
Detection of Enterovirus D68 in Wastewater Samples from the UK between July and November 2021

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wastewater Sample Collection and Processing
2.2. Modified Pan-EV Entire-Capsid Coding Region RT-PCR Amplification (mECRA)
- Primers 5′NCR (5′-TGGCGGAACCGACTACTTTGGGTG-3′) and CRE-R (5′-TCAATACGGTGTTTGCTCTTGAACTG-3′);
- Primers MM_EV_F2 (5′-CAGCGGAACCGACTACTTT-3′) and MM_EV_R1 (5′-AATACGGCATTTGGACTTGAACTGT-3′).
2.3. VP1-Nested PCR Amplification and Nucleotide Sequence Analysis of the VP1 Coding Region of EV-D68 Strains by Sanger Sequencing
- Universal primers: D68_VP1_PubF (5′-ACCATTTACATGCRGCAGAGG-3′) and D68-2ARpub (5′-ACATCTGAYTGCCARTCYAC-3′);
- Clade B3 primers: D68_VP1_PubF (5′-ACCATTTACATGCRGCAGAGG-3′) and 2016_D68_VP1R (5′-CCTGGACCAGTAGTCACTATATTATG-3′);
- Clade D primers: 14.001-D68-VP1F (5′-ATTGGACAACTAGAGCATTTACATGAG-3′) and 14.001-D68-VP1R (5′-CAGTATTCACTAACCGAATGTCGTG-3′).
2.4. NGS Analysis of mECRA Products
2.5. Phylogenetic Analysis of EV-D68 Strains from Wastewater Samples
3. Results
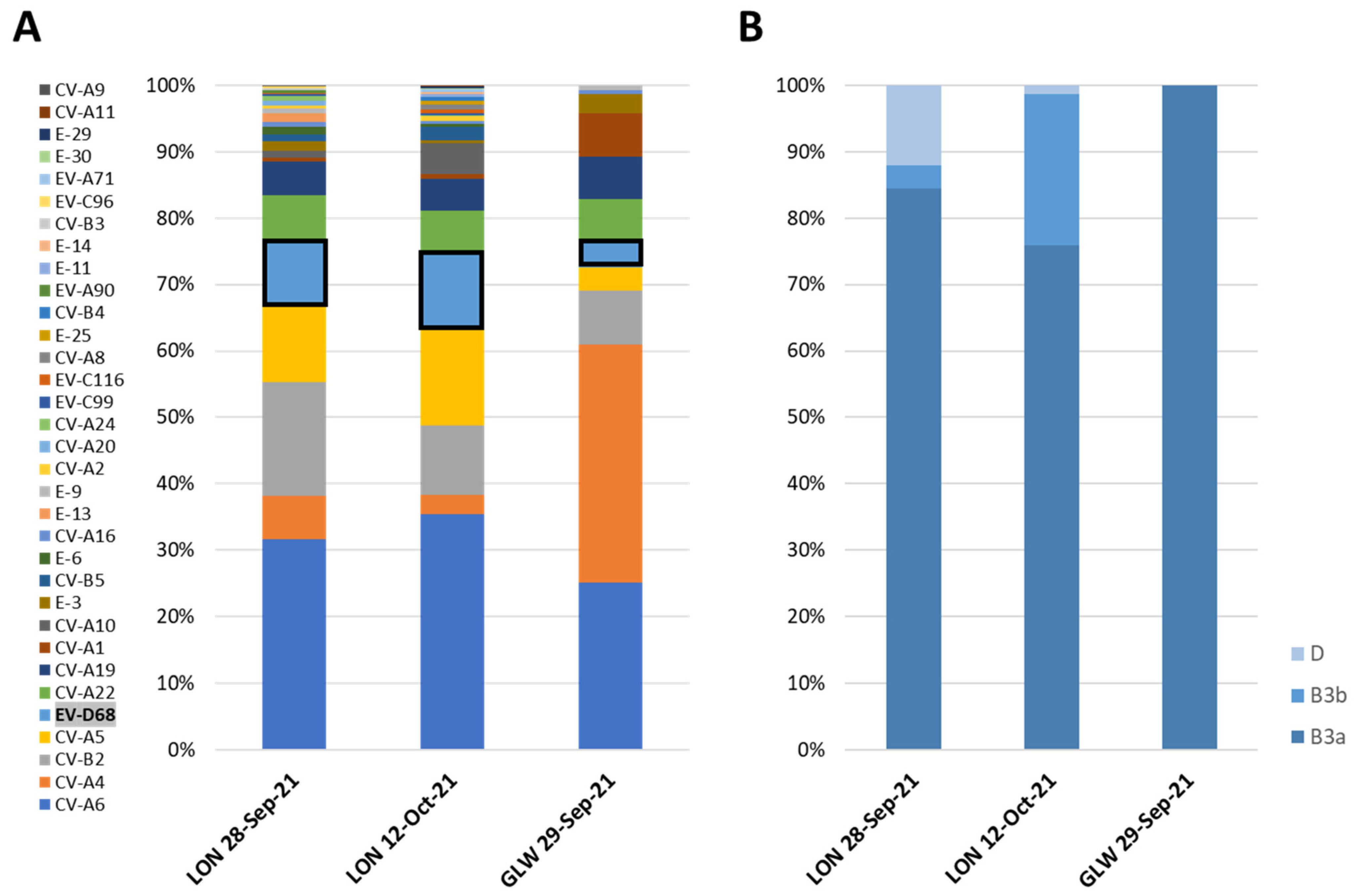
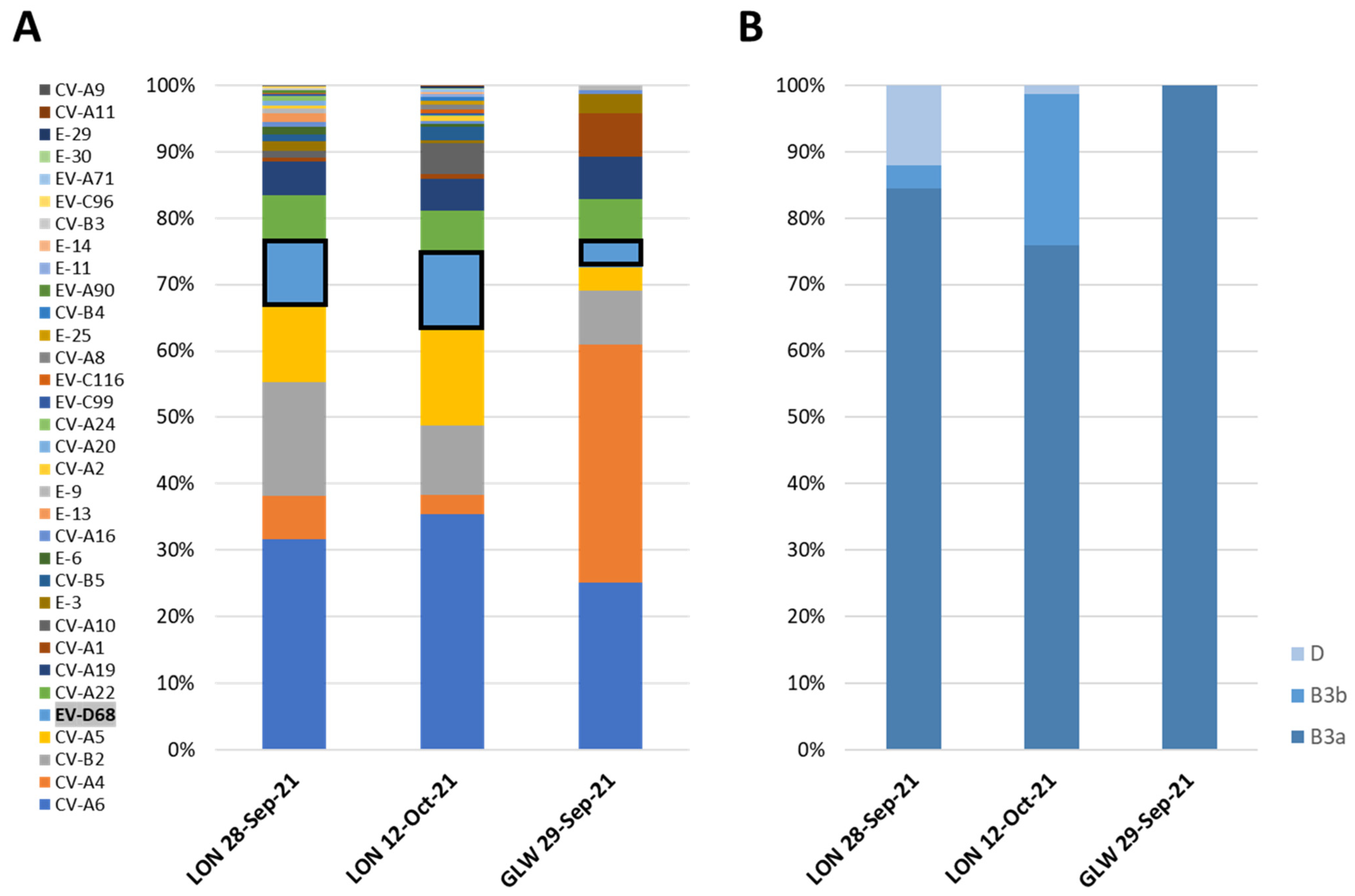
3.1. Detection of EV-D68 in Wastewater Samples by VP1-Nested PCR and Nucleotide Sequence Analysis

{kind=link}
{kind=link}
| VP1-Nested PCR Positive for Enterovirus D68 (%) 1 | ||||
|---|---|---|---|---|
| Location | Date of Collection | Universal Primers | Clade B3 Primers | Clade D Primers |
| London | 13 July 2021 | 0 | 16.7 | 0 |
| Glasgow | 29 July 2021 | 16.7 | 83.3 | 0 |
| London | 10 August 2021 | 33.3 | 50 | 0 |
| Glasgow | 26 August 2021 | 83.3 | 33.3 | 0 |
| London | 14 September 2021 | 50 | 50 | 0 |
| London | 28 September 2021 | 83.3 | 100 | 33.3 |
| Glasgow | 29 September 2021 | 66.7 | 100 | 0 |
