
RNA Origami: Packaging a Segmented Genome in Orbivirus Assembly and Replication



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Overview of Orbivirus Replication
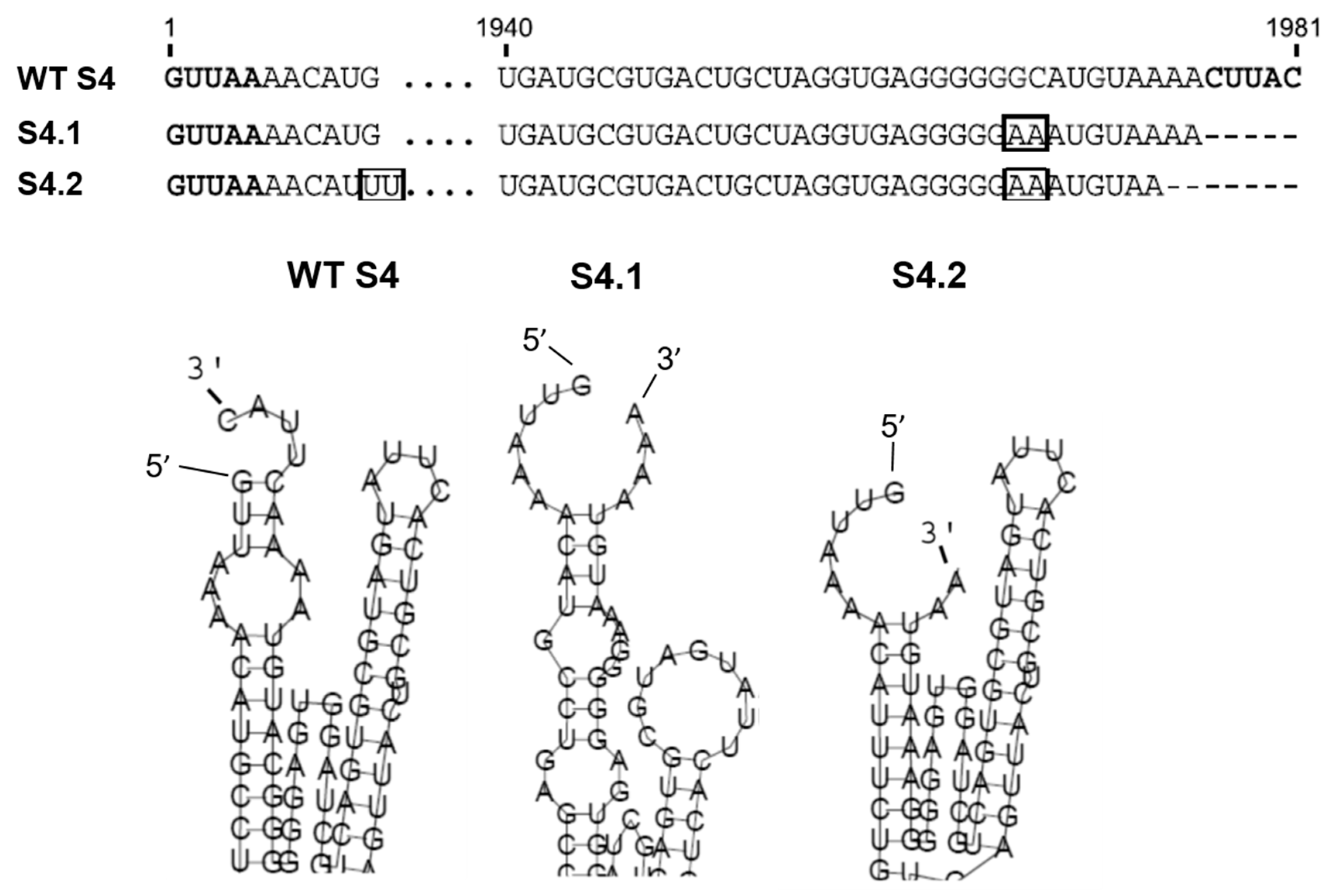
3. Structural Constraints in Genomic Segments Regulate BTV Genome Packaging
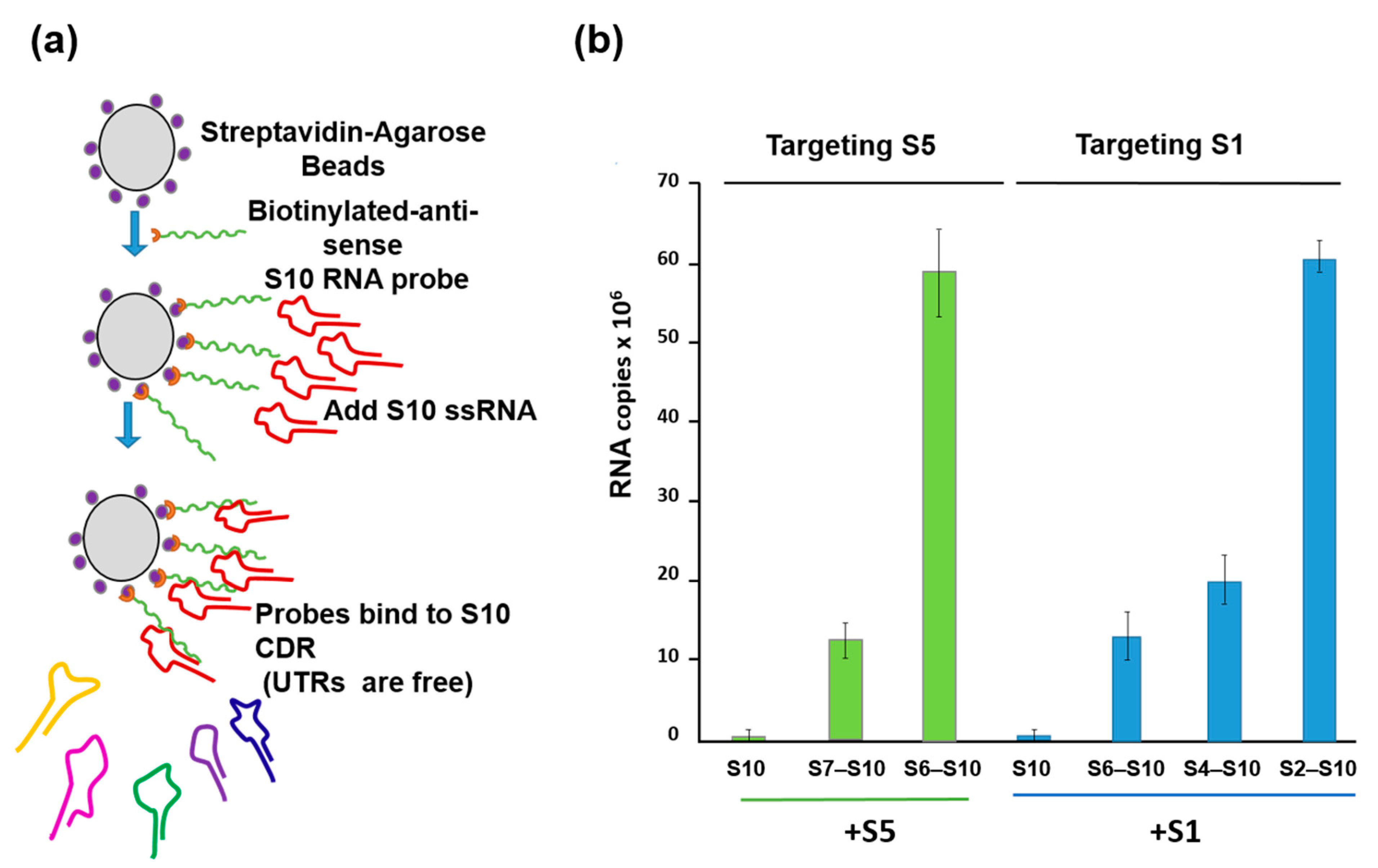
4. How Is a Complete Set of ssRNA Molecules Selected for Packaging?
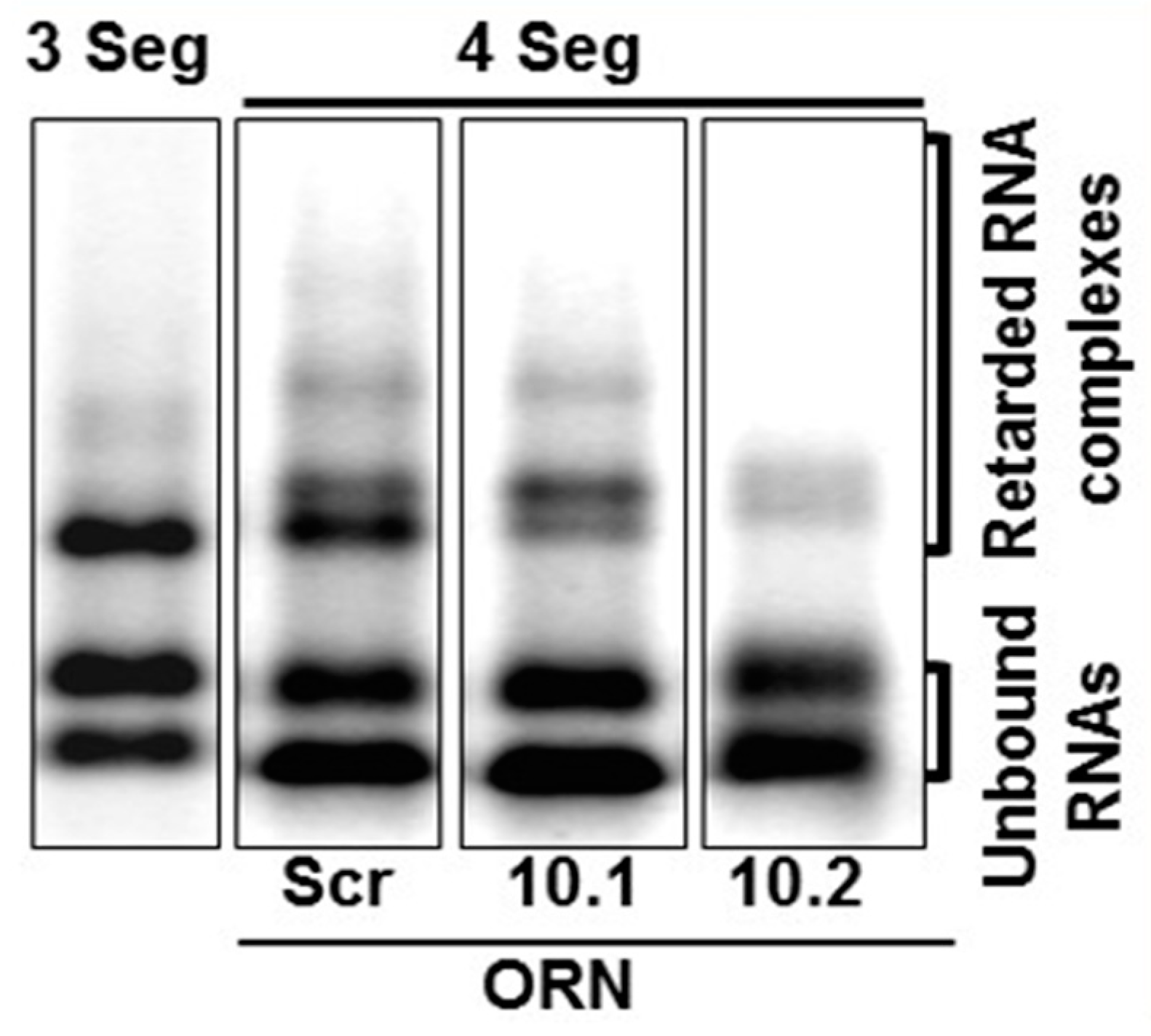
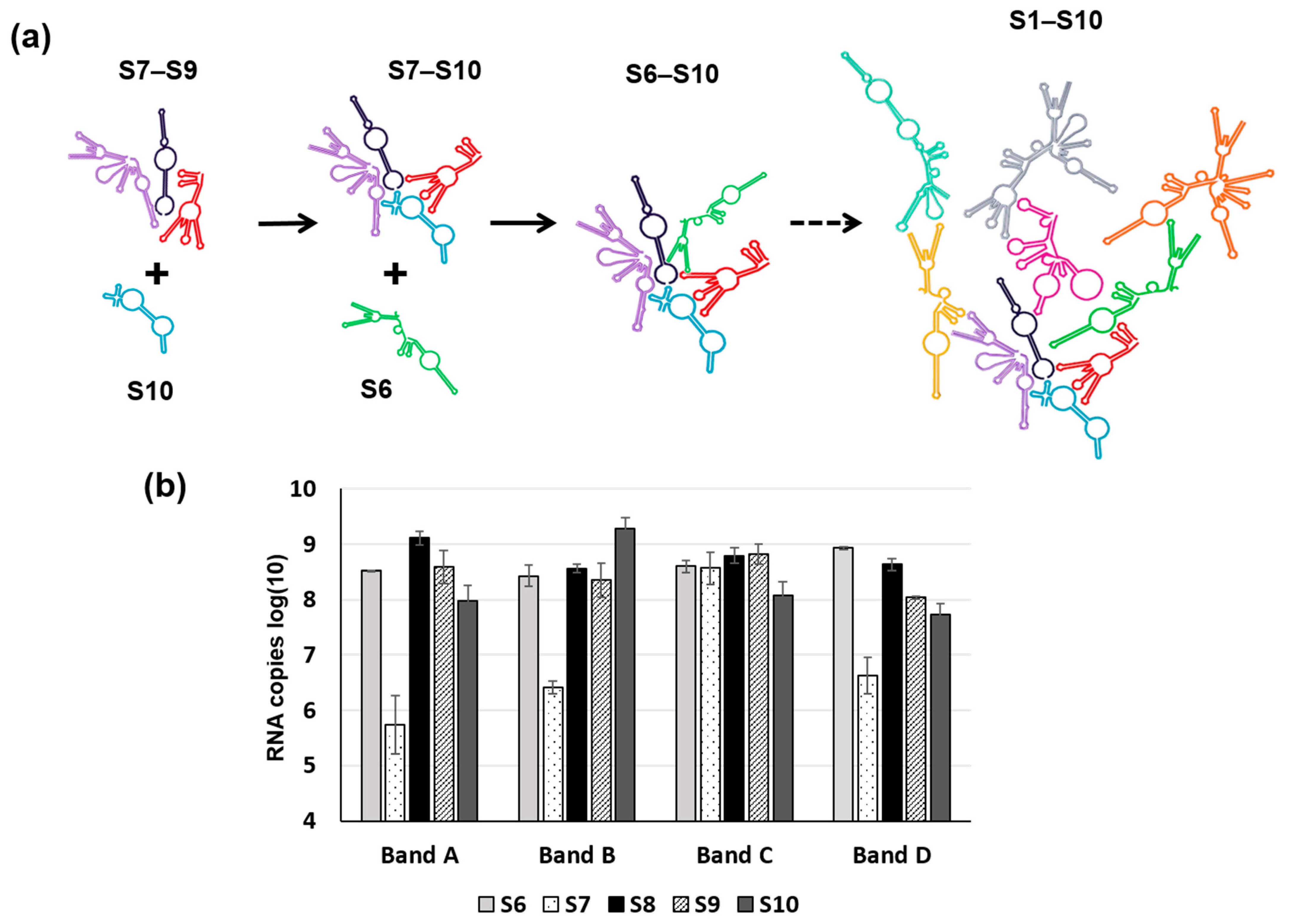
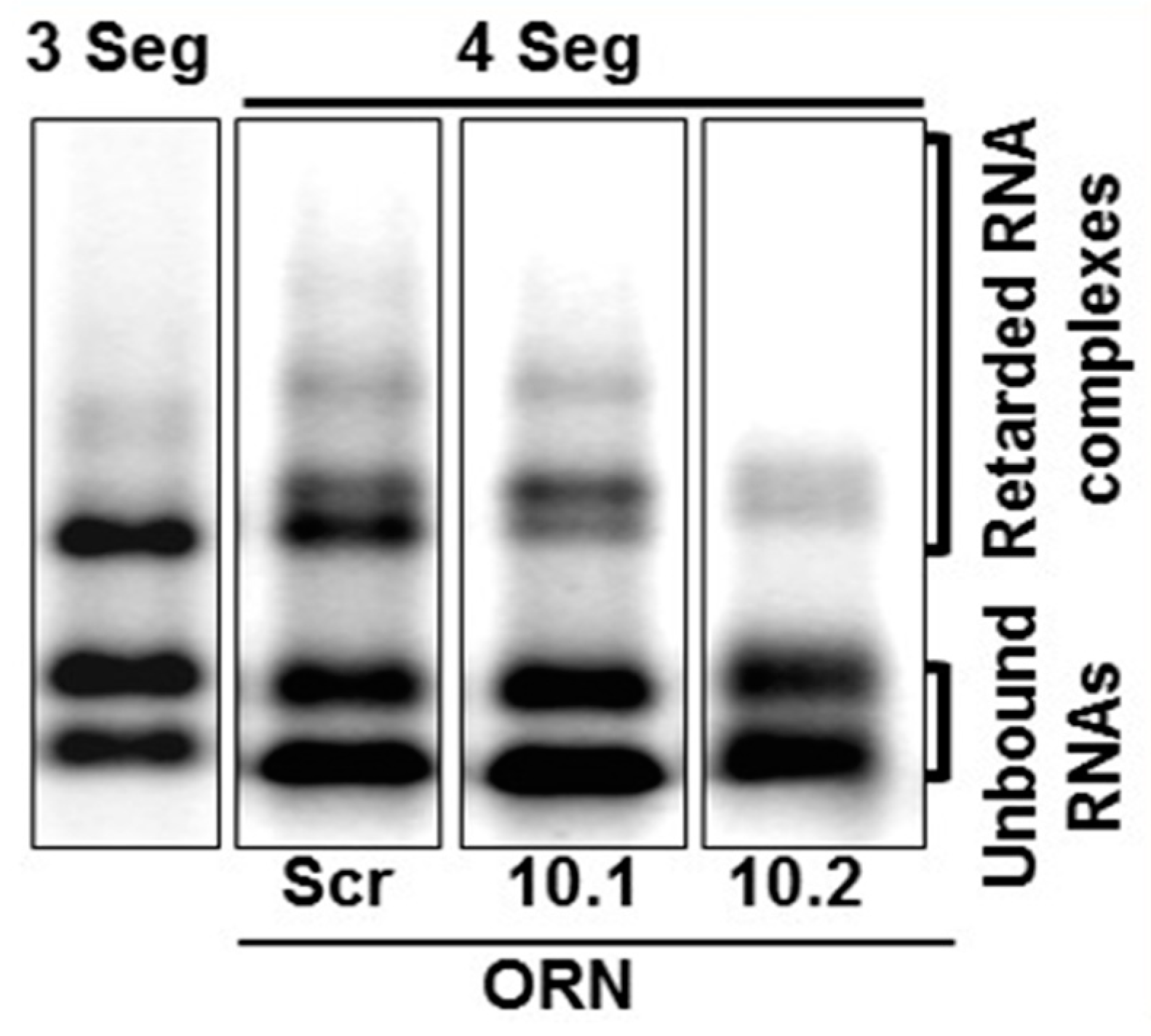
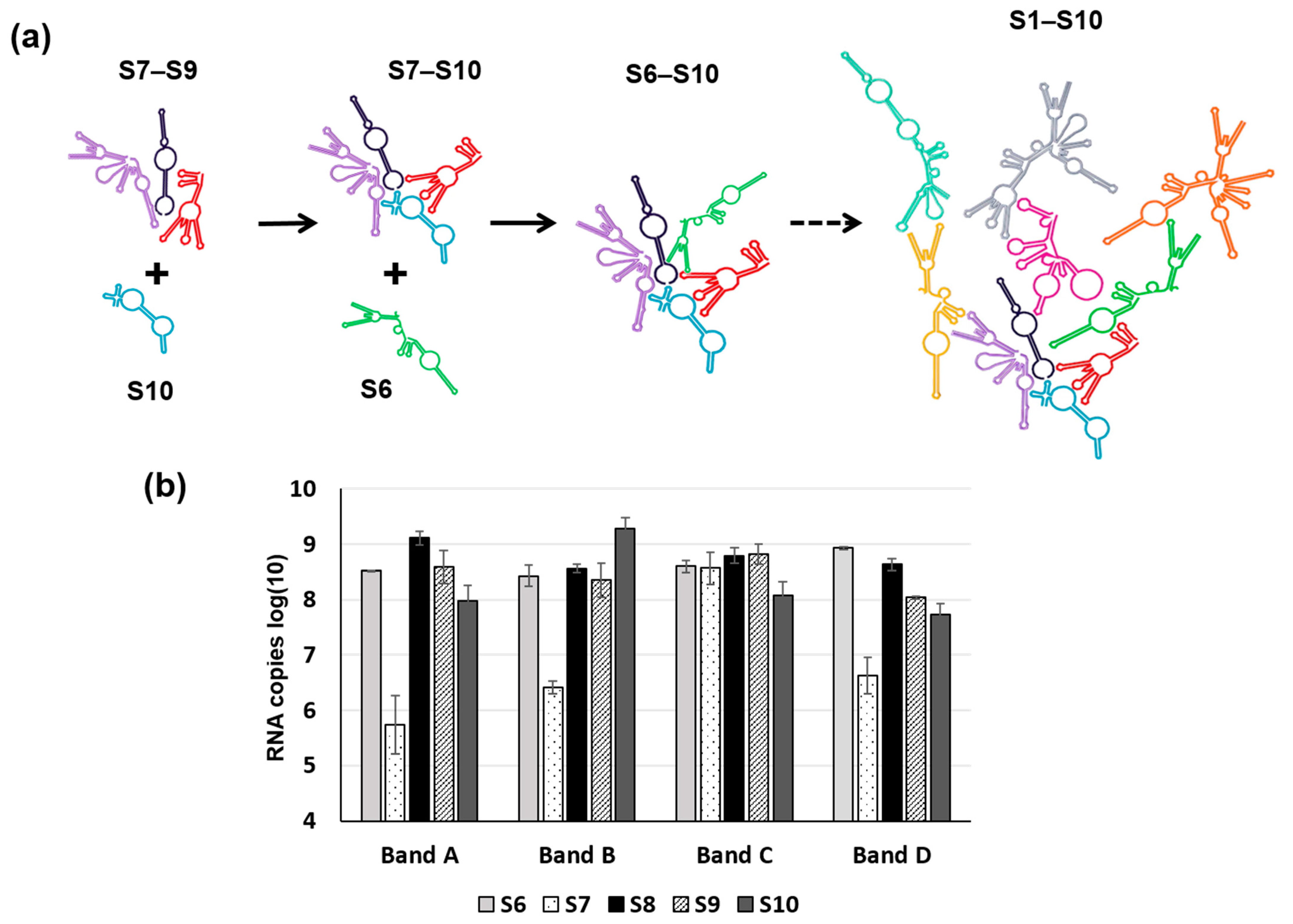
5. RNA Complex Formation and RNA Network May Initiate Packaging
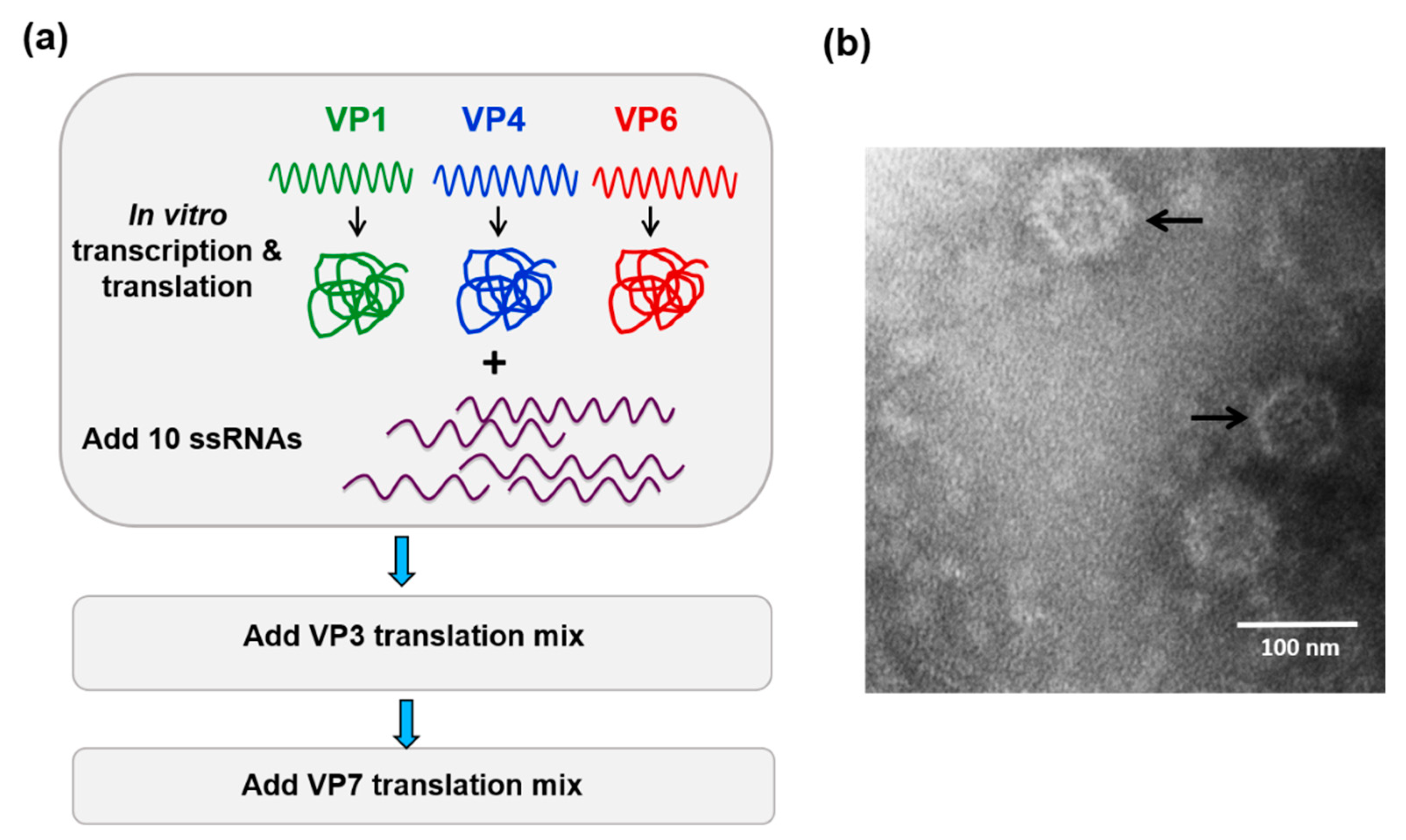
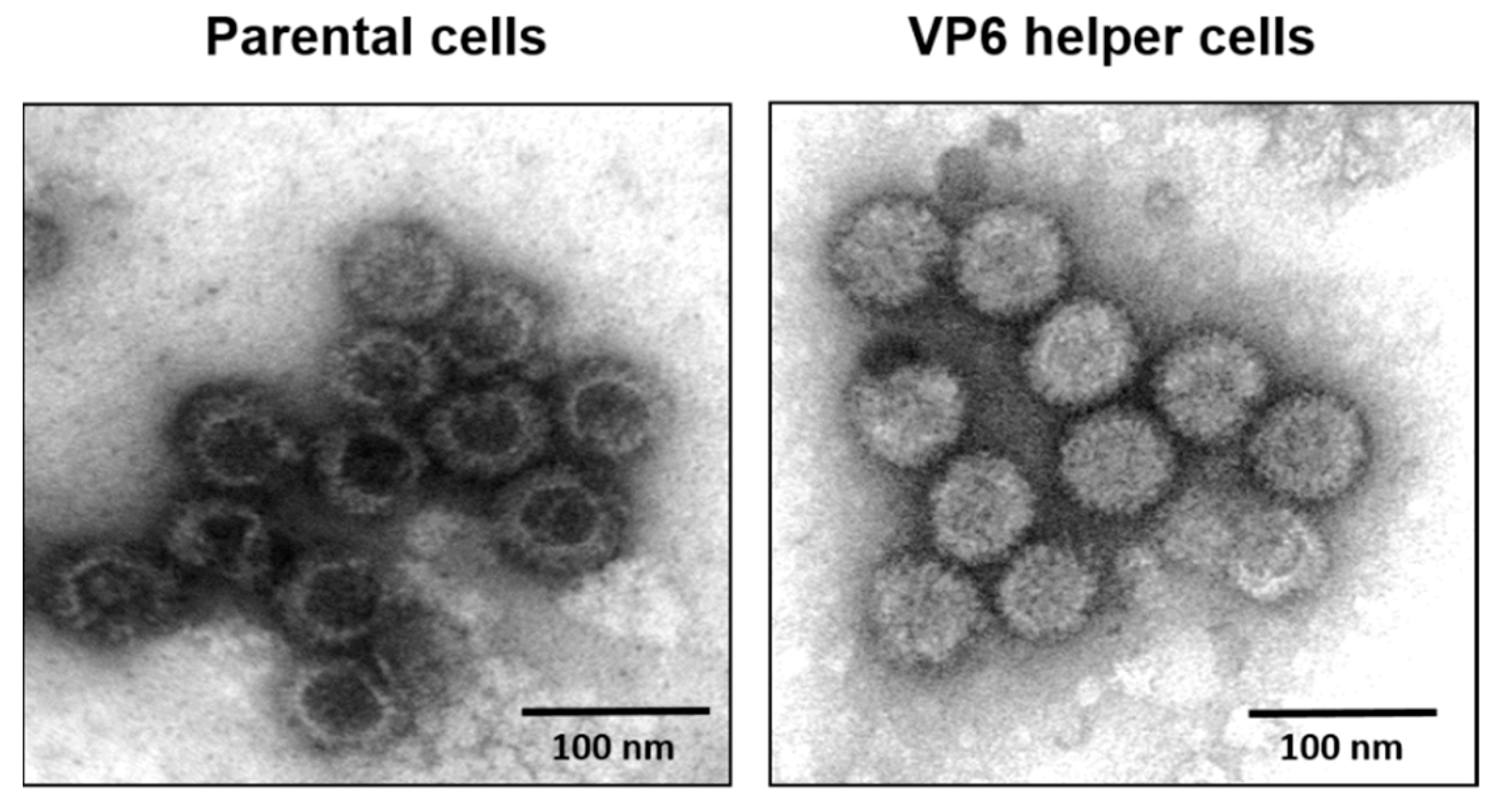
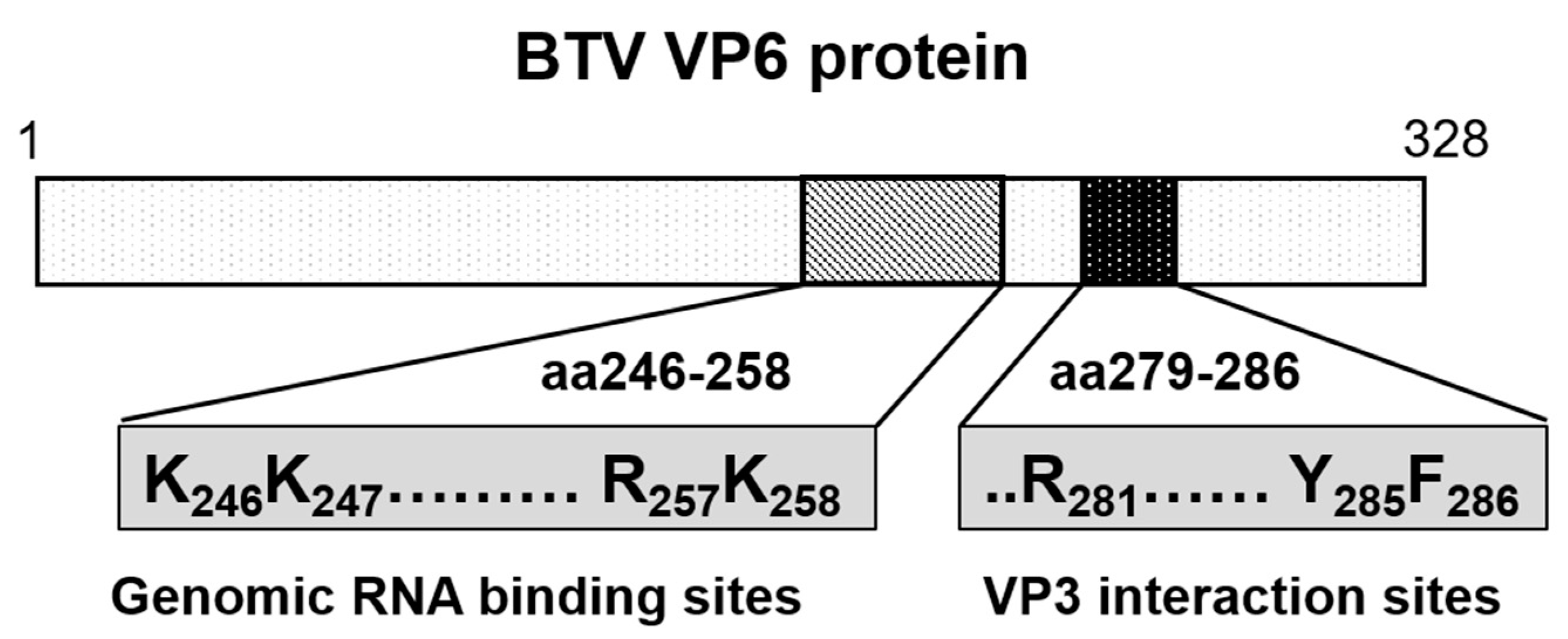
6. Role of Viral Inner Capsid Proteins in RNA Packaging
7. Do Any of the BTV Non-Structural Proteins Play Any Role in RNA Packaging?
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, S.; Rao, V.; Rossmann, M.G. Genome packaging in viruses. Curr. Opin. Struct. Biol. 2010, 20, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Borodavka, A.; Desselberger, U.; Patton, J.T. Genome packaging in multi-segmented dsRNA viruses: Distinct mechanisms with similar outcomes. Curr. Opin. Virol. 