
New Parvoviruses and Picornavirus in Tissues and Feces of Foals with Interstitial Pneumonia

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Samples Collection and Diagnostic Testing
2.2. Metagenomics
2.3. Bioinformatics
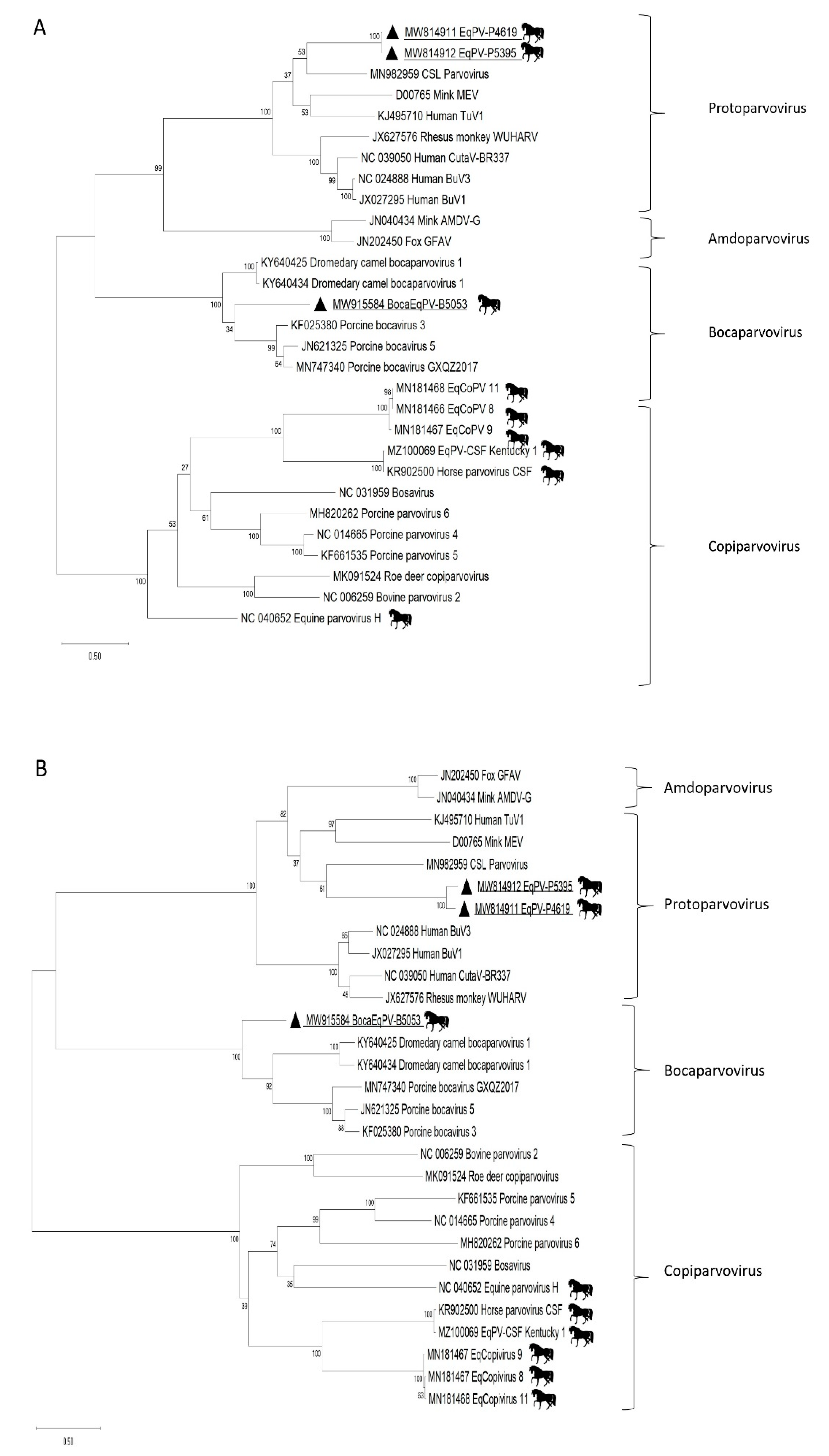
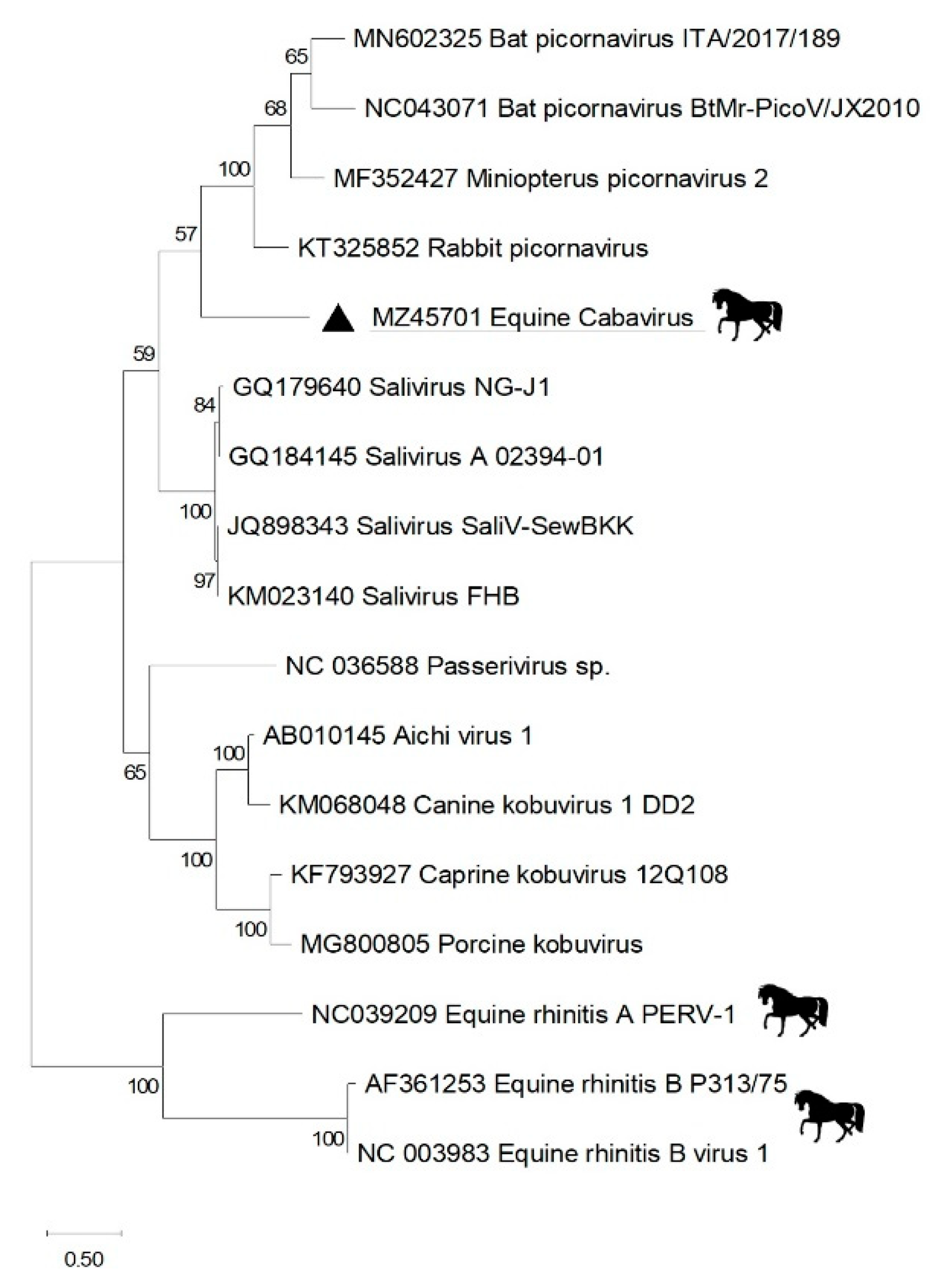
2.4. Phylogenetics
3. Results
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilkerson, J.R.; Bailey, K.E.; Diaz-Mendez, A.; Hartley, C.A. Update on viral diseases of the equine respiratory tract. Vet. Clin. N. Am. Equine Pract. 2015, 31, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Manuja, A.; Gulati, B.R.; Virmani, N.; Tripathi, B.N. Zoonotic Viral Diseases of Equines and Their Impact on Human and Animal Health. Open Virol. J. 2018, 12, 80–98. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.J.; Riddell, S.; Klein, R.; Arkinstall, R.; Haining, J.; Frazer, L.; Mottley, C.; Evans, R.; Johnson, D.; Pallister, J. Experimental Hendra virus infection of dogs: Virus replication, shedding and potential for transmission. Aust Vet. J. 2017, 95, 10–18. [Google Scholar] [CrossRef]
- Pusterla, N.; Vin, R.; Leutenegger, C.M.; Mittel, L.D.; Divers, T.J. Enteric coronavirus infection in adult horses. Vet. J. 2018, 231, 13–18. [Google Scholar] [CrossRef]
- Khusro, A.; Aarti, C.; Rivas-Caceres, R.R.; Barbabosa-Pliego, A. Equine Herpesvirus-I Infection in Horses: Recent Updates on its Pathogenicity, Vaccination, and Preventive Management Strategies. J. Equine Vet. Sci. 2020, 87, 102923. [Google Scholar] [CrossRef]
- Ma, G.; Azab, W.; Osterrieder, N. Equine herpesviruses type 1 (EHV-1) and 4 (EHV-4)--masters of co-evolution and a constant threat to equids and beyond. Vet. Microbiol. 2013, 167, 123–134. [Google Scholar] [CrossRef]
- Hartley, C.A.; Dynon, K.J.; Mekuria, Z.H.; El-Hage, C.M.; Holloway, S.A.; Gilkerson, J.R. Equine gammaherpesviruses: Perfect parasites? Vet. Microbiol. 2013, 167, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Punsmann, S.; Hoppe, J.; Klopfleisch, R.; Venner, M. Acute interstitial pneumonia in foals: A severe, multifactorial syndrome with lung tissue recovery in surviving foals. Equine Vet. J. 2021, 53, 718–726. [Google Scholar] [CrossRef]
- Buergelt, C.D.; Hines, S.A.; Cantor, G.; Stirk, A.; Wilson, J.H. A retrospective study of proliferative interstitial lung disease of horses in Florida. Vet. Pathol. 1986, 23, 750–756. [Google Scholar] [CrossRef]
