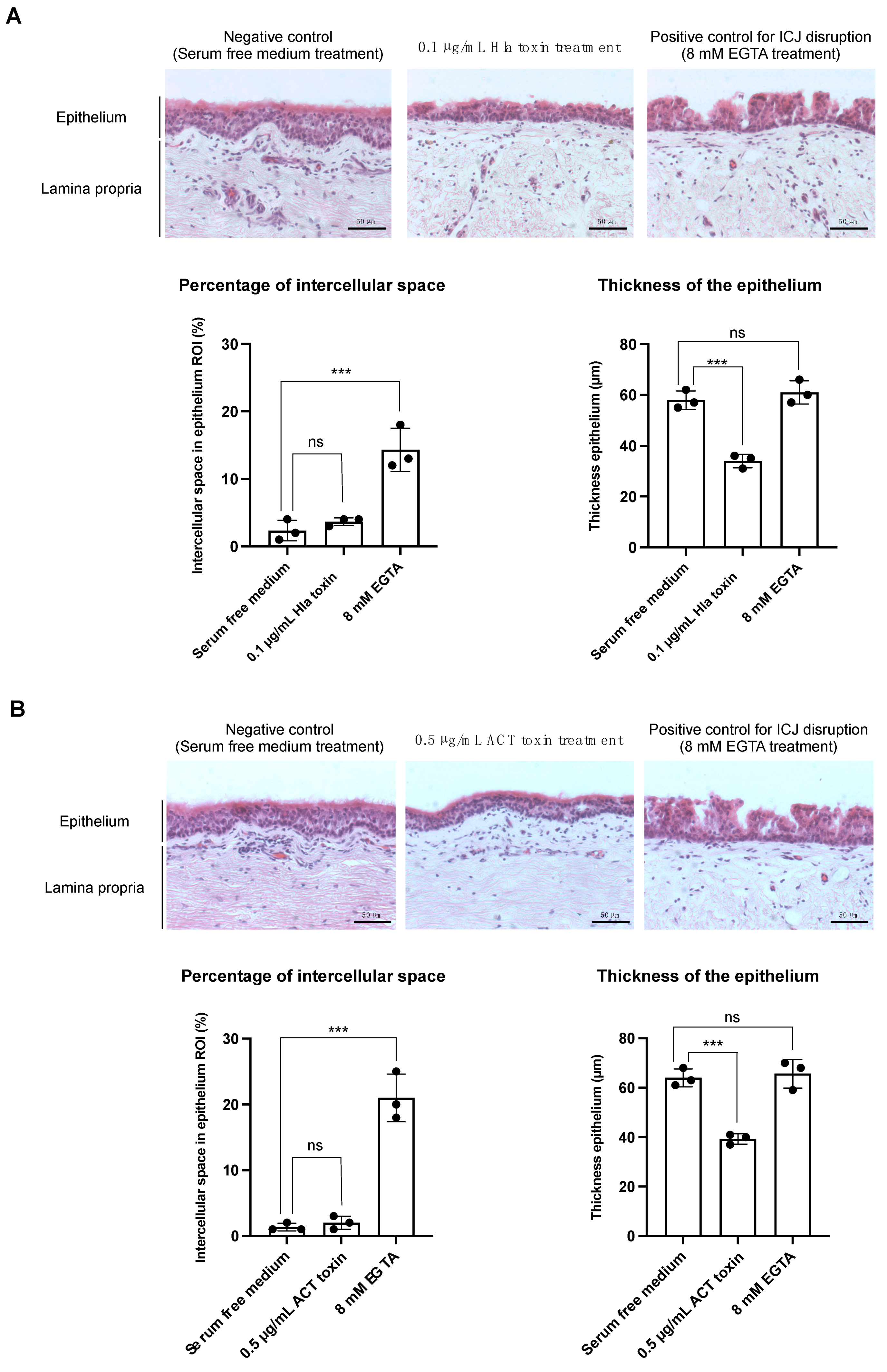
3.1. The Hla and ACT Toxins Cause Morphological Changes in the Respiratory Mucosa, Characterised by a Reduction in Epithelial Thickness
3.1.1. Treatment with the α-Hemolysin Toxin Causes a Reduction in Epithelial Thickness in Equine Respiratory Mucosal Explants
3.1.2. Treatment with the Adenylate Cyclase Toxin Causes a Reduction in Epithelial Thickness in Equine Respiratory Mucosal Explants
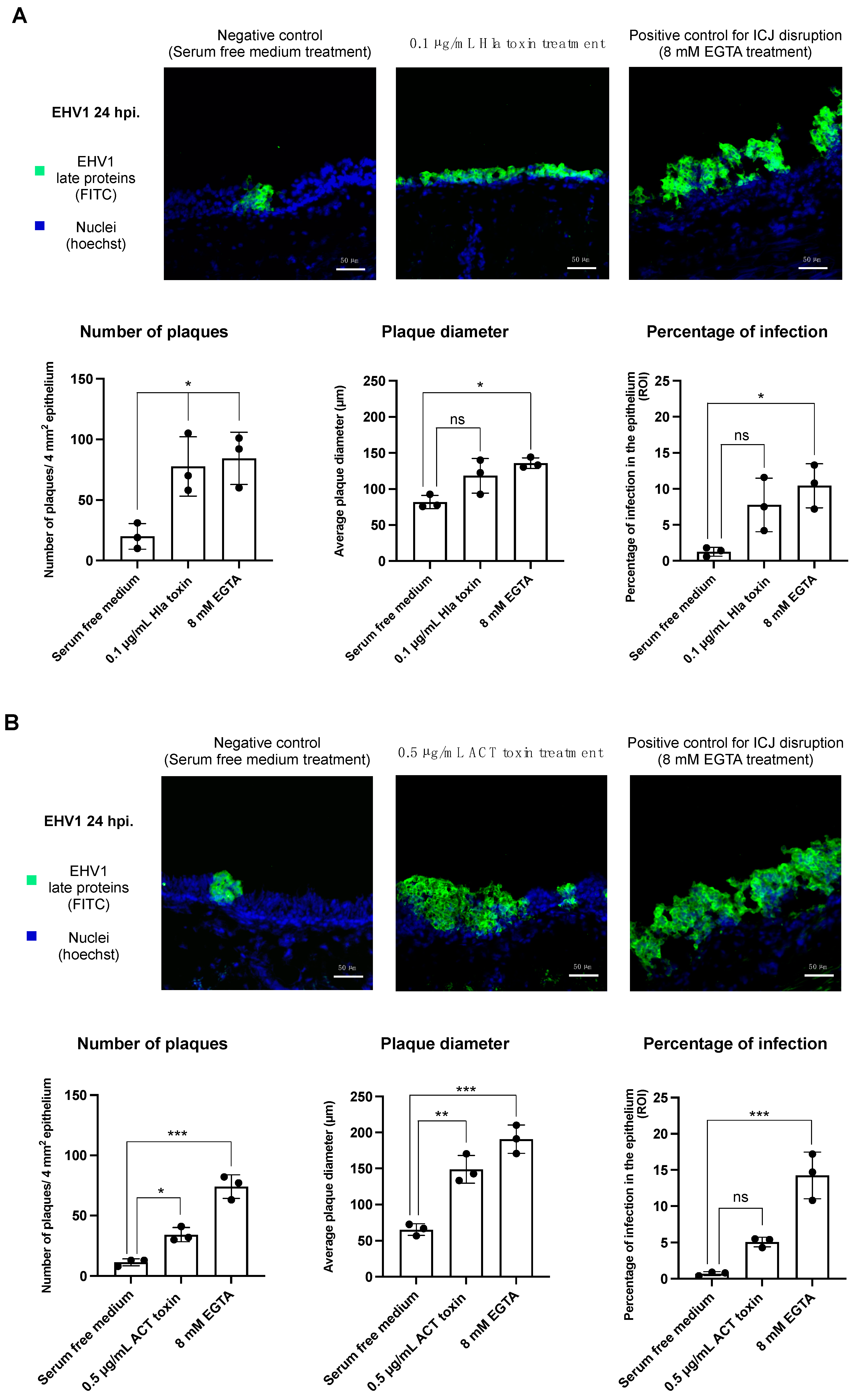
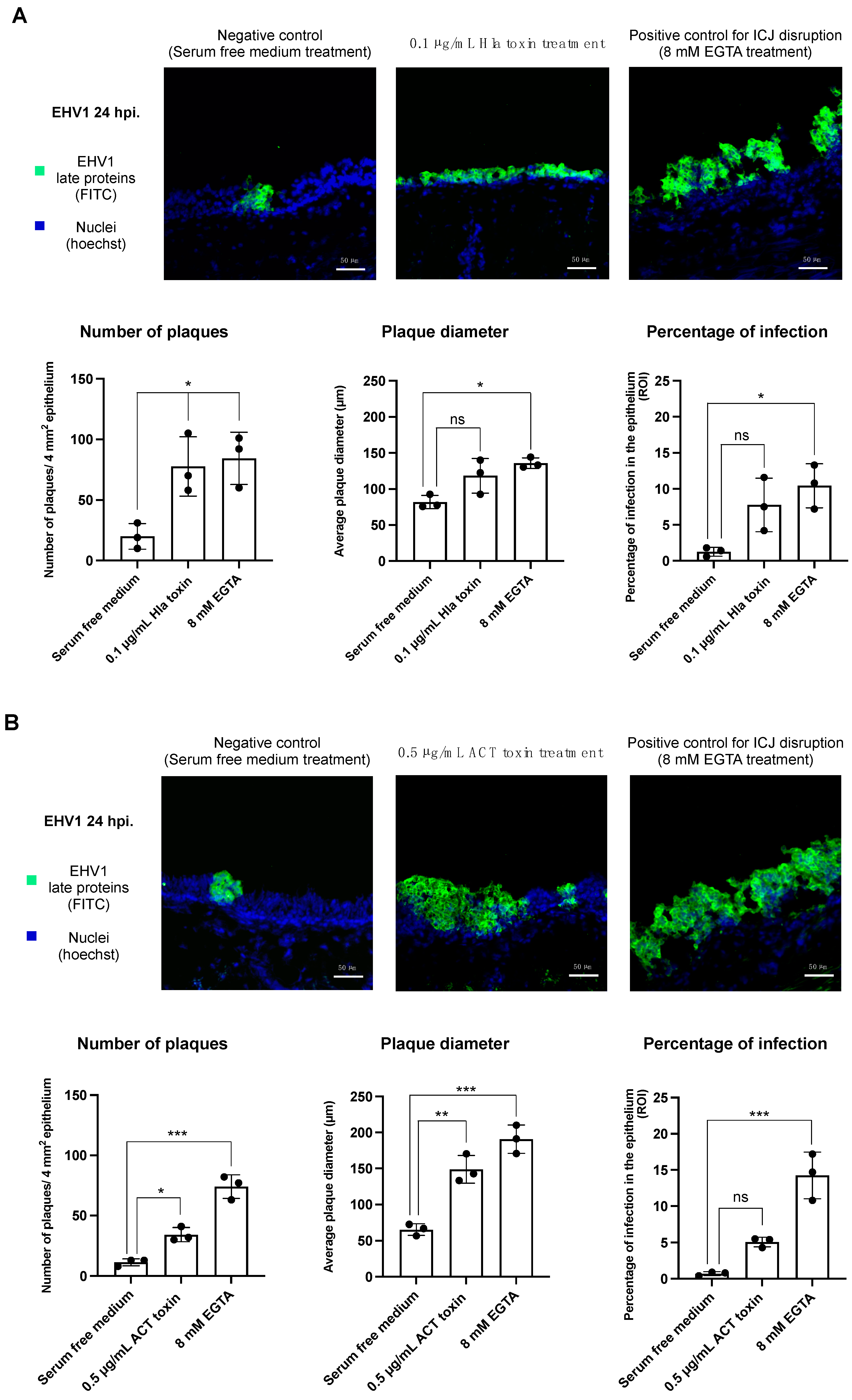
3.2. Treatment with the Hla or ACT Toxins Predisposes the Equine Respiratory Epithelium to EHV-1 Infection
3.2.1. Infection of EHV-1 in the Respiratory Epithelium following α-Hemolysin Toxin Treatment
3.2.2. Infection of EHV-1 in the Respiratory Epithelium following Adenylate Cyclase Toxin Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ainsworth, D.M.; Hackett, R.P. Disorders of the respiratory system. Equine Intern. Med. 2004, 2, 289–353. [Google Scholar]
- Van Cleemput, J.; Poelaert, K.; Laval, K.; Impens, F.; Van den Broeck, W.; Gevaert, K.; Nauwynck, H.J. Pollens destroy respiratory epithelial cell junctions and drive alphaherpesvirus infection. Sci. Rep. 2019, 9, 4787. [Google Scholar] [CrossRef]
- Van Cleemput, J.; Poelaert, K.; Laval, K.; Van den Broeck, W.; Nauwynck, H.J. Deoxynivalenol, but not fumonisin B1, afla-toxin B1 or diesel exhaust particles disrupt integrity of the horse’s respiratory epithelium and predispose it for equine her-pesvirus type 1 infection. Vet. Microbiol. 2019, 234, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Van den Eede, A.; Hermans, K.; Van den Abeele, A.; Floré, K.; Dewulf, J.; Vanderhaeghen, W.; Némeghaire, S.; Butaye, P.; Gasthuys, F.; Haesebrouck, F.; et al. The nasal vestibulum is the optimal sampling site for MRSA screening in hospitalised horses. Vet. J. 2013, 197, 415–419. [Google Scholar] [CrossRef]
