
Comparative Analysis of Novel Strains of Porcine Astrovirus Type 3 in the USA

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Diagnostic Samples
2.2. RT-qPCR and Sanger Sequencing
2.3. Phylogenic and Sequence Analysis
2.4. UTRs and Frameshift Signal Analysis
2.5. Identification of Putative Genome-Linked Virus Protein (VPg)
2.6. Linear Antigen Epitope Prediction
3. Results
3.1. Diagnostic Samples
3.2. RT-qPCR and Sanger Sequencing
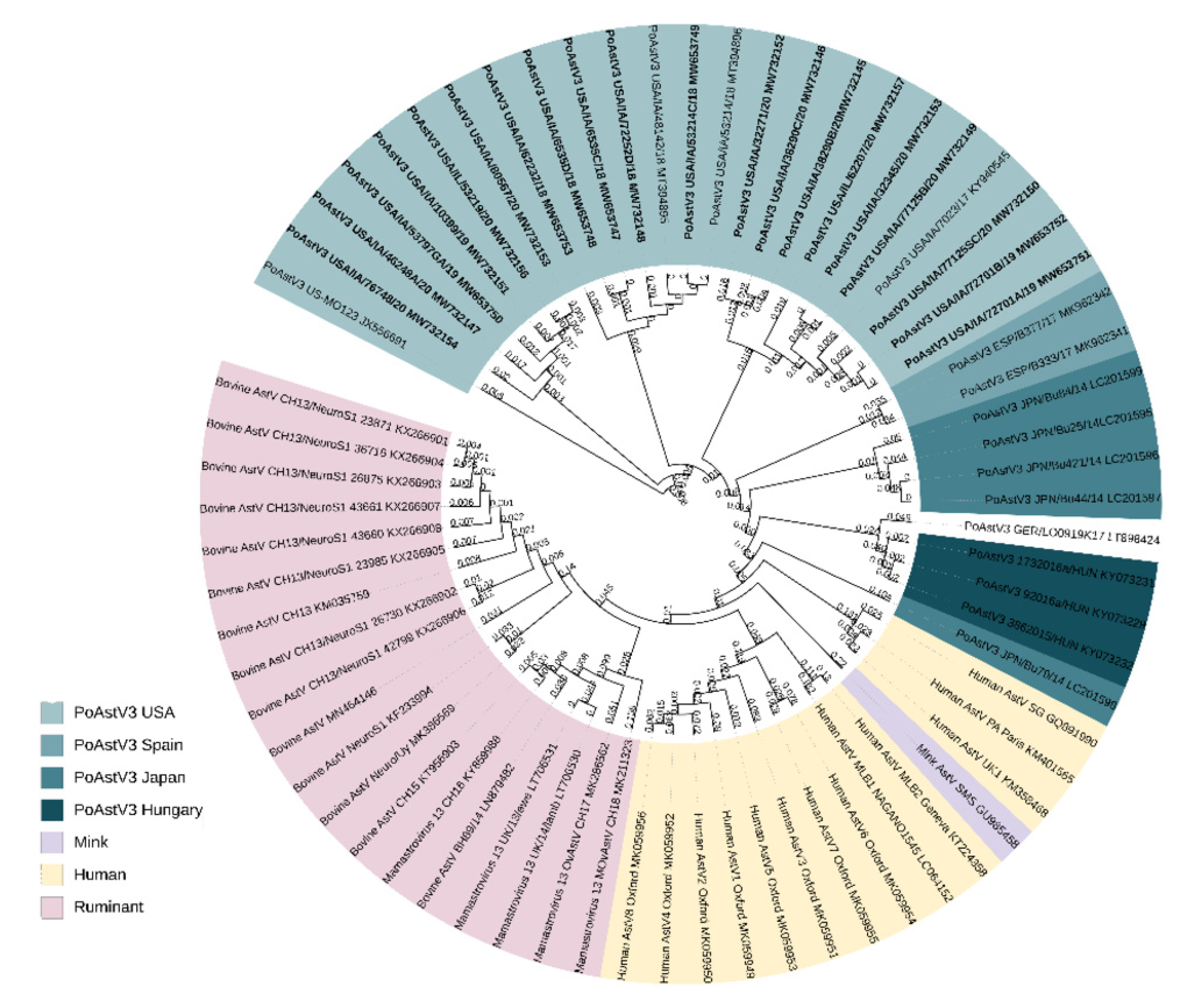
3.3. Phylogenic and Sequence Analysis
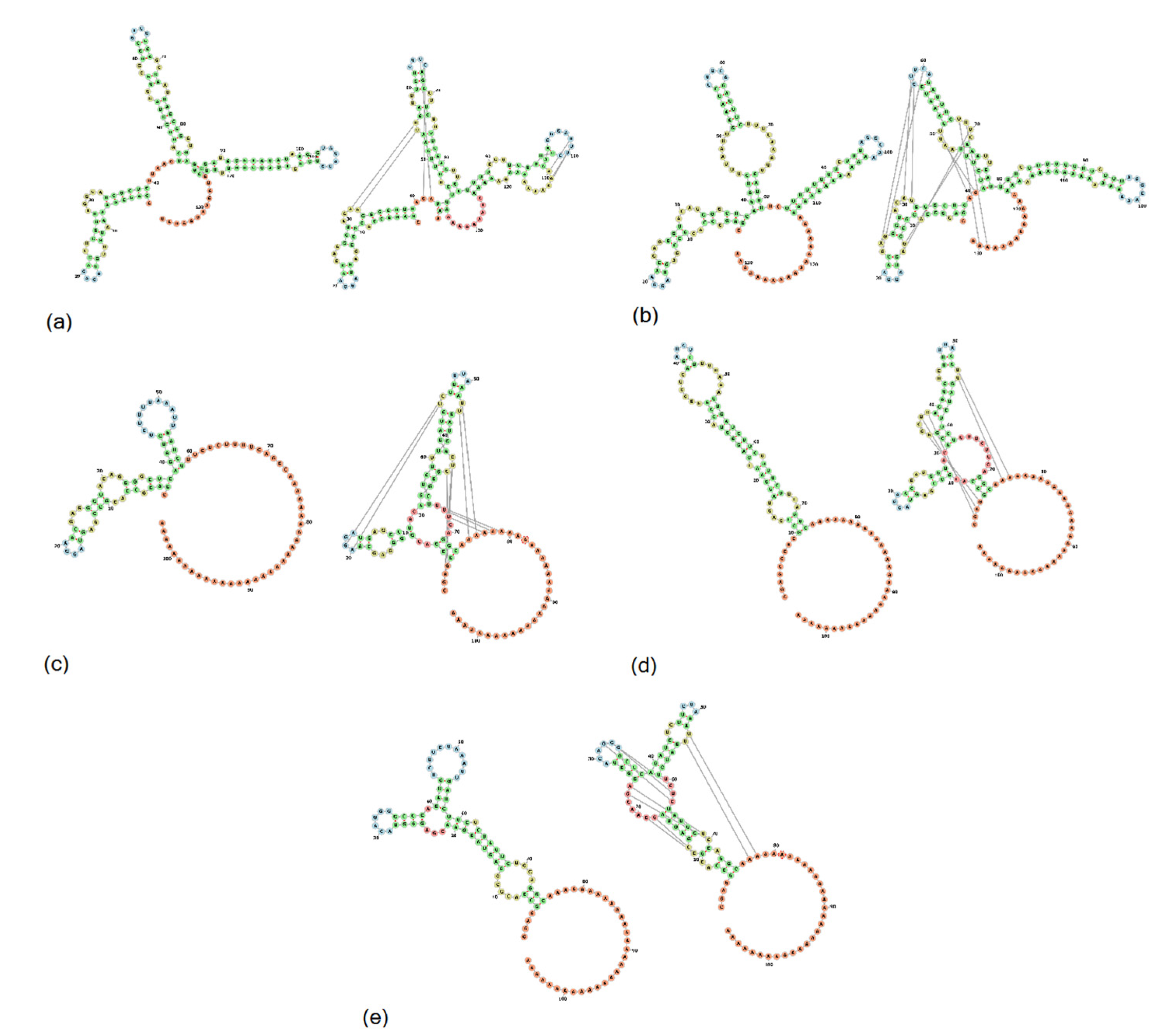
3.4. UTR Analysis
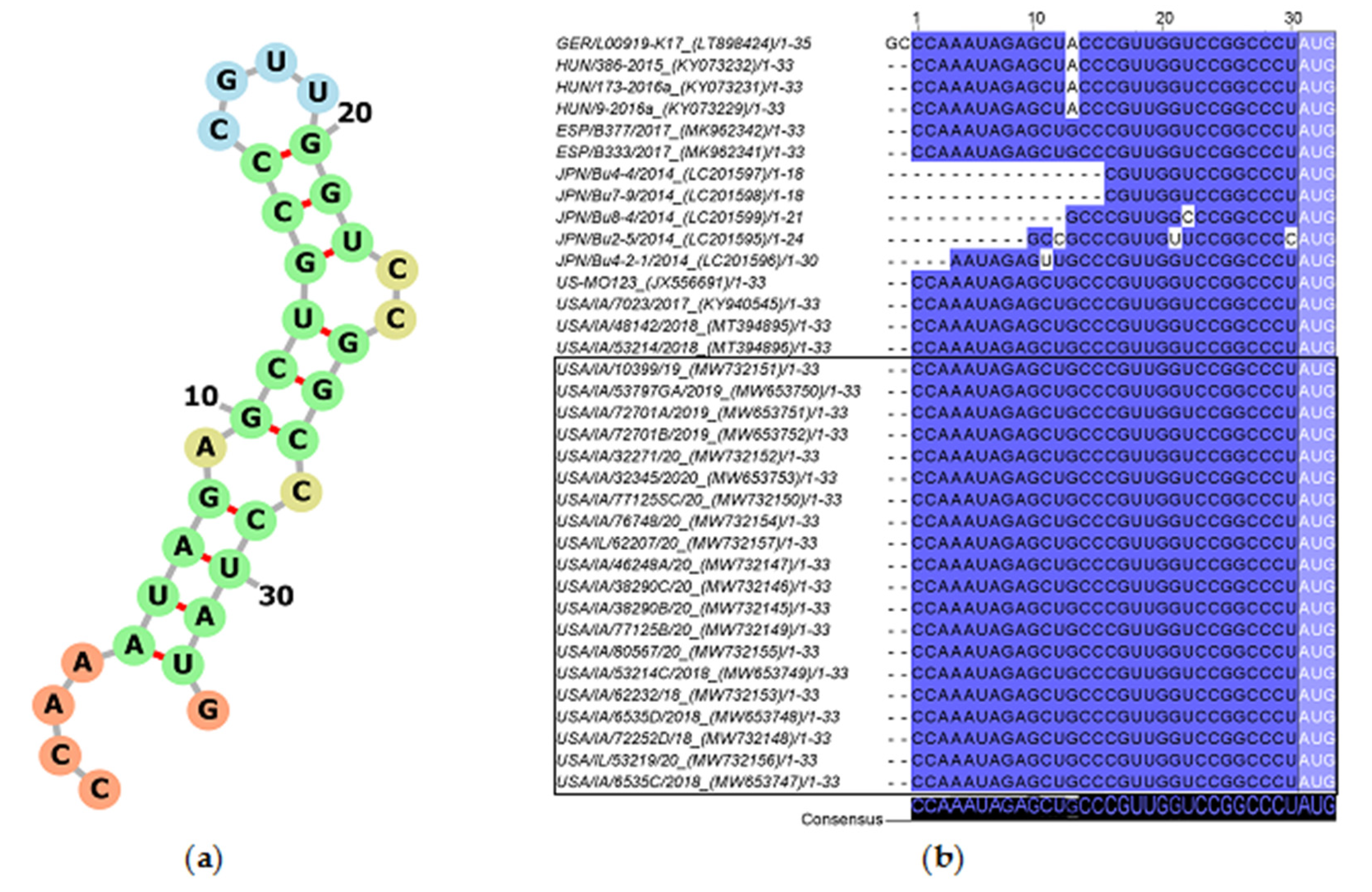
3.4.1. 5′UTR Analysis
3.4.2. Frameshift Analysis
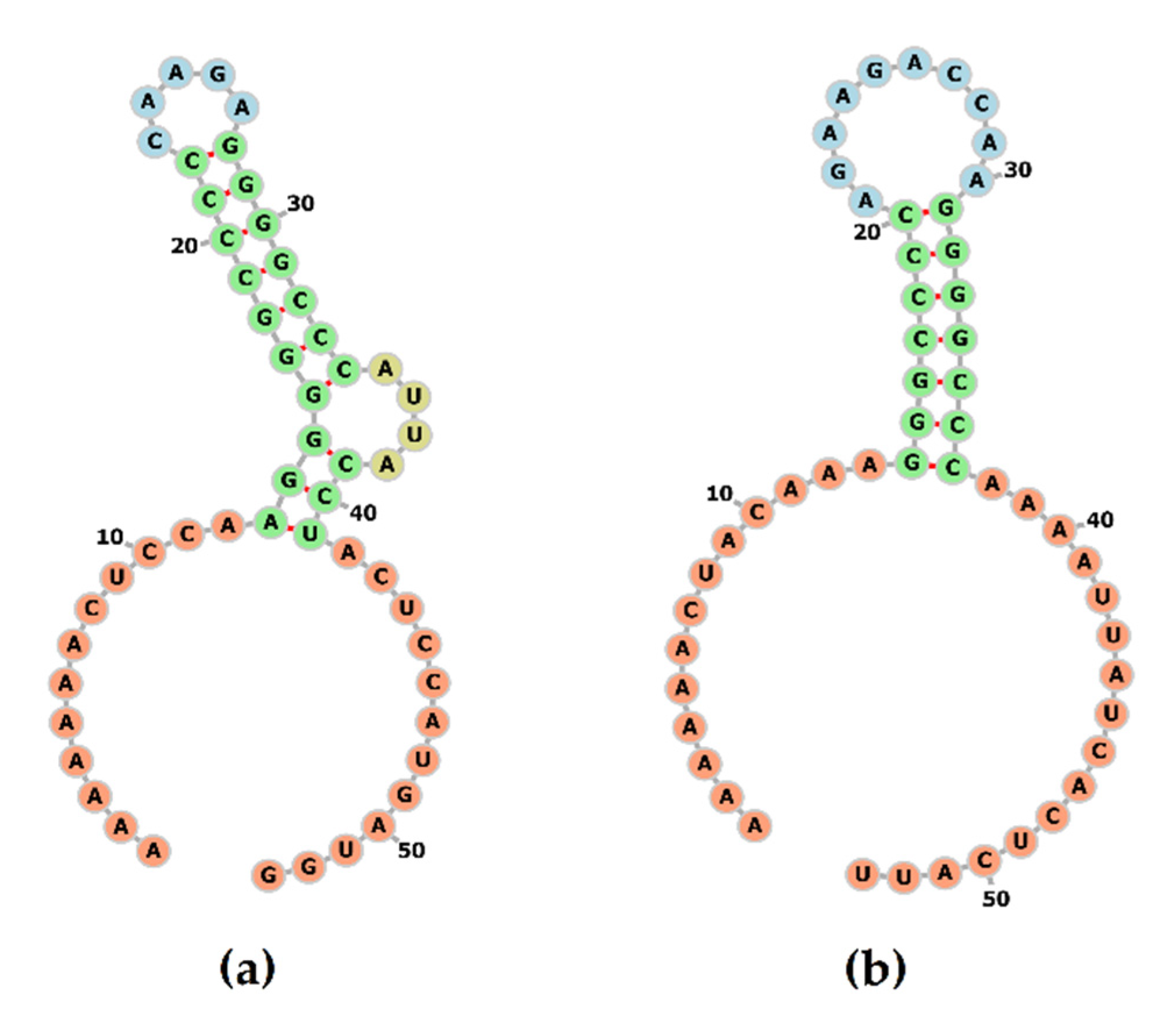
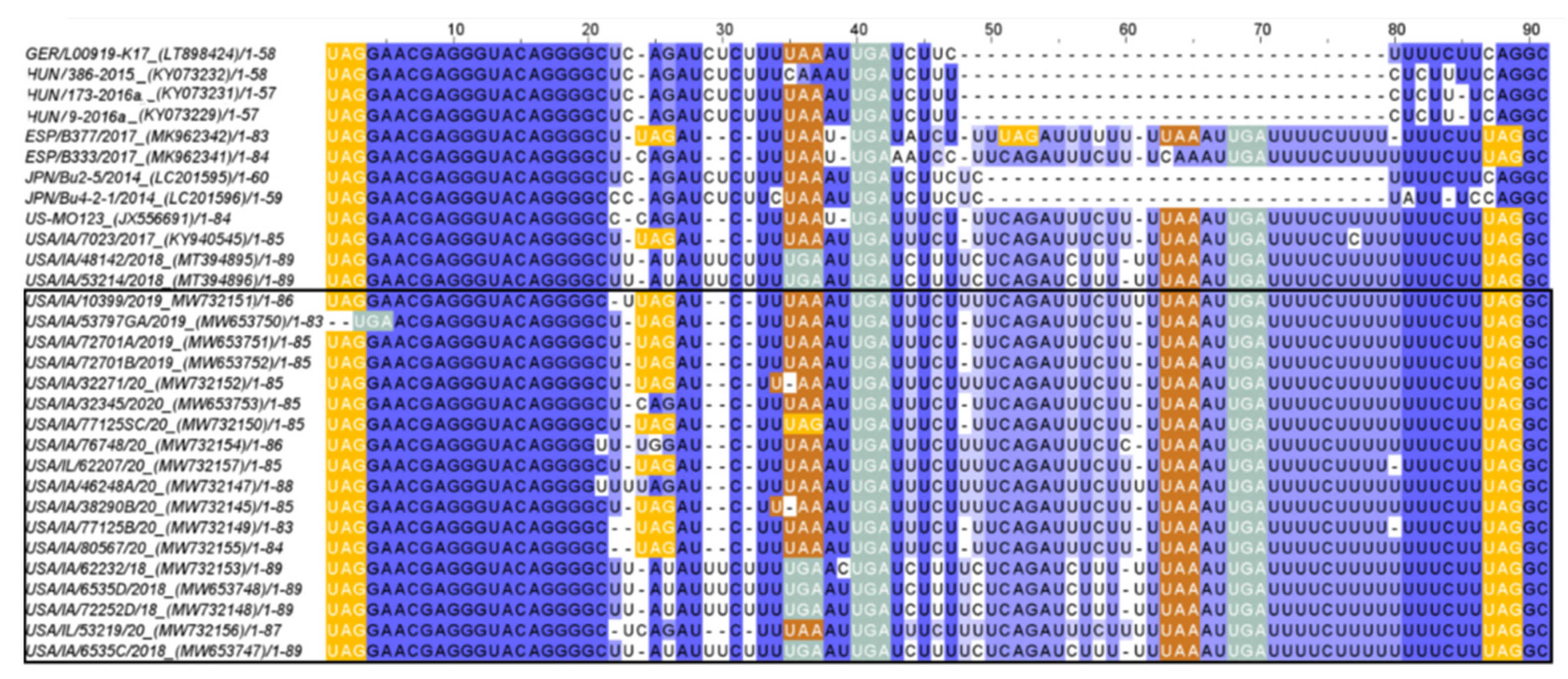
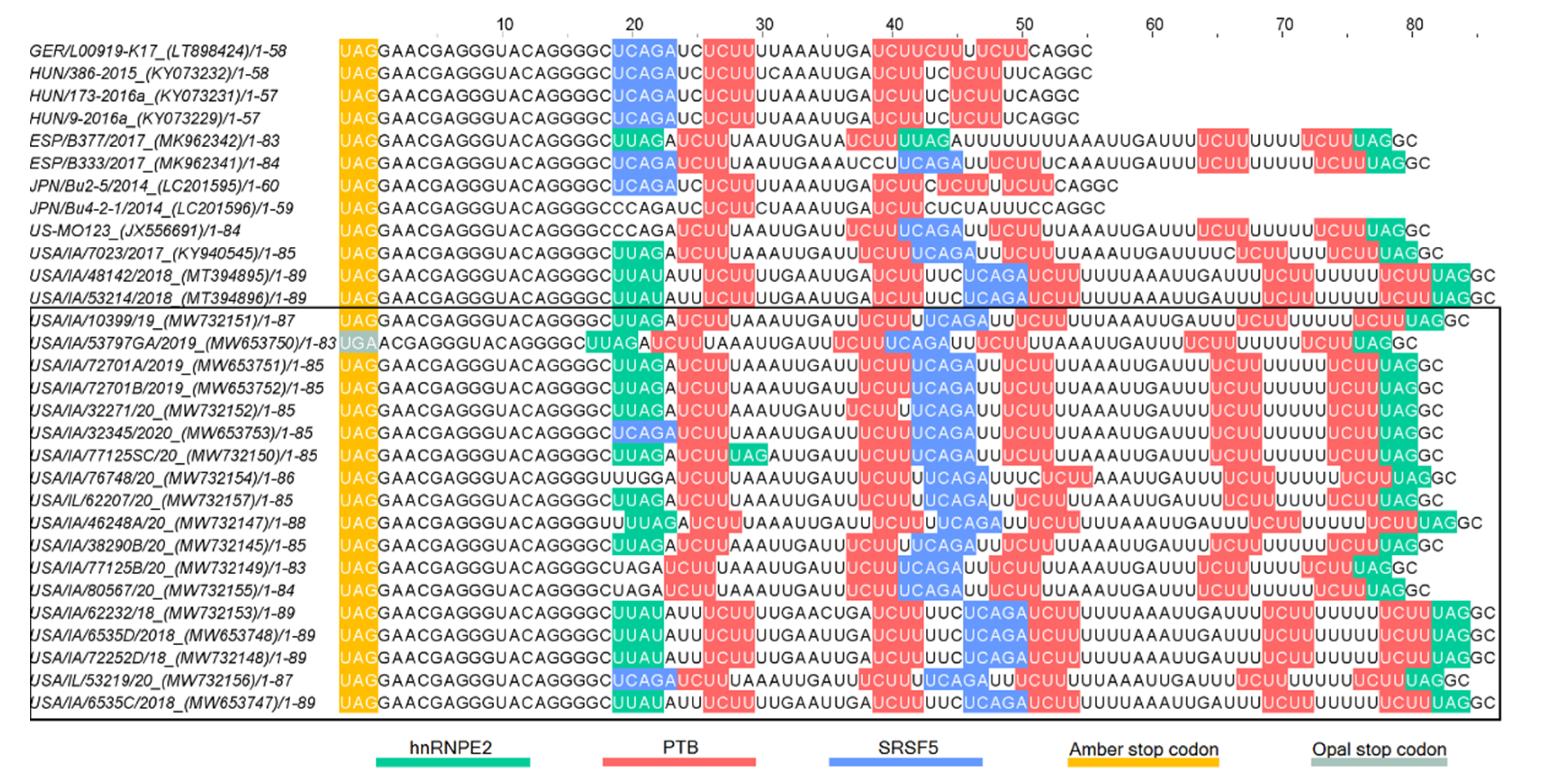
3.4.3. 3′UTR Analysis
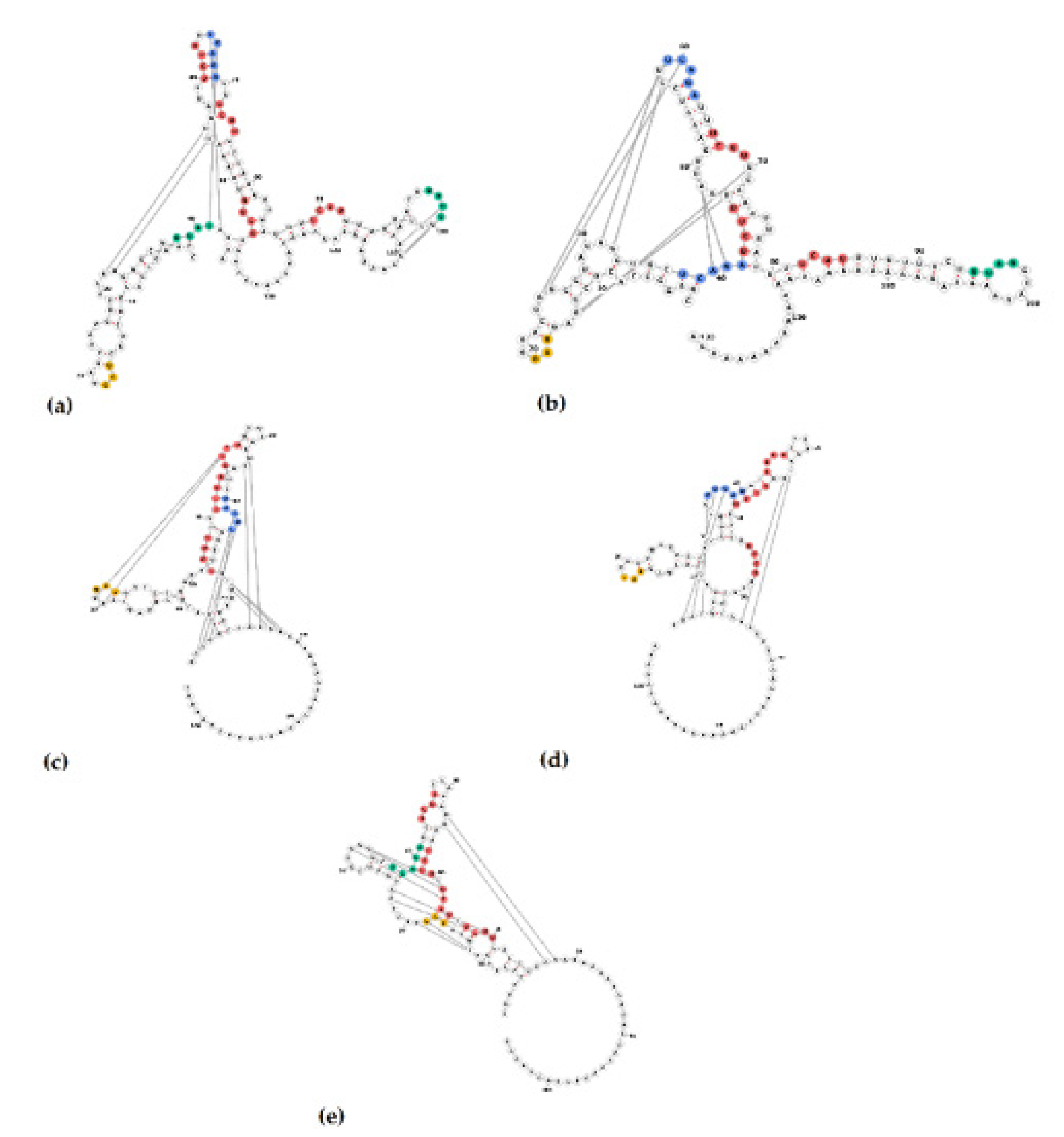
3.5. Identification of Putative VPg and RNA-Protein Binding Regions
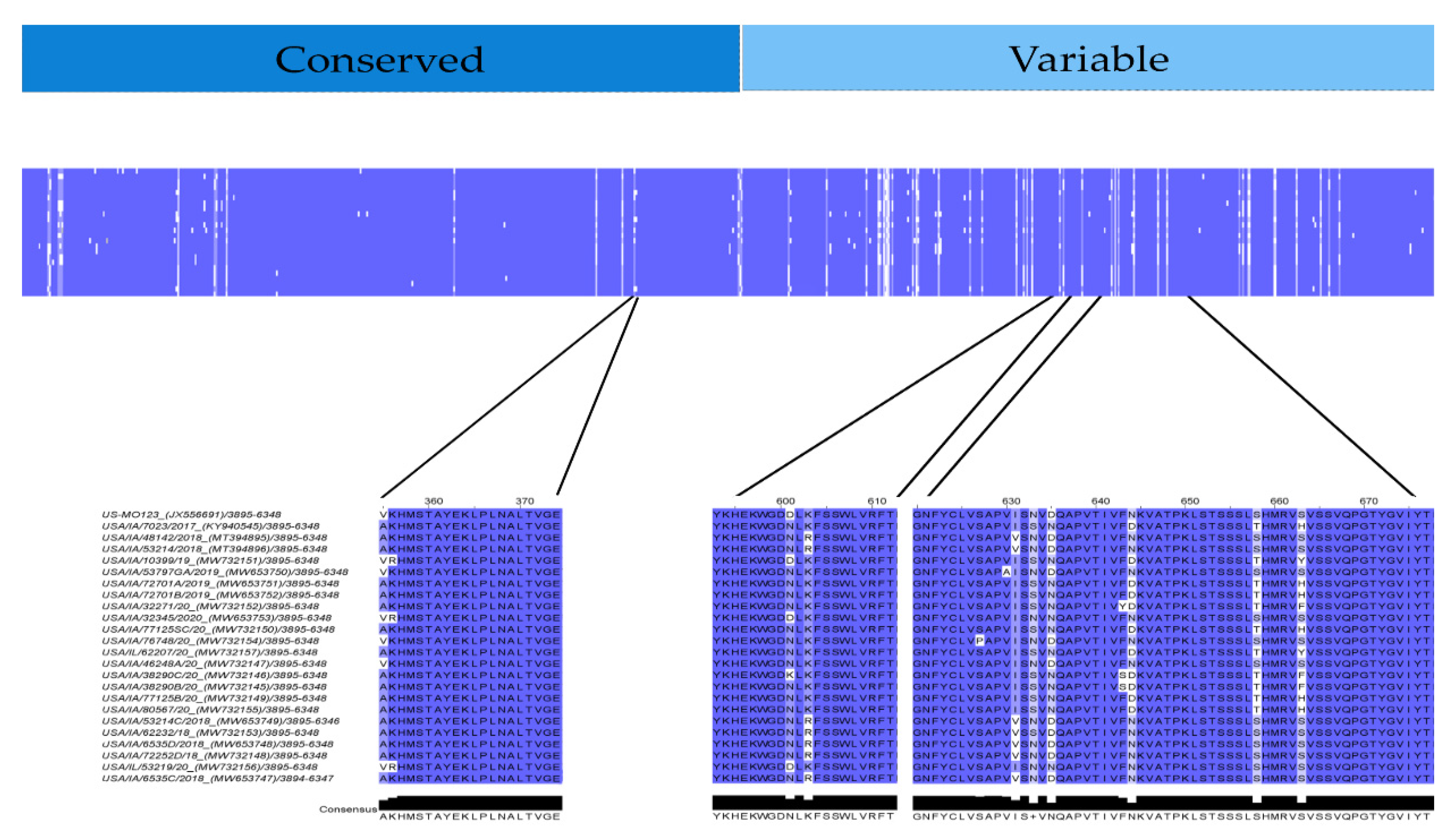
3.6. Linear Antigen Epitope Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrew, M.Q.K.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. (Eds.) Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Inc.: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Kurtz, J.B.; Lee, T.W.; Pickering, D. Astrovirus associated gastroenteritis in a children’s ward. J. Clin. Pathol. 1977, 30, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D.; et al. Astrovirus encephalitis in boy with x-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Englund, L.; Chriel, M.; Dietz, H.H.; Hedlund, K.O. Astrovirus epidemiologically linked to pre-weaning diarrhoea in mink. Vet. Microbiol. 2002, 85, 1–11. [Google Scholar] [CrossRef]
- de Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals—Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, N.; Honce, R.; Schultz-Cherry, S. Astrovirus evolution and emergence. Infect. Genet. Evol. 2019, 69, 30–37. [Google Scholar] [CrossRef]
- Blomström, A.L.; Widén, F.; Hammer, A.S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E.; et al. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385–1392. [Google Scholar] [CrossRef]