| London | 12 October 2021 | 100 | 100 | 50 |
| London | 26 October 2021 | 83.3 | 100 | 50 |
| Glasgow | 27 October 2021 | 66.7 | 100 | 83.3 |
| London | 23 November 2021 | 100 | 100 | 0 |
| Glasgow | 24 November 2021 | 100 | 100 | 33.3 |
3.2. NGS Analysis of EV-D68 VP1 Sequences from Wastewater Samples
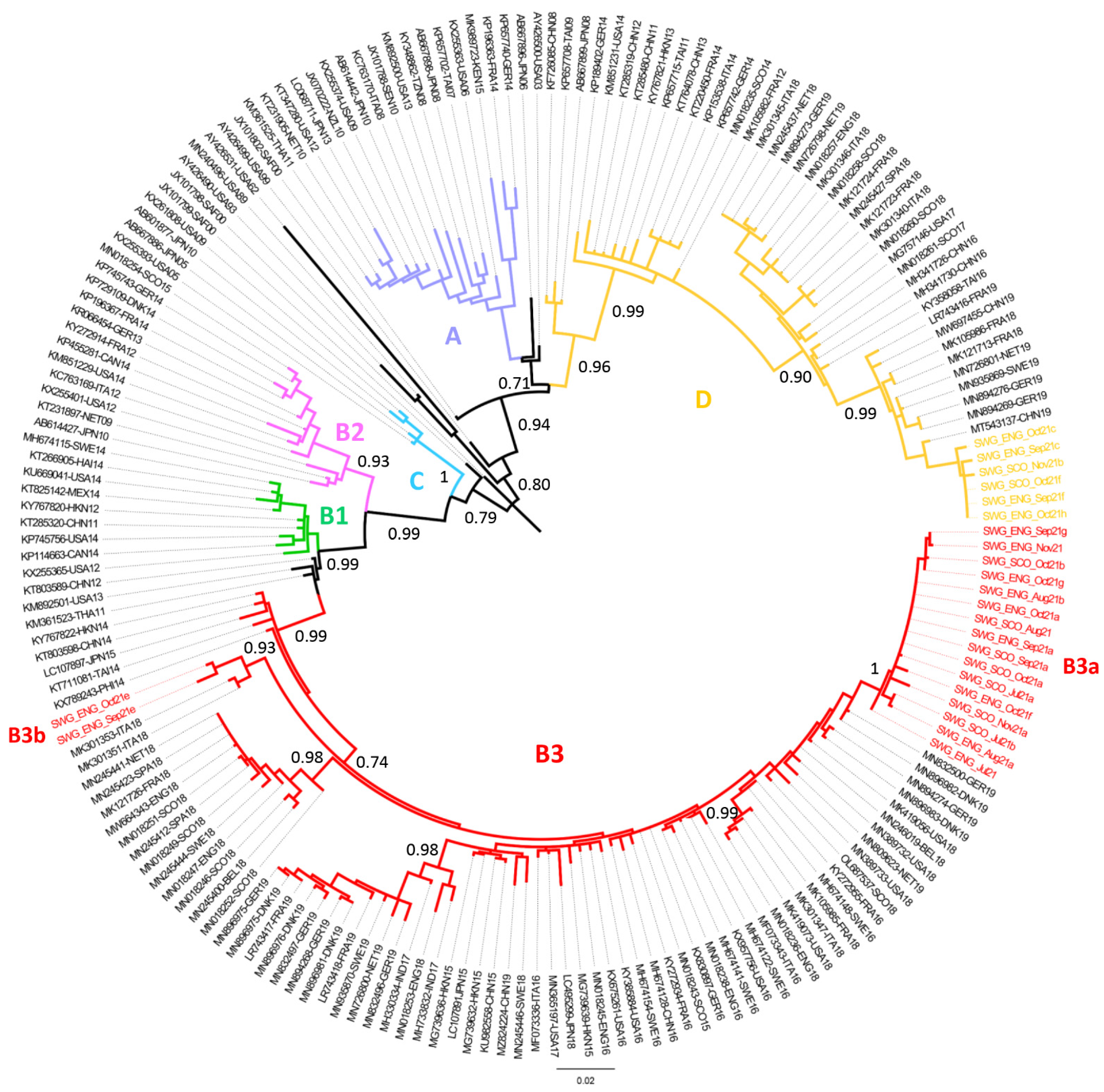
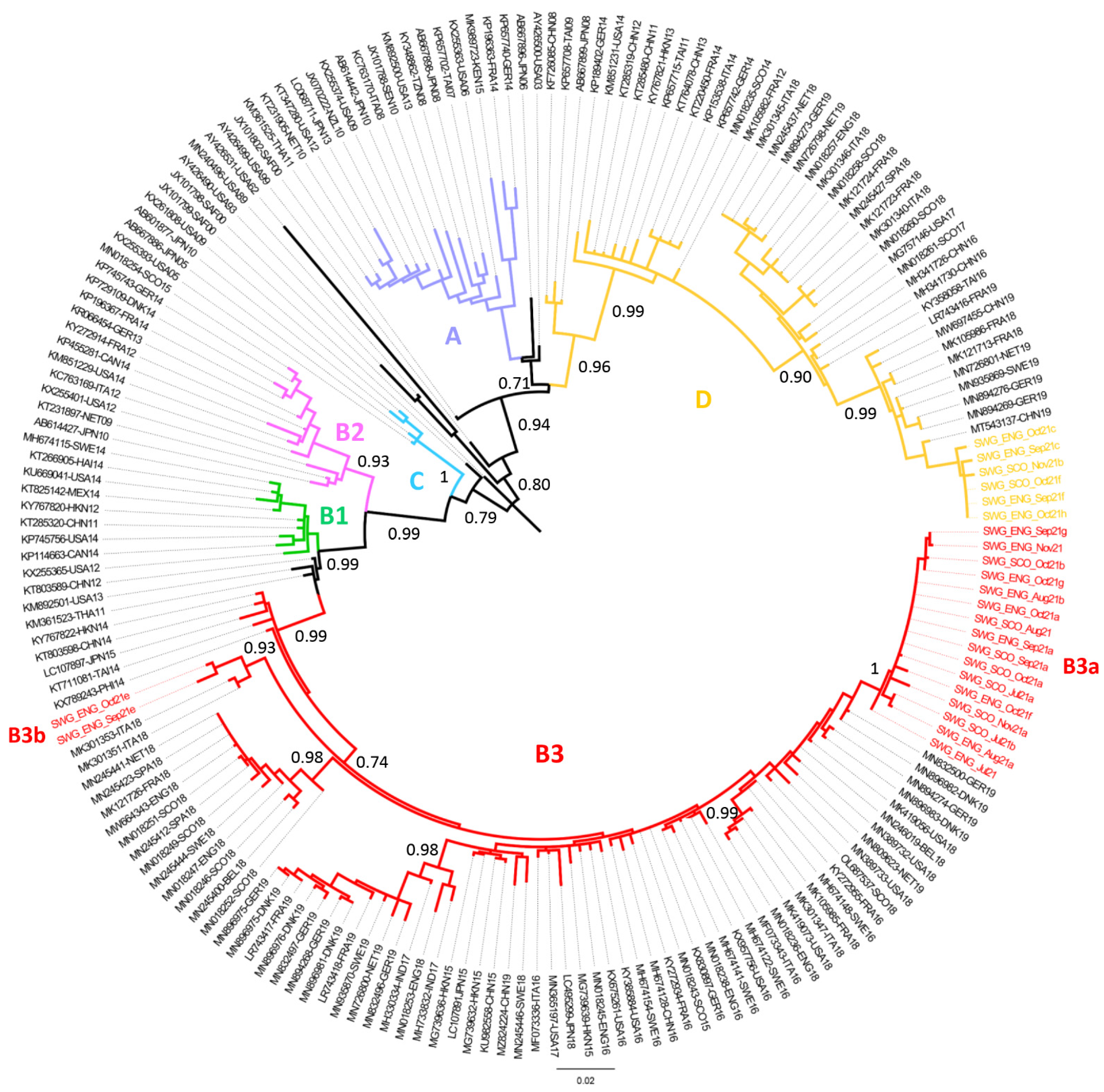
3.3. Phylogenetic Analysis of EV-D68 Strains Identified in UK Wastewater Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EV | Enterovirus |
| E- | Echovirus |
| CV- | Coxsackievirus |
| NGS | Next-Generation Sequencing |
| ES | Environmental Surveillance |
| ORF | Open Reading Frame |
| Country Abbreviations | USA, United States; NET, Netherland; IND, India; JPN, Japan; CHN, China; THA, Thailand; GER, Germany; MAL, Malaysia; DNK, Denmark; TAI, Taiwan; FRA, France; HKN, Hong Kong; SAF, South Africa; SEN, Senegal; SWE, Sweden; ITA, Italy; MEX, Mexico; PHI, Philippines; SPA, Spain; CAN, Canada; KEN, Kenya; SCO, Scotland; ENG, England; HAI, Haiti; TZN, Tanzania; NZL, New Zealand. |
References
- Cassidy, H.; Poelman, R.; Knoester, M.; Van Leer-Buter, C.C.; Niesters, H.G.M. Enterovirus D68—The New Polio? Front. Microbiol. 2018, 9, 2677. [Google Scholar] [CrossRef]
- Messacar, K.; Asturias, E.J.; Hixon, A.M.; Van Leer-Buter, C.; Niesters, H.G.M.; Tyler, K.L.; Abzug, M.J.; Dominguez, S.R. Enterovirus D68 and acute flaccid myelitis—Evaluating the evidence for causality. Lancet Infect. Dis. 2018, 18, e239–e247. [Google Scholar] [CrossRef] [Green Version]
- Dyda, A.; Stelzer-Braid, S.; Adam, D.; A Chughtai, A.; MacIntyre, C.R. The association between acute flaccid myelitis (AFM) and Enterovirus D68 (EV-D68)—What is the evidence for causation? Eurosurveillance 2018, 23, 17-00310. [Google Scholar] [CrossRef] [Green Version]