- Prescott, J.F. Rhodococcus equi: An animal and human pathogen. Clin. Microbiol. Rev. 1991, 4, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Lakritz, J.; Wilson, W.D.; Berry, C.R.; Schrenzel, M.D.; Carlson, G.P.; Madigan, J.E. Bronchointerstitial pneumonia and respiratory distress in young horses: Clinical, clinicopathologic, radiographic, and pathological findings in 23 cases (1984–1989). J. Vet. Int. Med. 1993, 7, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Dunkel, B.; Dolente, B.; Boston, R.C. Acute lung injury/acute respiratory distress syndrome in 15 foals. Equine Vet. J. 2005, 37, 435–440. [Google Scholar] [CrossRef]
- Cann, A.J.; Fandrich, S.E.; Heaphy, S. Analysis of the virus population present in equine faeces indicates the presence of hundreds of uncharacterized virus genomes. Virus Genes 2005, 30, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Steel, O.; Kraberger, S.; Sikorski, A.; Young, L.M.; Catchpole, R.J.; Stevens, A.J.; Ladley, J.J.; Coray, D.S.; Stainton, D.; Dayaram, A.; et al. Circular replication-associated protein encoding DNA viruses identified in the faecal matter of various animals in New Zealand. Infect. Genet Evol. 2016, 43, 151–164. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Li, L.X.; Shi, N.; Sun, X.T.; Liu, Q.; Jin, N.Y.; Si, X.K. First isolation and characterization of Getah virus from cattle in northeastern China. BMC Vet. Res. 2019, 15, 320. [Google Scholar] [CrossRef]
- Chandriani, S.; Skewes-Cox, P.; Zhong, W.; Ganem, D.E.; Divers, T.J.; Van Blaricum, A.J.; Tennant, B.C.; Kistler, A.L. Identification of a previously undescribed divergent virus from the Flaviviridae family in an outbreak of equine serum hepatitis. Proc. Natl. Acad. Sci. USA 2013, 110, E1407–E1415. [Google Scholar] [CrossRef]
- Kapoor, A.; Simmonds, P.; Cullen, J.M.; Scheel, T.K.; Medina, J.L.; Giannitti, F.; Nishiuchi, E.; Brock, K.V.; Burbelo, P.D.; Rice, C.M.; et al. Identification of a pegivirus (GB virus-like virus) that infects horses. J. Virol. 2013, 87, 7185–7190. [Google Scholar] [CrossRef]
- Lyons, S.; Kapoor, A.; Sharp, C.; Schneider, B.S.; Wolfe, N.D.; Culshaw, G.; Corcoran, B.; McGorum, B.C.; Simmonds, P. Nonprimate hepaciviruses in domestic horses, United kingdom. Emerg. Infect. Dis. 2012, 18, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Lyons, S.; Kapoor, A.; Schneider, B.S.; Wolfe, N.D.; Culshaw, G.; Corcoran, B.; Durham, A.E.; Burden, F.; McGorum, B.C.; Simmonds, P. Viraemic frequencies and seroprevalence of non-primate hepacivirus and equine pegiviruses in horses and other mammalian species. J. Gen. Virol. 2014, 95, 1701–1711. [Google Scholar] [CrossRef][Green Version]