- Boujon, C.L.; Koch, M.C.; Wuthrich, D.; Werder, S.; Jakupovic, D.; Bruggmann, R.; Seuberlich, T. Indication of cross-species transmission of astrovirus associated with encephalitis in sheep and cattle. Emerg. Infect. Dis. 2017, 23, 1604–1608. [Google Scholar] [CrossRef]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Hoper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Küchler, L.; Rüfli, I.; Koch, M.C.; Hierweger, M.M.; Kauer, R.V.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Zanolari, P.; Seuberlich, T.; et al. Astrovirus-associated polioencephalomyelitis in an alpaca. Viruses 2020, 13, 50. [Google Scholar] [CrossRef]
- Arruda, B.; Arruda, P.; Hensch, M.; Chen, Q.; Zheng, Y.; Yang, C.; Honorato Gatto, I.; Matias Ferreyra, F.; Gauger, P.; Schwartz, K.; et al. Porcine astrovirus type 3 in central nervous system of swine with polioencephalomyelitis. Emerg. Infect. Dis. J. 2018, 23, 2097. [Google Scholar] [CrossRef]
- Boujon, C.L.; Koch, M.C.; Kauer, R.V.; Keller-Gautschi, E.; Hierweger, M.M.; Hoby, S.; Seuberlich, T. Novel encephalomyelitis-associated astrovirus in a muskox (ovibos moschatus): A surprise from the archives. Acta Vet. Scand. 2019, 61, 31. [Google Scholar] [CrossRef]
- Vu, D.L.; Bosch, A.; Pintó, R.M.; Ribes, E.; Guix, S. Human astrovirus mlb replication in vitro: Persistence in extraintestinal cell lines. J. Virol. 2019, 93, e00557-19. [Google Scholar] [CrossRef]
- Janowski, A.B.; Bauer, I.K.; Holtz, L.R.; Wang, D. Propagation of astrovirus va1, a neurotropic human astrovirus, in cell culture. J. Virol. 2017, 91, e00740-17. [Google Scholar] [CrossRef]
- Janowski, A.B.; Klein, R.S.; Wang, D.; Griffin, D.E. Differential in vitro infection of neural cells by astroviruses. Mbio 2019, 10, e01455-19. [Google Scholar] [CrossRef] [PubMed]
- Lulla, V.; Firth, A.E. A hidden gene in astroviruses encodes a viroporin. Nat. Commun. 2020, 11, 4070. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and pathological phenotype of bovine astrovirus ch13/neuros1 infection in neurologically-diseased cattle: Towards assessment of causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Boujon, C.L.; Koch, M.C.; Seuberlich, T. The expanding field of mammalian astroviruses: Opportunities and challenges in clinical virology. In Advances in Virus Research; Beer, M., Höper, D., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 99, pp. 109–137. [Google Scholar]
- Monceyron, C.; Grinde, B.; Jonassen, T.O. Molecular characterisation of the 3′-end of the astrovirus genome. Arch Virol. 1997, 142, 699–706. [Google Scholar] [CrossRef]