- Kramer, R.; Sabatier, M.; Wirth, T.; Pichon, M.; Lina, B.; Schuffenecker, I.; Josset, L. Molecular diversity and biennial circulation of enterovirus D68: A systematic screening study in Lyon, France, 2010 to 2016. Eurosurveillance 2018, 23, 1700711. [Google Scholar] [CrossRef] [Green Version]
- Messacar, K.; Pretty, K.; Reno, S.; Dominguez, S.R. Continued biennial circulation of enterovirus D68 in Colorado. J. Clin. Virol. 2019, 113, 24–26. [Google Scholar] [CrossRef]
- Holm-Hansen, C.C.; Midgley, S.E.; Fischer, T.K. Global emergence of enterovirus D68: A systematic review. Lancet Infect. Dis. 2016, 16, e64–e75. [Google Scholar] [CrossRef]
- Kidd, S.; Lopez, A.S.; Konopka-Anstadt, J.L.; Nix, W.A.; Routh, J.A.; Oberste, M.S. Enterovirus D68–Associated Acute Flaccid Myelitis, United States, 2020. Emerg. Infect. Dis. 2020, 26, 26. [Google Scholar] [CrossRef]
- The United Kingdom Acute Flaccid Paralysis (Afp) Task Force. An increase in reports of acute flaccid paralysis (AFP) in the United Kingdom, 1 January 2018–21 January 2019: Early findings. Eurosurveillance 2019, 24, 1900093. [Google Scholar] [CrossRef] [Green Version]
- Midgley, S.E.; Benschop, K.; Dyrdak, R.; Mirand, A.; Bailly, J.-L.; Bierbaum, S.; Buderus, S.; Böttcher, S.; Eis-Hübinger, A.-M.; Hönemann, M.; et al. Co-circulation of multiple enterovirus D68 subclades, including a novel B3 cluster, across Europe in a season of expected low prevalence, 2019/20. Eurosurveillance 2020, 25, 1900749. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Pons-Salort, M.; Messacar, K.; Cook, C.; Meyers, L.; Farrar, J.; Grenfell, B.T. Epidemiological dynamics of enterovirus D68 in the United States and implications for acute flaccid myelitis. Sci. Transl. Med. 2021, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.J.; Winn, A.K.; Budd, A.P.; Prill, M.M.; Steel, J.; Midgley, C.M.; Kniss, K.; Burns, E.; Rowe, T.; Foust, A.; et al. Changes in Influenza and Other Respiratory Virus Activity During the COVID-19 Pandemic—United States, 2020–2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Stoffel, L.; A Agyeman, P.K.; Keitel, K.; Barbani, M.T.; Duppenthaler, A.; Kopp, M.V.; Aebi, C. Striking Decrease of Enteroviral Meningitis in Children During the COVID-19 Pandemic. Open Forum Infect. Dis. 2021, 8, ofab115. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.; Perez, A.; Lively, J.Y.; Avadhanula, V.; Boom, J.A.; Chappell, J.; Englund, J.A.; Fregoe, W.; Halasa, N.B.; Harrison, C.J.; et al. Enterovirus D68-Associated Acute Respiratory Illness—New Vaccine Surveillance Network, United States, July–November 2018–2020. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 1623–1628. [Google Scholar] [CrossRef] [PubMed]
- Benschop, K.S.; Albert, J.; Anton, A.; Andrés, C.; Aranzamendi, M.; Armannsdóttir, B.; Bailly, J.-L.; Baldanti, F.; Baldvinsdóttir, G.E.; Beard, S.; et al. Re-emergence of enterovirus D68 in Europe after easing the COVID-19 lockdown, September 2021. Eurosurveillance 2021, 26, 2100998. [Google Scholar] [CrossRef]
- Asghar, H.; Diop, O.M.; Weldegebriel, G.; Malik, F.; Shetty, S.; El Bassioni, L.; Akande, A.O.; Al Maamoun, E.; Zaidi, S.; Adeniji, A.J.; et al. Environmental Surveillance for Polioviruses in the Global Polio Eradication Initiative. J. Infect. Dis. 2014, 