2018, 33, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Hanhijärvi, K.J.; Ziedaite, G.; Bamford, D.; Hæggström, E.; Poranen, M. Single-molecule measurements of viral ssRNA packaging. RNA 2016, 23, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luque, D.; Rivas, G.; Alfonso, C.; Carrascosa, J.L.; Rodriguez, J.F.; Caston, J.R. Infectious bursal disease virus is an icosahedral polyploid dsRNA virus. Proc. Natl. Acad. Sci. USA 2009, 106, 2148–2152. [Google Scholar] [CrossRef] [Green Version]
- Schreur, P.J.W.; Kormelink, R.; Kortekaas, J. Genome packaging of the Bunyavirales. Curr. Opin. Virol. 2018, 33, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Hundley, F.; Biryahwaho, B.; Gow, M.; Desselberger, U. Genome rearrangements of bovine rotavirus after serial passage at high multiplicity of infection. Virology 1985, 143, 88–103. [Google Scholar] [CrossRef]
- Joklik, W.K.; Roner, M.R. What reassorts when reovirus genome segments reassort? J. Biol. Chem. 1995, 270, 4181–4184. [Google Scholar] [CrossRef] [Green Version]
- McDonald, S.M.; Patton, J.T. Assortment and packaging of the segmented rotavirus genome. Trends Microbiol. 2011, 19, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Nason, E.L.; Rothagel, R.; Mukherjee, S.K.; Kar, A.K.; Forzan, M.; Prasad, B.V.V.; Roy, P. Interactions between the inner and outer capsids of bluetongue virus. J. Virol. 2004, 78, 8059–8067. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Boyce, M.; Bhattacharya, B.; Schein, S.; Roy, P.; Zhou, Z.H. Bluetongue virus coat protein VP2 contains sialic acid-binding domains, and VP5 resembles enveloped virus fusion proteins. Proc. Natl. Acad. Sci. USA 2010, 107, 6292–6297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Patel, A.; Celma, C.C.; Yu, X.; Roy, P.; Zhou, Z.H. Atomic model of a nonenveloped virus reveals pH sensors for a coordinated process of cell entry. Nat. Struct. Mol. Biol. 2016, 23, 74–80. [Google Scholar] [CrossRef]
- Du, J.; Bhattacharya, B.; Ward, T.; Roy, P. Trafficking of bluetongue virus visualized by recovery of tetracysteine-tagged virion particles. J. Virol. 2014, 88, 12656–12668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.; Roy, P. The molecular biology of Bluetongue virus replication. Virus Res. 2014, 182, 5–20. [Google Scholar] [CrossRef]
- Roy, P. Bluetongue virus structure and assembly. Curr. Opin. Virol. 2017, 24, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Boyce, M.; Wehrfritz, J.; Noad, R.; Roy, P. Purified recombinant bluetongue virus VP1 exhibits RNA replicase activity. J. Virol. 2004, 78, 3994–4002. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, E.; Roy, P. Minimum requirements for bluetongue virus primary replication in vivo. J. Virol. 2013, 87, 882–889. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, E.; Yamazaki, K.; Tsuruta, H.; Roy, P. Interaction between a Unique Minor Protein and a Major Capsid Protein of Bluetongue Virus Controls Virus Infectivity. J. Virol. 