- Xiao, C.-T.; Halbur, P.G.; Opriessnig, T. Complete genome sequence of a newly identified porcine astrovirus genotype 3 strain us-mo123. J. Virol. 2012, 86, 13126. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Xiao, C.T.; Halbur, P.G. Porcine astrovirus type 5-associated enteritis in pigs. J. Comp. Pathol. 2020, 181, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.C. Detection by electron microscopy of caliciviruses, astroviruses and rotavirus-like particles in the faeces of piglets with diarrhoea. Vet. Rec. 1980, 107, 532–533. [Google Scholar] [PubMed]
- Indik, S.; Valicek, L.; Smid, B.; Dvorakova, H.; Rodak, L. Isolation and partial characterization of a novel porcine astrovirus. Vet. Microbiol. 2006, 117, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, A.; Hause, B.M. Detection and characterization of a novel genotype of porcine astrovirus 4 from nasal swabs from pigs with acute respiratory disease. Arch Virol. 2016, 161, 2575–2579. [Google Scholar] [CrossRef]
- Guix, S.; Bosch, A.; Pintó, R.M. Astrovirus taxonomy. In Astrovirus Research: Essential Ideas, Everyday Impacts, Future Directions; Schultz-Cherry, S., Ed.; Springer: New York, NY, USA, 2013; pp. 97–118. [Google Scholar]
- Boros, Á.; Albert, M.; Pankovics, P.; Bíró, H.; Pesavento, P.A.; Phan, T.G.; Delwart, E.; Reuter, G. Outbreaks of neuroinvasive astrovirus associated with encephalomyelitis, weakness, and paralysis among weaned pigs, hungary. Emerg. Infect. Dis. J. 2018, 23, 1982. [Google Scholar] [CrossRef]
- Matias Ferreyra, F.S.; Bradner, L.K.; Burrough, E.R.; Cooper, V.L.; Derscheid, R.J.; Gauger, P.C.; Harmon, K.M.; Madson, D.; Pineyro, P.E.; Schwartz, K.J.; et al. Polioencephalomyelitis in domestic swine associated with porcine astrovirus type 3. Vet. Pathol. 2020, 57, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Swine Disease Reporting System. Iowa State University. Available online: https://fieldepi.research.cvm.iastate.edu/SDRS/ (accessed on 12 July 2021).
- Anonymous. Morrison Swine Health Monitoring Program; University of Minnesota: Minnesota, MN, USA, 2021. [Google Scholar]
- Matias Ferreyra, F.; Arruda, E.H.P.; Bradner, K.L.; Harmon, M.K.; Zhang, M.; Gimenez-Lirola, L.; Arruda, L.B. Experimental porcine astrovirus type 3-associated polioencephalomyelitis in swine. Vet. Pathol. 2021, in press. [Google Scholar]