210, S294–S303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraro, G.B.; Veneri, C.; Mancini, P.; Iaconelli, M.; Suffredini, E.; Bonadonna, L.; Lucentini, L.; Bowo-Ngandji, A.; Kengne-Nde, C.; Mbaga, D.S.; et al. A State-of-the-Art Scoping Review on SARS-CoV-2 in Sewage Focusing on the Potential of Wastewater Surveillance for the Monitoring of the COVID-19 Pandemic. Food Environ. Virol. 2021, 1–40. [Google Scholar] [CrossRef]
- Weil, M.; Mandelboim, M.; Mendelson, E.; Manor, Y.; Shulman, L.; Ram, D.; Barkai, G.; Shemer, Y.; Wolf, D.; Kra-Oz, Z.; et al. Human enterovirus D68 in clinical and sewage samples in Israel. J. Clin. Virol. 2016, 86, 52–55. [Google Scholar] [CrossRef]
- Bisseux, M.; Colombet, J.; Mirand, A.; Roque-Afonso, A.-M.; Abravanel, F.; Izopet, J.; Archimbaud, C.; Peigue-Lafeuille, H.; Debroas, D.; Bailly, J.-L.; et al. Monitoring human enteric viruses in wastewater and relevance to infections encountered in the clinical setting: A one-year experiment in central France, 2014 to 2015. Eurosurveillance 2018, 23, 17-00237. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, M.; Wilton, T.; Hajarha, Y.; Klapsa, D.; Martin, J. Detection of Enterovirus D68 in wastewater samples from the United Kingdom during outbreaks reported globally between 2014 and 2018. BioRxiv 2019. [Google Scholar] [CrossRef]
- Mishra, S.; Mindermann, S.; Sharma, M.; Whittaker, C.; A Mellan, T.; Wilton, T.; Klapsa, D.; Mate, R.; Fritzsche, M.; Zambon, M.; et al. Changing composition of SARS-CoV-2 lineages and rise of Delta variant in England. EClinicalMedicine 2021, 39, 101064. [Google Scholar] [CrossRef]