2018, 92, e01784-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramadevi, N.; Burroughs, N.J.; Mertens, P.; Jones, I.M.; Roy, P. Capping and methylation of mRNA by purified recombinant VP4 protein of bluetongue virus. Proc. Natl. Acad. Sci. USA 1998, 95, 13537–13542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stäuber, N.; Martínez-Costas, J.; Sutton, G.; Monastyrskaya, K.; Roy, P. Bluetongue virus VP6 protein binds ATP and exhibits an RNA-dependent ATPase function and a helicase activity that catalyze the unwinding of double-stranded RNA substrates. J. Virol. 1997, 71, 7220–7226. [Google Scholar] [CrossRef] [Green Version]
- Sung, P.-Y.; Vaughan, R.; Rahman, S.K.; Yi, G.; Kerviel, A.; Kao, C.C.; Roy, P. The Interaction of Bluetongue Virus VP6 and Genomic RNA Is Essential for Genome Packaging. J. Virol. 2019, 93, e02023-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenco, S.; Roy, P. In vitro reconstitution of Bluetongue virus infectious cores. Proc. Natl. Acad. Sci. USA 2011, 108, 13746–13751. [Google Scholar] [CrossRef] [Green Version]
- Boyce, M.; Celma, C.C.P.; Roy, P. Development of reverse genetics systems for bluetongue virus: Recovery of infectious virus from synthetic RNA transcripts. J. Virol. 2008, 82, 8339–8348. [Google Scholar] [CrossRef] [Green Version]