- Maddox, T.W.; Scantlebury, C.E.; Clegg, P.D.; Dawson, S.; Pinchbeck, G.L.; Williams, N.J. A review of the characteristics and treatment of methicillin-resistant Staphylococcus aureus (MRSA) in the horse and a case series of MRSA infection in 4 horses. Equine Vet. Educ. 2010, 22, 91–102. [Google Scholar] [CrossRef]
- Weese, J.S.; Rousseau, J.; Traub-Dargatz, J.L.; Willey, B.M.; McGeer, A.J.; Low, D.E. Community-associated methicil-lin-resistant Staphylococcus aureus in horses and humans who work with horses. J. Am. Vet. Med. Assoc. 2005, 226, 580–583. [Google Scholar] [CrossRef]
- Hamidou Soumana, I.; Linz, B.; Harvill, E.T. Environmental Origin of the Genus Bordetella. Front. Microbiol. 2017, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cantu, M.C.; Hartmann, F.A.; Brown, B.J. Bordetella bronchiseptica and equine respiratory infections: A review of 30 cases. Equine Vet. Educ. 2000, 12, 45–50. [Google Scholar] [CrossRef]
- Vaid, R.K.; Shanmugasundaram, K.; Anand, T.; Bera, B.C.; Tigga, M.; Dedar, R.; Riyesh, T.; Bardwaj, S.; Virmani, N.; Tripathi, B.N.; et al. Characterization of isolates of Bordetella bronchiseptica from horses. J. Equine Sci. 2018, 29, 25–31. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Masin, J.; Osicka, R.; Bumba, L.; Sebo, P. Bordetella adenylate cyclase toxin: A unique combination of a pore-forming moiety with a cell-invading adenylate cyclase enzyme. Pathog. Dis. 2015, 73, ftv075. [Google Scholar] [CrossRef] [Green Version]
- Kwak, Y.K.; Vikström, E.; Magnusson, K.E.; Vécsey-Semjén, B.; Colque-Navarro, P.; Möllby, R. The Staphylococcus aureus alpha-toxin perturbs the barrier function in Caco-2 epithelial cell monolayers by altering junctional integrity. Infect. Immun. 2012, 80, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Seilie, E.S.; Bubeck Wardenburg, J. Staphylococcus aureus pore-forming toxins: The interface of pathogen and host com-plexity. Semin. Cell Dev. Biol. 2017, 72, 101–116. [Google Scholar]