- Wilton, T.; Bujaki, E.; Klapsa, D.; Majumdar, M.; Zambon, M.; Fritzsche, M.; Mate, R.; Martin, J. Rapid Increase of SARS-CoV-2 Variant B.1.1.7 Detected in Sewage Samples from England between October 2020 and January 2021. mSystems 2021, 6, e0035321. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; Klapsa, D.; Wilton, T.; Zambon, M.; Bentley, E.; Bujaki, E.; Fritzsche, M.; Mate, R.; Majumdar, M. Tracking SARS-CoV-2 in Sewage: Evidence of Changes in Virus Variant Predominance during COVID-19 Pandemic. Viruses 2020, 12, 1144. [Google Scholar] [CrossRef]
- Majumdar, M.; Klapsa, D.; Wilton, T.; Akello, J.O.; Anscombe, C.; Allen, D.; Mee, E.T.; Minor, P.D.; Martin, J. Isolation of Vaccine-Like Poliovirus Strains in Sewage Samples From the United Kingdom. J. Infect. Dis. 2018, 217, 1222–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumdar, M.; Martin, J. Detection by Direct Next Generation Sequencing Analysis of Emerging Enterovirus D68 and C109 Strains in an Environmental Sample From Scotland. Front. Microbiol. 2018, 9, 1956. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, M.; Sharif, S.; Klapsa, D.; Wilton, T.; Alam, M.M.; Garcia, M.D.F.; Rehman, L.; Mujtaba, G.; McAllister, G.; Harvala, H.; et al. Environmental Surveillance Reveals Complex Enterovirus Circulation Patterns in Human Populations. Open Forum Infect. Dis. 2018, 5, ofy250. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, M.; Klapsa, D.; Wilton, T.; Bujaki, E.; Fernandez-Garcia, M.D.; Faleye, T.O.C.; Oyero, A.O.; Adewumi, M.O.; Ndiaye, K.; Adeniji, J.A.; et al. High Diversity of Human Non-Polio Enterovirus Serotypes Identified in Contaminated Water in Nigeria. Viruses 2021, 13, 249. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.G.; Majumdar, M.; Troman, C.; O’Toole, Á.; Benny, B.; Abraham, D.; Praharaj, I.; Kang, G.; Sharif, S.; Alam, M.M.; et al. Rapid and Sensitive Direct Detection and Identification of Poliovirus from Stool and Environmental Surveillance Samples by Use of Nanopore Sequencing. J. Clin. Microbiol. 