- Fukusho, A.; Yu, Y.; Yamaguchi, S.; Roy, P. Completion of the sequence of bluetongue virus serotype 10 by the characterization of a structural protein, VP6, and a non-structural protein, NS2. J. Gen. Virol. 1989, 70 Pt 7, 1677–1689. [Google Scholar] [CrossRef]
- Roy, P.; Ritter, G.D.; Akashi, H.; Collisson, E.; Inaba, Y. A genetic probe for identifying bluetongue virus infections in vivo and in vitro. J. Gen. Virol. 1985, 66 Pt 7, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, C.; Sung, P.-Y.; Celma, C.C.; Roy, P. Structural constraints in the packaging of bluetongue virus genomic segments. J. Gen. Virol. 2014, 95 Pt 10, 2240–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, P.-Y.; Roy, P. Sequential packaging of RNA genomic segments during the assembly of Bluetongue virus. Nucleic Acids Res. 2014, 42, 13824–13838. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.J.; Tay, Y. Noncoding RNA: RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Rio, D.C. Mechanisms and Regulation of Alternative Pre-mRNA Splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [Green Version]
- Stadler, P.F. Evolution of RNA-Based Networks. Curr. Top. Microbiol. Immunol. 2015, 392, 43–59. [Google Scholar]
- Borodavka, A.; Dykeman, E.C.; Schrimpf, W.; Lamb, D. Protein-mediated RNA folding governs sequence-specific interactions between rotavirus genome segments. Elife 2017, 6, e27453. [Google Scholar] [CrossRef]