- Ziesemer, S.; Eiffler, I.; Schönberg, A.; Müller, C.; Hochgräfe, F.; Beule, A.G.; Hildebrandt, J.P. Staphylococcus aureus α-Toxin Induces Actin Filament Remodeling in Human Airway Epithelial Model Cells. Am. J. Respir. Cell Mol. Biol. 2018, 58, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Kulkarni, N.N.; Asbjarnarson, A.; Linhartova, I.; Osicka, R.; Sebo, P.; Gudmundsson, G.H. Bordetella pertussis Adenylate Cyclase Toxin Disrupts Functional Integrity of Bronchial Epithelial Layers. Infect. Immun. 2018, 86, e00445-17. [Google Scholar] [CrossRef] [Green Version]
- Laval, K.; Poelaert, K.C.K.; Van Cleemput, J.; Zhao, J.; Vandekerckhove, A.P.; Gryspeerdt, A.C.; Garré, B.; van der Meulen, K.; Baghi, H.B.; Dubale, H.N.; et al. The Pathogenesis and Immune Evasive Mechanisms of Equine Herpesvirus Type 1. Front. Microbiol. 2021, 12, 662686. [Google Scholar] [CrossRef]
- Kydd, J.H.; Smith, K.C.; Hannant, D.; Livesay, G.J.; Mumford, J.A. Distribution of equid herpesvirus-1 (EHV-1) in respiratory tract associated lymphoid tissue: Implications for cellular immunity. Equine Vet. J. 1994, 26, 470–473. [Google Scholar] [CrossRef]
- Gryspeerdt, A.C.; Vandekerckhove, A.P.; Garré, B.; Barbé, F.; Van de Walle, G.R.; Nauwynck, H.J. Differences in replication kinetics and cell tropism between neurovirulent and non-neurovirulent EHV1 strains during the acute phase of infection in horses. Vet. Microbiol. 2010, 142, 242–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandekerckhove, A.P.; Glorieux, S.; Gryspeerdt, A.C.; Steukers, L.; Duchateau, L.; Osterrieder, N.; Van de Walle, G.R.; Nauwynck, H.J. Replication kinetics of neurovirulent versus non-neurovirulent equine herpesvirus type 1 strains in equine nasal mucosal explants. J. Gen. Virol. 2010, 91 Pt 8, 2019–2028. [Google Scholar] [CrossRef] [PubMed]