- Tomlinson, J.E.; Wolfisberg, R.; Fahnoe, U.; Sharma, H.; Renshaw, R.W.; Nielsen, L.; Nishiuchi, E.; Holm, C.; Dubovi, E.; Rosenberg, B.R.; et al. Equine pegiviruses cause persistent infection of bone marrow and are not associated with hepatitis. PLoS Pathog. 2020, 16, e1008677. [Google Scholar] [CrossRef]
- Divers, T.J.; Tennant, B.C.; Kumar, A.; McDonough, S.; Cullen, J.; Bhuva, N.; Jain, K.; Chauhan, L.S.; Scheel, T.K.H.; Lipkin, W.I.; et al. New Parvovirus Associated with Serum Hepatitis in Horses after Inoculation of Common Biological Product. Emerg. Infect. Dis. 2018, 24, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, J.E.; Jager, M.; Struzyna, A.; Laverack, M.; Fortier, L.A.; Dubovi, E.; Foil, L.D.; Burbelo, P.D.; Divers, T.J.; Van de Walle, G.R. Tropism, pathology, and transmission of equine parvovirus-hepatitis. Emerg. Microbes Infect. 2020, 9, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Giannitti, F.; Low, J.; Keyes, C.; Ullmann, L.S.; Deng, X.; Aleman, M.; Pesavento, P.A.; Pusterla, N.; Delwart, E. Exploring the virome of diseased horses. J. Gen. Virol. 2015, 96, 2721–2733. [Google Scholar] [CrossRef]
- Altan, E.; Li, Y.; Sabino-Santos, G., Jr.; Sawaswong, V.; Barnum, S.; Pusterla, N.; Deng, X.; Delwart, E. Viruses in Horses with Neurologic and Respiratory Diseases. Viruses 2019, 11, 942. [Google Scholar] [CrossRef] [PubMed]
- Hui, A.; Altan, E.; Slovis, N.; Fletcher, C.; Deng, X.; Delwart, E. Circovirus in Blood of a Febrile Horse with Hepatitis. Viruses 2021, 13, 944. [Google Scholar] [CrossRef]
- Smith, K.L.; Li, Y.; Breheny, P.; Cook, R.F.; Henney, P.J.; Sells, S.; Pronost, S.; Lu, Z.; Crossley, B.M.; Timoney, P.J.; et al. New real-time PCR assay using allelic discrimination for detection and differentiation of equine herpesvirus-1 strains with A2254 and G2254 polymorphisms. J. Clin. Microbiol. 2012, 50, 1981–1988. [Google Scholar] [CrossRef]
- Ghoniem, S.M.; El Deeb, A.H.; Aggour, M.G.; Hussein, H.A. Development and evaluation of a multiplex reverse-transcription real-time PCR assay for detection of equine respiratory disease viruses. J. Vet. Diagn. Invest. 2018, 30, 924–928. [Google Scholar] [CrossRef]
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a real-time reverse transcriptase PCR assay for type A influenza virus and the avian H5 and H7 hemagglutinin subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef]
- Altan, E.; Delaney, M.A.; Colegrove, K.M.; Spraker, T.R.; Wheeler, E.A.; Deng, X.; Li, Y.; Gulland, F.M.D.; Delwart, E. Complex Virome in a Mesenteric Lymph Node from a Californian Sea Lion (Zalophus Californianus) with Polyserositis and Steatitis. Viruses 2020, 12, 793. [Google Scholar] [CrossRef]