- Reuter, G.; Pankovics, P.; Boros, Á. Nonsuppurative (aseptic) meningoencephalomyelitis associated with neurovirulent astrovirus infections in humans and animals. Clin. Microbiol. Rev. 2018, 31, e00040-18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Monroe, S.S.; Koonin, E.V.; Stine, S.E.; Glass, R.I. Rna sequence of astrovirus: Distinctive genomic organization and a putative retrovirus-like ribosomal frameshifting signal that directs the viral replicase synthesis. Proc. Natl. Acad. Sci. USA 1993, 90, 10539–10543. [Google Scholar] [CrossRef] [PubMed]
- Jonassen, C.M.; Jonassen, T.O.; Grinde, B. A common rna motif in the 3′end of the genomes of astroviruses, avian infectious bronchitis virus and an equine rhinovirus. J. Gen. Virol. 1998, 79, 715–718. [Google Scholar] [CrossRef]
- Tengs, T.; Kristoffersen, A.B.; Bachvaroff, T.R.; Jonassen, C.M. A mobile genetic element with unknown function found in distantly related viruses. J. Virol. 2013, 10, 132. [Google Scholar] [CrossRef]
- de Nova-Ocampo, M.; Soliman, M.C.; Espinosa-Hernandez, W.; Velez-Del Valle, C.; Salas-Benito, J.; Valdes-Flores, J.; Garcia-Morales, L. Human astroviruses: In silico analysis of the untranslated region and putative binding sites of cellular proteins. Mol. Biol. Rep. 2019, 46, 1413–1424. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. Fruits of virus discovery: New pathogens and new experimental models. J. Virol. 2015, 89, 1486–1488. [Google Scholar] [CrossRef][Green Version]
- Firth, A.E.; Atkins, J.F. Candidates in astroviruses, seadornaviruses, cytorhabdoviruses and coronaviruses for +1 frame overlapping genes accessed by leaky scanning. J. Virol. 2010, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Brierley, I.; Vidakovic, M. V, 2.Ribosomal frameshifting in astroviruses. Perspect. Med. Virol. 2003, 9, 587–606. [Google Scholar] [PubMed]
- Moss, W.N.; Steitz, J.A. In silico discovery and modeling of non-coding rna structure in viruses. Methods 2015, 91, 48–56. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using clustal omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (itol): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (itol) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Tassoni, L.; Zamperin, G.; Schiavon, E.; Vendramin, V.; Cavicchio, L.; Mion, M.; Tonon, F.T.; Monne, I.; Beato, M.S. First whole genome characterization of porcine astrovirus detected in swine faeces in italy. Vet. Ital. 2019, 55, 221–229. [Google Scholar]
- Amimo, J.O.; Machuka, E.M.; Abworo, E.O.; Vlasova, A.N.; Pelle, R. Whole genome sequence analysis of porcine astroviruses reveals novel genetically diverse strains circulating in east african smallholder pig farms. Viruses 2020, 12, 1262. [Google Scholar] [CrossRef]