- Baghi, H.B.; Laval, K.; Favoreel, H.; Nauwynck, H.J. Isolation and characterization of equine nasal mucosal CD172a + cells. Vet. Immunol. Immunopathol. 2014, 157, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Allen, G. Advances in understanding of the pathogenesis, epidemiology and immunological control of equine herpesvirus abortion. In Equine Infectious Diseases: Proceedings of the Eighth International Conference; Nakajima, H., Plowright, W., Eds.; Newmarket: R&W Publishers: Suffolk, UK, 1999; pp. 129–146. [Google Scholar]
- Wilson, W.D. Equine herpesvirus 1 myeloencephalopathy. Vet. Clin. N. Am. Equine Pract. 1997, 13, 53–72. [Google Scholar] [CrossRef]
- Grinde, B. Herpesviruses: Latency and reactivation—Viral strategies and host response. J. Oral Microbiol. 2013, 5, 22766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slater, J.D.; Borchers, K.; Thackray, A.M.; Field, H.J. The trigeminal ganglion is a location for equine herpesvirus 1 latency and reactivation in the horse. J. Gen. Virol. 1994, 75 Pt 8, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Van Cleemput, J.; Poelaert, K.; Laval, K.; Maes, R.; Hussey, G.S.; Van den Broeck, W.; Nauwynck, H.J. Access to a main alphaherpesvirus receptor, located basolaterally in the respiratory epithelium, is masked by ICJ. Sci. Rep. 2017, 7, 16656. [Google Scholar] [CrossRef] [Green Version]
- Vandekerckhove, A.; Glorieux, S.; Broeck, W.V.; Gryspeerdt, A.; van der Meulen, K.M.; Nauwynck, H.J. In vitro culture of equine respiratory mucosa explants. Vet. J. 2009, 181, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Vairo, S.; Van den Broeck, W.; Favoreel, H.; Scagliarini, A.; Nauwynck, H. Development and use of a polarized equine upper respiratory tract mucosal explant system to study the early phase of pathogenesis of a European strain of equine arteritis virus. Vet. Res. 2013, 44, 22. [Google Scholar] [CrossRef] [Green Version]