- Newburn, L.R.; White, K.A. Trans-Acting RNA-RNA Interactions in Segmented RNA Viruses. Viruses 2019, 11, 751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo, T., Jr.; Sung, P.Y.; Roy, P. Disruption of Specific RNA-RNA Interactions in a Double-Stranded RNA Virus Inhibits Genome Packaging and Virus Infectivity. PLoS Pathog. 2015, 11, e1005321. [Google Scholar] [CrossRef] [PubMed]
- Fajardo, T., Jr.; AlShaikhahmed, K.; Roy, P. Generation of infectious RNA complexes in Orbiviruses: RNA-RNA interactions of genomic segments. Oncotarget 2016, 7, 72559–72570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo, T., Jr.; Sung, P.Y.; Celma, C.C.; Roy, P. Rotavirus Genomic RNA Complex Forms via Specific RNA-RNA Interactions: Disruption of RNA Complex Inhibits Virus Infectivity. Viruses 2017, 9, 167. [Google Scholar] [CrossRef] [Green Version]
- Alshaikhahmed, K.; Leonov, G.; Sung, P.-Y.; Bingham, R.; Twarock, R.; Roy, P. Dynamic network approach for the modelling of genomic sub-complexes in multi-segmented viruses. Nucleic Acids Res. 2018, 46, 12087–12098. [Google Scholar] [CrossRef]
- Roy, P.; Adachi, A.; Urakawa, T.; Booth, T.F.; Thomas, C.P. Identification of bluetongue virus VP6 protein as a nucleic acid-binding protein and the localization of VP6 in virus-infected vertebrate cells. J. Virol. 