- Waterhouse, A.; Procter, J.; Martin, D.; Clamp, M.; Barton, G. Jalview version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Lorenz, A.R.; Bernhart, S.H.; Neubock, R.; Hofacker, I.L. The vienna rna websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar]
- Lorenz, R.; Bernhart, S.H.; Zu Siederdissen, C.H.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. Viennarna package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Kato, Y.; Hamada, M.; Akutsu, T.; Asai, K. Ipknot: Fast and accurate prediction of rna secondary structures with pseudoknots using integer programming. Bioinformatics 2011, 27, i85–i93. [Google Scholar] [CrossRef] [PubMed]
- Kerpedjiev, P.; Hammer, S.; Hofacker, I.L. Forna (force-directed RNA): Simple and effective online rna secondary structure diagrams. Bioinformatics 2015, 31, 3377–3379. [Google Scholar] [CrossRef]
- Al-Mutairy, B.; Walter, J.E.; Pothen, A.; Mitchell, D.K. Genome prediction of putative genome-linked viral protein (vpg) of astroviruses. Virus Genes 2005, 31, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Mittelholzer, C.; Hedlund, K.O.; Englund, L.; Dietz, H.H.; Svensson, L. Molecular characterization of a novel astrovirus associated with disease in mink. J. Gen. Virol. 2003, 84, 3087–3094. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the expasy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Peng, Z.; Kurgan, L. High-throughput prediction of rna, dna and protein binding regions mediated by intrinsic disorder. Nucleic Acids Res. 2015, 43, e121. [Google Scholar] [CrossRef]
- Yao, B.; Zhang, L.; Liang, S.; Zhang, C. Svmtrip: A method to predict antigenic epitopes using support vector machine to integrate tri-peptide similarity and propensity. PLoS ONE 2012, 7, e45152. [Google Scholar] [CrossRef] [PubMed]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. Vaxijen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef]
- Rawal, G.; Matias Ferreyra, F.S.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Mueller, A.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Detection and cellular tropism of porcine astrovirus type 3 on breeding farms. Viruses 2019, 11, 1051. [Google Scholar] [CrossRef]
- Rawal, G.; Ferreyra, F.M.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Allison, G.; Linhares, D.C.L.; Arruda, B.L. Ecology of porcine astrovirus type 3 in a herd with associated neurologic disease. Viruses 2020, 12, 992. [Google Scholar] [CrossRef]