- van Der Meulen, K.M.; Nauwynck, H.J.; Buddaert, W.; Pensaert, M.B. Replication of equine herpesvirus type 1 in freshly isolated equine peripheral blood mononuclear cells and changes in susceptibility following mitogen stimulation. J. Gen. Virol. 2000, 81 Pt 1, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Laval, K.; Favoreel, H.W.; Poelaert, K.C.; Van Cleemput, J.; Nauwynck, H.J. Equine Herpesvirus Type 1 Enhances Viral Replication in CD172a+ Monocytic Cells upon Adhesion to Endothelial Cells. J. Virol. 2015, 89, 10912–10923. [Google Scholar] [CrossRef] [Green Version]
- van der Meulen, K.M.; Nauwynck, H.J.; Pensaert, M.B. Absence of viral antigens on the surface of equine herpesvi-rus-1-infected peripheral blood mononuclear cells: A strategy to avoid complement-mediated lysis. J. Gen. Virol. 2003, 84 Pt 1, 93–97. [Google Scholar] [CrossRef]
- Yang, X.; Forier, K.; Steukers, L.; Van Vlierberghe, S.; Dubruel, P.; Braeckmans, K.; Glorieux, S.; Nauwynck, H.J. Immobili-zation of pseudorabies virus in porcine tracheal respiratory mucus revealed by single particle tracking. PLoS ONE 2012, 7, e51054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beadling, C.; Slifka, M.K. How do viral infections predispose patients to bacterial infections? Curr. Opin. Infect. Dis. 2004, 17, 185–191. [Google Scholar] [CrossRef]
- Sender, V.; Hentrich, K.; Henriques-Normark, B. Virus-Induced Changes of the Respiratory Tract Environment Promote Secondary Infections With Streptococcus pneumoniae. Front. Cell Infect. Microbiol. 2021, 11, 643326. [Google Scholar] [CrossRef] [PubMed]
- Bin, L.; Kim, B.E.; Brauweiler, A.; Goleva, E.; Streib, J.; Ji, Y.; Schlievert, P.M.; Leung, D.Y. Staphylococcus aureus α-toxin modulates skin host response to viral infection. J. Allergy Clin. Immunol. 