2020, 58, 00920. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.L.; Church, D.M.; Lash, A.; Leipe, D.D.; Madden, T.L.; Pontius, J.U.; Schuler, G.D.; Schriml, L.M.; Tatusova, T.A.; Wagner, L.; et al. Database resources of the National Center for Biotechnology Information: 2002 update. Nucleic Acids Res. 2002, 30, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Kroneman, A.; Vennema, H.; Deforche, K.; Avoort, H.; Peñaranda, S.; Oberste, M.; Vinje, J.; Koopmans, M. An automated genotyping tool for enteroviruses and noroviruses. J. Clin. Virol. 2011, 51, 121–125. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Imamura, T.; Okamoto, M.; Nakakita, S.-I.; Suzuki, A.; Saito, M.; Tamaki, R.; Lupisan, S.; Roy, C.N.; Hiramatsu, H.; Sugawara, K.-E.; et al. Antigenic and Receptor Binding Properties of Enterovirus 68. J. Virol. 2014, 88, 2374–2384. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- UK Health Security Agency. Enterovirus: Summary of Strain Characterisation. 2021. Available online: https://www.gov.uk/government/publications/enterovirus-strain-characterisation/enterovirus-summary-of-strain-characterisation (accessed on 6 December 2021).
- Brinkman, N.E.; Fout, G.S.; Keely, S. Retrospective Surveillance of Wastewater To Examine Seasonal Dynamics of Enterovirus Infections. mSphere 2017, 2, e00099-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisseux, M.; Debroas, D.; Mirand, A.; Archimbaud, C.; Peigue-Lafeuille, H.; Bailly, J.-L.; Henquell, C. Monitoring of enterovirus diversity in wastewater by ultra-deep sequencing: An effective complementary tool for clinical enterovirus surveillance. Water Res. 2020, 169, 115246. [Google Scholar] [CrossRef] [PubMed]