1990, 64, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kar, A.K.; Roy, P. Defining the structure-function relationships of bluetongue virus helicase protein VP6. J. Virol. 2003, 77, 11347–11356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celma, C.C.P.; Boyce, M.; van Rijn, P.A.; Eschbaumer, M.; Wernike, K.; Hoffmann, B.; Beer, M.; Haegeman, A.; De Clercq, K.; Roy, P. Rapid generation of replication-deficient monovalent and multivalent vaccines for bluetongue virus: Protection against virulent virus challenge in cattle and sheep. J. Virol. 2013, 87, 9856–9864. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Shivakoti, S.; Ding, K.; Cui, Y.; Roy, P.; Zhou, Z.H. In situ structures of RNA-dependent RNA polymerase inside bluetongue virus before and after uncoating. Proc. Natl. Acad. Sci. USA 2019, 116, 16535–16540. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.P.; Booth, T.F.; Roy, P. Synthesis of bluetongue virus-encoded phosphoprotein and formation of inclusion bodies by recombinant baculovirus in insect cells: It binds the single-stranded RNA species. J. Gen. Virol. 1990, 71, 2073–2083. [Google Scholar] [CrossRef]
- Criglar, J.M.; Anish, R.; Hu, L.; Crawford, S.E.; Sankaran, B.; Prasad, B.V.V.; Estes, M.K. Phosphorylation cascade regulates the formation and maturation of rotaviral replication factories. Proc. Natl. Acad. Sci. USA 2018, 115, E12015–E12023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taraporewala, Z.F.; Patton, J.T. Nonstructural