- Xiao, C.T.; Gimenez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (pastvs) with high prevalence and frequent co-infection of individual pigs with multiple pastv types. J. Gen. Virol. 2013, 94, 570–582. [Google Scholar] [CrossRef]
- Fuentes, C.; Bosch, A.; Pintó, R.M.; Guix, S. Identification of human astrovirus genome-linked protein (vpg) essential for virus infectivity. J. Virol. 2012, 86, 10070–10078. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, Z.A.; Kieft, J.S. Viral rna structure-based strategies to manipulate translation. Nat. Rev. Microbiol. 2019, 17, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Brierley, I. Non-canonical translation in rna viruses. J. Gen. Virol. 2012, 93, 1385–1409. [Google Scholar] [CrossRef] [PubMed]
- Willcocks, M.M.; Carter, M.J. The 3′terminal sequence of a human astrovirus. Arch Virol. 1992, 124, 279–289. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | Sequence Target | Region Target | |
|---|---|---|---|
| POAstV3 US-MO123—JX556691 (5′-3′) | PoAstV3 USA/IA/7023/2017—KY940545 (5′-3′) | ||
| F1_T7 | ATGGTACCTAATACGACTCACTATAGCCAAATAGAGCTGCCCGTTGGTCC | ORF1 | |
| R1008 | CAGAGTAGCGTACATCGGGCATG | CGGAGTAACGCACATCGGGCATG | ORF1a |
| F392 | TCTCCCAAGAGCTCTCCCAA | NA | ORF1a |
| R1345 | CCTGTTCCTGAGCCATCCTG | NA | ORF1a |
| F485 | GCCGGTTAGGGTTCATCACA | ORF1a | |
| R1591 | GCAACTGACAGAGCACCTGA | ORF1a | |
| F845 | CATATTCAAAGGCCCAAGTTCTGGCCCTTG | CATACTCAAAGCCCAGGTTCTAGCTCTTG | ORF1a |
| R1662 | CAGGTGAACCACTCATACCATTCCGA | CAGGTGAGCCACTCATACCATTCCGG | ORF1a |
| F1465 | GTCCATCGTGTGCCGGATAAGGACATAG | GTCCACCGTGTGCCAGATAAGGACATAG | ORF1a |
| R2217 | GTGTCTTGAGGCTCTTCCTTAACAGGC | GTGTCCTGAGGCTCTTCCTTAACAGGC | ORF1a |
| F2001 | CCGTGCTGTCTGGTCAGAAGAAGAGTAC | CCGTGCTGTCTGGTCAGAAGAGGAATAC | ORF1a |
| R3203 | CCACATTGGCCAGAGCGTGTC | ORF1a | |
| F2462 | GGGAGAAAGGCTTGGTTCCT | ORF1a | |
| R3920 | CAGGGAGGTTATGGCCAAGG | ORF2 | |
| F3127 | GTTCTGACCCTATATTTGCACGTATAGGATGTCAC | GTTCTGACCCTATATTTGCACGTATAGGTTGTCAC | ORF1a |
| R4855 | GTCACACGATCTAGTGTTCCAAGGGC | GTCACACGATCCAGAGTTCCAAGGGC | ORF2 |
| F4308 | CTTCTCAGTGCGAGTCTCGG | ORF2 | |
| R5326 | AAAGTGGTGGCCGTAGATGG | ORF2 | |
| F4672 | CCGCGACAGACGTGGTGG | ORF2 | |
| R5924 | CGTCAGTCACATTATAGTTGCCAAGGG | CGTCAGCCACATTGTAGTTACCAAGGG | ORF2 |
| F5486 | GAGCTGGTAAAGGCATGGGT | NA | ORF2 |
| R6283 | TGGATAAGAGCGTCGGCATC | ORF2 | |