- Harvala, H.; Calvert, J.; Van Nguyen, D.; Clasper, L.; Gadsby, N.; Molyneaux, P.; Templeton, K.; McWilliams Leitch, C.; Simmonds, P. Comparison of diagnostic clinical samples and environmental sampling for enterovirus and parechovirus surveillance in Scotland, 2010 to 2012. Eurosurveillance 2014, 19, 20772. [Google Scholar] [CrossRef] [Green Version]
- Solomon, T.; Lewthwaite, P.; Perera, D.; Cardosa, M.J.; McMinn, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect. Dis. 2010, 10, 778–790. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedcastle, A.; Wilton, T.; Pegg, E.; Klapsa, D.; Bujaki, E.; Mate, R.; Fritzsche, M.; Majumdar, M.; Martin, J. Detection of Enterovirus D68 in Wastewater Samples from the UK between July and November 2021. Viruses 2022, 14, 143. https://doi.org/10.3390/v14010143
Tedcastle A, Wilton T, Pegg E, Klapsa D, Bujaki E, Mate R, Fritzsche M, Majumdar M, Martin J. Detection of Enterovirus D68 in Wastewater Samples from the UK between July and November 2021. Viruses. 2022; 14(1):143. https://doi.org/10.3390/v14010143
Chicago/Turabian StyleTedcastle, Alison, Thomas Wilton, Elaine Pegg, Dimitra Klapsa, Erika Bujaki, Ryan Mate, Martin Fritzsche, Manasi Majumdar, and Javier Martin. 2022. "Detection of Enterovirus D68 in Wastewater Samples from the UK between July and November 2021" Viruses 14, no. 1: 143. https://doi.org/10.3390/v14010143
APA StyleTedcastle, A., Wilton, T., Pegg, E., Klapsa, D., Bujaki, E., Mate, R., Fritzsche, M., Majumdar, M., & Martin, J. (2022). Detection of Enterovirus D68 in Wastewater Samples from the UK between July and November 2021. Viruses, 14(1), 143. https://doi.org/10.3390/v14010143