proteins involved in genome packaging and replication of rotaviruses and other members of the Reoviridae. Virus Res. 2004, 101, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lymperopoulos, K.; Wirblich, C.; Brierley, I.; Roy, P. Sequence specificity in the interaction of Bluetongue virus non-structural protein 2 (NS2) with viral RNA. J. Biol. Chem. 2003, 278, 31722–31730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lymperopoulos, K.; Noad, R.; Tosi, S.; Nethisinghe, S.; Brierley, I.; Roy, P. Specific binding of Bluetongue virus NS2 to different viral plus-strand RNAs. Virology 2006, 353, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sung, P.-Y.; Roy, P. RNA Origami: Packaging a Segmented Genome in Orbivirus Assembly and Replication. Viruses 2021, 13, 1841. https://doi.org/10.3390/v13091841
Sung P-Y, Roy P. RNA Origami: Packaging a Segmented Genome in Orbivirus Assembly and Replication. Viruses. 2021; 13(9):1841. https://doi.org/10.3390/v13091841
Chicago/Turabian StyleSung, Po-Yu, and Polly Roy. 2021. "RNA Origami: Packaging a Segmented Genome in Orbivirus Assembly and Replication" Viruses 13, no. 9: 1841. https://doi.org/10.3390/v13091841
APA StyleSung, P.-Y., & Roy, P. (2021). RNA Origami: Packaging a Segmented Genome in Orbivirus Assembly and Replication. Viruses, 13(9), 1841. https://doi.org/10.3390/v13091841