- Li, L.; Deng, X.; Mee, E.T.; Collot-Teixeira, S.; Anderson, R.; Schepelmann, S.; Minor, P.D.; Delwart, E. Comparing viral metagenomics methods using a highly multiplexed human viral pathogens reagent. J. Virol. Methods 2015, 213, 139–146. [Google Scholar] [CrossRef]
- Brunak, S.; Engelbrecht, J.; Knudsen, S. Prediction of human mRNA donor and acceptor sites from the DNA sequence. J. Mol. Biol. 1991, 220, 49–65. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Tsoi, H.W.; Patteril, N.G.; Yeung, H.C.; Joseph, S.; Wong, E.Y.M.; Muhammed, R.; Chow, F.W.N.; Wernery, U.; et al. Two novel dromedary camel bocaparvoviruses from dromedaries in the Middle East with unique genomic features. J. Gen. Virol. 2017, 98, 1349–1359. [Google Scholar] [CrossRef]
- Patel, J.R.; Heldens, J. Equine herpesviruses 1 (EHV-1) and 4 (EHV-4)--epidemiology, disease and immunoprophylaxis: A brief review. Vet. J. 2005, 170, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Dhama, K.; Karthik, K.; Khandia, R.; Munjal, A.; Khurana, S.K.; Chakraborty, S.; Malik, Y.S.; Virmani, N.; Singh, R.; et al. A Comprehensive Review on Equine Influenza Virus: Etiology, Epidemiology, Pathobiology, Advances in Developing Diagnostics, Vaccines, and Control Strategies. Front. Microbiol. 2018, 9, 1941. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.M. A Brief Introduction to Equine Influenza and Equine Influenza Viruses. Methods Mol. Biol. 2020, 2123, 355–360. [Google Scholar] [CrossRef]
- van Maanen, C. Equine herpesvirus 1 and 4 infections: An update. Vet. Q. 2002, 24, 58–78. [Google Scholar] [CrossRef] [PubMed]
- Plummer, G. An equine respiratory virus with enterovirus properties. Nature 1962, 195, 519–520. [Google Scholar] [CrossRef]
- Studdert, M.J.; Gleeson, L.J. Isolation and characterisation of an equine rhinovirus. Zent. Vet. B 1978, 25, 225–237. [Google Scholar] [CrossRef]
- Wutz, G.; Auer, H.; Nowotny, N.; Grosse, B.; Skern, T.; Kuechler, E. Equine rhinovirus serotypes 1 and 2: Relationship to each other and to aphthoviruses and cardioviruses. J. Gen. Virol. 1996, 77, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Black, W.D.; Wilcox, R.S.; Stevenson, R.A.; Hartley, C.A.; Ficorilli, N.P.; Gilkerson, J.R.; Studdert, M.J. Prevalence of serum neutralising antibody to equine rhinitis A virus (ERAV), equine rhinitis B virus 1 (ERBV1) and ERBV2. Vet. Microbiol. 2007, 119, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Horsington, J.; Lynch, S.E.; Gilkerson, J.R.; Studdert, M.J.; Hartley, C.A. Equine picornaviruses: Well known but poorly understood. Vet. Microbiol. 2013, 167, 78–85. [Google Scholar] [CrossRef]