| F6108 | CCGTCCGACCATGATTGACA | ORF2 | |
| R6352 | CGAGGGTACAGGGGCTTAGA | TCTAAGCCCCTGTACCCTCG | 3′ UTR |
| F5598 | ACCTGGTATCGGCTGGACT | ORF2 | |
| R6405_M13 | TGTAAAACGACGGCCAGTGCCTAAAGAAAAAAAAGAAAATCAATTTAA | 3′ UTR | |
| F2804 | CCCTAATATAGCCGCCACGT | NA | ORF1a |
| R3987 | GTGTCCTGATGTGCCTAGCA | NA | ORF2 |
| F946 | ACCCACAGTGAAAGGACAGC | NA | ORF1 |
| F5623 | CCTGGTATCGGCTGGACTGTT | NA | ORF2 |
| Strain Origin | ORF1a | nsp1a | ORF1ab | nsp1ab | ORF2 | Capsid Protein |
|---|---|---|---|---|---|---|
| USA | 89.7–100% | 94.7–100% | 91.9–100% | 96.3–100% | 90.7–100% | 93.5–100% |
| Japan | 89.1–100% | 95.2–99.9% | 89.4–99.9% | 95.3–99.9% | 65.1–99.9% | 65.6–99.9% |
| Spain | 92.9% | 96.3% | 93.2% | 96.6% | 92.7% | 95.6% |
| Hungary | 99.6–99.7% | 99.4–99.6% | 99.6–99.6% | 99.6–99.7% | 99.1–99.4% | 98.8–99.2% |
| Prediction Tool: SVMTriP Online Prediction Tool | |||||
|---|---|---|---|---|---|
| Recommended epitopes | Score | ORF 2 region | ORF 2 position | VaxiJen v2.0 result (score) | |
| USA | AKHMSTAYEKLPLNALTVGE | 1 | Conserved | 355–374 | Probable antigen (0.5478) |
| YKHEKWGDNLKFSSWLVRFT | 0.92 | Variable | 593–612 | Probable antigen (0.8778) | |
| Hungary | AKHMSTAYEKLPLNALTVGE | 1 | Conserved | 292–311 | Probable antigen (0.5478) |
| TNCADVLAYKHEKWGDNLKF | 0.953 | Variable | 522–541 | Probable antigen (0.8269) | |
| LAKEVAKEVVKEEKRNQARS | 0.893 | Conserved | 15–34 | Probable non-antigen (0.1453) | |
| Japan | IMTMTIPQRSNLARHMSTAY | 1 | Conserved | 347–368 | Probable non-antigen (0.0369) |
| LAKEVAKEVVKEEKKVQARR | 0.964 | Conserved | 77–96 | Probable non-antigen (0.0094) | |
| TNCADVLAYKHEKWGDNLKF | 0.892 | Variable | 591–610 | Probable antigen (0.8269) | |
| Spain | AKHMSTAYEKLPLNALTVGE | 1 | Conserved | 355–374 | Probable antigen (0.5478) |
| TNCADVLAYKHEKWGDDLKF | 0.879 | Variable | 585–604 | Probable antigen (0.8161) | |
| ANQATSHSPVINNVFPVLGQ | 0.862 | Variable | 496–515 | Probable non-antigen (0.3368) | |
| Germany | LAKEVAKEVVKEEKRFQARS | 1 | Conserved | 77–96 | Probable non-antigen (0.2543) |
| PVFLGLNLGASSSTVDNIFL | 0.999 | Variable | 544–563 | Probable antigen (0.9103) | |
| AKHMSTAYEKLPLNALTVGE | 0.911 | Conserved | 354–373 | Probable antigen (0.5478) | |
| TNCADVLAYKHEKWGDNLKF | 0.869 | Variable | 584–603 | Probable antigen (0.8269) | |
| Prediction tool: Immunomedicine predicted antigenic peptides web-based tool | |||||