2012, 130, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, H.; Miyake, M.; Kamitani, S.; Horiguchi, Y. The morphological changes in cultured cells caused by Bordetella pertussis adenylate cyclase toxin. FEMS Microbiol. Lett. 2008, 279, 174–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeil, E.; Capaldo, C.T.; Macara, I.G. Zonula occludens-1 function in the assembly of tight junctions in Madin-Darby canine kidney epithelial cells. Mol. Biol. Cell 2006, 17, 1922–1932. [Google Scholar] [CrossRef] [Green Version]
- Stevens Lowe, J.S.; Anderson, P.G. Chapter 3—Epithelial Cells. In Human Histology, 4th ed.; Mosby Elsevier: Maryland Heights, MO, USA, 2005; pp. 37–54. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Crombrugge, E.; Vanbeylen, E.; Van Cleemput, J.; Van den Broeck, W.; Laval, K.; Nauwynck, H. Bacterial Toxins from Staphylococcus aureus and Bordetella bronchiseptica Predispose the Horse’s Respiratory Tract to Equine Herpesvirus Type 1 Infection. Viruses 2022, 14, 149. https://doi.org/10.3390/v14010149
Van Crombrugge E, Vanbeylen E, Van Cleemput J, Van den Broeck W, Laval K, Nauwynck H. Bacterial Toxins from Staphylococcus aureus and Bordetella bronchiseptica Predispose the Horse’s Respiratory Tract to Equine Herpesvirus Type 1 Infection. Viruses. 2022; 14(1):149. https://doi.org/10.3390/v14010149
Chicago/Turabian StyleVan Crombrugge, Eline, Emma Vanbeylen, Jolien Van Cleemput, Wim Van den Broeck, Kathlyn Laval, and Hans Nauwynck. 2022. "Bacterial Toxins from Staphylococcus aureus and Bordetella bronchiseptica Predispose the Horse’s Respiratory Tract to Equine Herpesvirus Type 1 Infection" Viruses 14, no. 1: 149. https://doi.org/10.3390/v14010149
APA StyleVan Crombrugge, E., Vanbeylen, E., Van Cleemput, J., Van den Broeck, W., Laval, K., & Nauwynck, H. (2022). Bacterial Toxins from Staphylococcus aureus and Bordetella bronchiseptica Predispose the Horse’s Respiratory Tract to Equine Herpesvirus Type 1 Infection. Viruses, 14(1), 149. https://doi.org/10.3390/v14010149