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus Taxonomy Profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Penzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Penzes, J.J.; Soderlund-Venermo, M.; Canuti, M.; Eis-Hubinger, A.M.; Hughes, J.; Cotmore, S.F.; Harrach, B. Reorganizing the family Parvoviridae: A revised taxonomy independent of the canonical approach based on host association. Arch. Virol. 2020, 165, 2133–2146. [Google Scholar] [CrossRef]
- Christensen, A.; Kesti, O.; Elenius, V.; Eskola, A.L.; Dollner, H.; Altunbulakli, C.; Akdis, C.A.; Soderlund-Venermo, M.; Jartti, T. Human bocaviruses and paediatric infections. Lancet Child Adolesc. Health 2019, 3, 418–426. [Google Scholar] [CrossRef]
- Qiu, J.; Soderlund-Venermo, M.; Young, N.S. Human Parvoviruses. Clin. Microbiol. Rev. 2017, 30, 43–113. [Google Scholar] [CrossRef]
- Ong, D.S.; Schuurman, R.; Heikens, E. Human bocavirus in stool: A true pathogen or an innocent bystander? J. Clin. Virol. 2016, 74, 45–49. [Google Scholar] [CrossRef]
- Manteufel, J.; Truyen, U. Animal bocaviruses: A brief review. Intervirology 2008, 51, 328–334. [Google Scholar] [CrossRef]
- Lu, G.; Wu, L.; Ou, J.; Li, S. Equine Parvovirus-Hepatitis in China: Characterization of Its Genetic Diversity and Evidence for Natural Recombination Events Between the Chinese and American Strains. Front. Vet. Sci. 2020, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Badenhorst, M.; de Heus, P.; Auer, A.; Tegtmeyer, B.; Stang, A.; Dimmel, K.; Tichy, A.; Kubacki, J.; Bachofen, C.; Steinmann, E.; et al. Active equine parvovirus-hepatitis infection is most frequently detected in Austrian horses of advanced age. Equine Vet. J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Meister, T.L.; Tegtmeyer, B.; Bruggemann, Y.; Sieme, H.; Feige, K.; Todt, D.; Stang, A.; Cavalleri, J.V.; Steinmann, E. Characterization of Equine Parvovirus in Thoroughbred Breeding Horses from Germany. Viruses 2019, 11, 965. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ID | Accession | Ranch | Date of Death | Gender | Age | Breed |
|---|---|---|---|---|---|---|
| 1 | S2002675 | A | 27 March 2020 | Female | 3 days | American Quarter Horse |
| 2 | S2003716 | B | 13 May 2020 | Male | 2 month | American Quarter Horse |
| 3 | S2004217 | A | 5 May 2020 | Male | 3.5 month | American Quarter Horse |
| 4 | S2004619 | A | 11 June 2020 | Female | 4 month | American Quarter Horse |