| Recommended epitopes | Score | ORF 2 region | ORF 2 position | VaxiJen v2.0 result (score) | |
| USA | WVGDLFIVEVVG | >1.2 | Conserved | 298–309 | Probable non-antigen (0.0845) |
| GNFYCLVSAPVISNVNQAPVTIVFNKVATPKLSTSSSLSHMRVSVSSVQPGTYGVIYT | >1.2 | Variable | 620–677 | Probable antigen (0.5617) | |
| Hungary | WVGDLFIVEVVG | >1.2 | Conserved | 239–250 | Probable non-antigen (0.0845) |
| GNYYCLVSAPVISAVNQAPVTIVFDKVATPKLSTSSS | >1.2 | Variable | 561–597 | Probable non-antigen (0.3268) | |
| Japan | WVGDLFIVEVVG | >1.2 | Conserved | 308–319 | Probable non-antigen (0.0845) |
| GNFYCLVSAPIISNVDQAPVTIVANKVATPKLSTSSSLSHMRVSVSSVQPGTYGVIYT | >1.2 | Variable | 626–683 | Probable antigen (0.5449) | |
| Spain | GLWRLAGLKICCTPLVGPSAVTGSVYRLSLNL | >1.2 | Conserved | 195–226 | Probable non-antigen (0.1431) |
| WVGDLFIVEVVG | >1.2 | Conserved | 302–313 | Probable non-antigen (0.0845) | |
| GNFYCLVSAPVISNVDQAPVTIVFDKVATPKLSTSSSLSHMRVSVSSVQPGTYGVIYT | >1.2 | Variable | 624–681 | Probable antigen (0.5397) | |
| Germany | GNFYCLVSAPVVSDVNQAPVTIVSNKVATPKLSTSSSLSHMRVSVSSVQPGMYGVIYTLGS | >1.2 | Variable | 619–679 | Probable antigen (0.5507) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matias Ferreyra, F.; Harmon, K.; Bradner, L.; Burrough, E.; Derscheid, R.; Magstadt, D.R.; Michael, A.; de Almeida, M.N.; Schumacher, L.; Siepker, C.; et al. Comparative Analysis of Novel Strains of Porcine Astrovirus Type 3 in the USA. Viruses 2021, 13, 1859. https://doi.org/10.3390/v13091859
Matias Ferreyra F, Harmon K, Bradner L, Burrough E, Derscheid R, Magstadt DR, Michael A, de Almeida MN, Schumacher L, Siepker C, et al. Comparative Analysis of Novel Strains of Porcine Astrovirus Type 3 in the USA. Viruses. 2021; 13(9):1859. https://doi.org/10.3390/v13091859
Chicago/Turabian StyleMatias Ferreyra, Franco, Karen Harmon, Laura Bradner, Eric Burrough, Rachel Derscheid, Drew R. Magstadt, Alyona Michael, Marcelo Nunes de Almeida, Loni Schumacher, Chris Siepker, and et al. 2021. "Comparative Analysis of Novel Strains of Porcine Astrovirus Type 3 in the USA" Viruses 13, no. 9: 1859. https://doi.org/10.3390/v13091859
APA StyleMatias Ferreyra, F., Harmon, K., Bradner, L., Burrough, E., Derscheid, R., Magstadt, D. R., Michael, A., de Almeida, M. N., Schumacher, L., Siepker, C., Sitthicharoenchai, P., Stevenson, G., & Arruda, B. (2021). Comparative Analysis of Novel Strains of Porcine Astrovirus Type 3 in the USA. Viruses, 13(9), 1859. https://doi.org/10.3390/v13091859