| 5 | S2005053 | C | 28 June 2020 | Male | 2.5 month | Thoroughbred |
| 6 | S2005395 | A | 7 August 2020 | Male | 3.5 month | American Quarter Horse |
| Virus Target | Nested PCR Primers | Amplicon |
|---|---|---|
| Eqcopivirus | EqCopiV F1: CAAGGGACCCGAGCCGCCCC | 514 bp |
| EqCopiV R1: GGGCTGGGGTCTGTGTCCCC | ||
| EqCopiV F2: GAAAATGTAGAGGTAATTGG | ||
| EqCopiV R2: GGAATTCCTCAGGGTTTGCC | ||
| Protoparvoviruses | EqProtoV F1: TCATCATCATGATCTGGGCC | 253 bp |
| EqProtoV R1: ACCCAGTTGCCAAATTTGCC | ||
| EqProtoV F2: TCTATCCGCCAGGTCAGTGG | ||
| EqProtoV R2: TGATAAAGTTCACGTCTGCC | ||
| Bocaparvovirus | EqBocaV F1: GGCCCTTGTTGCACTTGTGG | 984 bp |
| EqBocaV R1: GCTGCGTTTACAGGCTCCCC | ||
| EqBocaV F2: CACACTACTTCTCCAGACGG | ||
| EqBocaV R2: CCTGGAAATAACCACCCTCC | ||
| Picornavirus | EqPicoV F1: CTACCCATGGTCGGCTAAGG | 306 bp |
| EqPicoV R1: TTCGTACGATGCGAAGTCCC | ||
| EqPicoV F2: GGAGGACCAGACGTTCACTG | ||
| EqPicoV R2: AGAAGCGAGTCCAGTCCAC |
| ID | Accession | Dx | Bacteriology | EHV-1 PCR (Lung) | EHV-4 PCR (Lung) | Influenza PCR (Lung) | VI (Lung) | Other Tests and Relevant Findings |
|---|---|---|---|---|---|---|---|---|
| 1 | S2002675 | Int Pneum | Aerobic: mixed flora (liver), no growth (lung) Salmonella culture: Not detected (liver) | NEG | NEG | ND | ND | HMS + Se: Suboptimal Se, 0.21 ppm (Ref range 0.3–1.0); liver |
| 2 | S2003716 | Int Pneum | Aerobic: No growth (lung) | NEG | NEG | NEG | ND | HMS + Se: Suboptimal Se, 0.22 ppm (Ref range 0.3–1.0); liver |
| 3 | S2004217 | Int Pneum | Aerobic: E. coli and Enterococcus hirae (liver), mixed flora (lung), E. coli and mixed flora (small intestine), E. coli and mixed flora (colon), E. hirae and E. coli (abdomen), Staph haemolyticus, mixed flora, E. coli (joint) Anaerobic: No growth (small intestine, colon) C. diff culture: Not detected (small intestine, colon) Salmonella culture: Not detected (liver, small intestine, colon) | NEG | NEG | NEG | ND | Fecal float: No parasite eggs/oocysts detected HMS + Se: Acceptable/non-diagnostic ranges; liver Peritonitis with GI content (perforation) |
| 4 | S2004619 | Int Pneum | Aerobic: No growth (lung, liver) Salmonella culture: Not detected (liver) | NEG | NEG | NEG | NEG | HMS + Se: Acceptable/non-diagnotic ranges; liver Fecal float: Parascaris eggsSevere ascariasis |
| 5 | S2005053 | Int Pneum | Aerobic: No growth (lung, liver) Salmonella culture: Not detected (liver) | NEG | NEG | NEG | ND | Fecal float: No parasite eggs/oocysts detected HMS + Se: Suboptimal Se, 0.23 ppm (Ref range 0.3–1.0); liver |
| 6 | S2005395 | Int Pneum | Aerobic: E. coli (liver), Acinetobacter Iwoffi and mixed flora (lung) Salmonella culture: Not detected (small intestine) | NEG | ND | NEG | NEG | Fecal float: No parasite eggs/oocysts detected HMS + Se: Acceptable/non-diagnostic ranges; liver |
| ID | Accession | Ranch | Tissue | Bocavirus | Eqcopivirus | Protoparvovirus | Picornavirus |
|---|---|---|---|---|---|---|---|
| 1 | S2005395 | A | Lung | Negative | Negative | Negative | Negative |
| Spleen | Negative | Negative | Positive | Positive | |||
| Colon Content | Negative | Negative | Positive | Positive | |||
| 2 | S2002675 | B | Lung | Negative | Negative | Negative | Negative |
| Spleen | Negative | Negative | Positive | Negative | |||
| 3 | S2003716 | A | Lung | Negative | Negative | Negative | Negative |
| Spleen | Negative | Negative | Negative | Negative | |||
| Colon Content | Negative | Negative | Negative | Negative | |||
| 4 | S2004619 | A | Lung | Negative | Negative | Positive | Negative |
| Spleen | Negative | Negative | Positive | Negative | |||
| Colon Content | Negative | Negative | Positive | Negative | |||
| 5 | S2004217 | C | Lung | Negative | Positive | Negative | Negative |
| Spleen | Negative | Positive | Negative | Negative | |||
| Colon Content | Negative | Positive | Negative | Positive | |||
| 6 | S2005053 | A | Lung | Negative | Negative | Negative | Positive |
| Spleen | Positive | Negative | Negative | Negative | |||
| Colon Content | Positive | Negative | Negative | Negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altan, E.; Hui, A.; Li, Y.; Pesavento, P.; Asín, J.; Crossley, B.; Deng, X.; Uzal, F.A.; Delwart, E. New Parvoviruses and Picornavirus in Tissues and Feces of Foals with Interstitial Pneumonia. Viruses 2021, 13, 1612. https://doi.org/10.3390/v13081612
Altan E, Hui A, Li Y, Pesavento P, Asín J, Crossley B, Deng X, Uzal FA, Delwart E. New Parvoviruses and Picornavirus in Tissues and Feces of Foals with Interstitial Pneumonia. Viruses. 2021; 13(8):1612. https://doi.org/10.3390/v13081612
Chicago/Turabian StyleAltan, Eda, Alvin Hui, Yanpeng Li, Patricia Pesavento, Javier Asín, Beate Crossley, Xutao Deng, Francisco A. Uzal, and Eric Delwart. 2021. "New Parvoviruses and Picornavirus in Tissues and Feces of Foals with Interstitial Pneumonia" Viruses 13, no. 8: 1612. https://doi.org/10.3390/v13081612
APA StyleAltan, E., Hui, A., Li, Y., Pesavento, P., Asín, J., Crossley, B., Deng, X., Uzal, F. A., & Delwart, E. (2021). New Parvoviruses and Picornavirus in Tissues and Feces of Foals with Interstitial Pneumonia. Viruses, 13(8), 1612. https://doi.org/10.3390/v13081612