
Pathology and Pathogenesis of Eurasian Blackbirds (Turdus merula) Naturally Infected with Usutu Virus

 ,
,  ,
,  , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Virology
Virus Detection and Sequencing
2.3. Pathology
2.4. Immunohistochemistry (IHC)
2.5. Statistical Analysis
3. Results
3.1. Animals
3.2. Virology
Virus Detection and Lineage Determination
3.3. Pathology
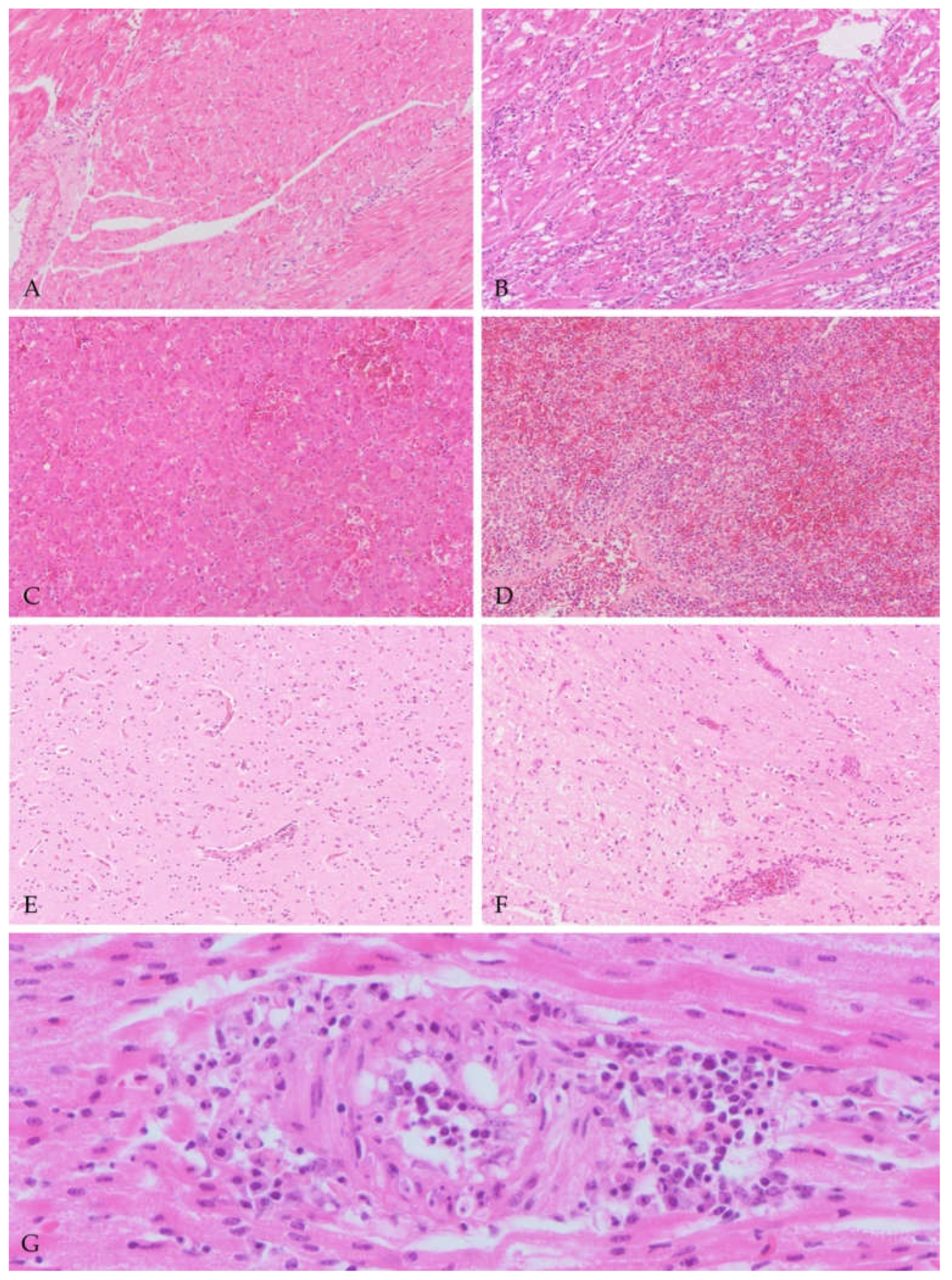
3.3.1. Macroscopic and Histologic Findings
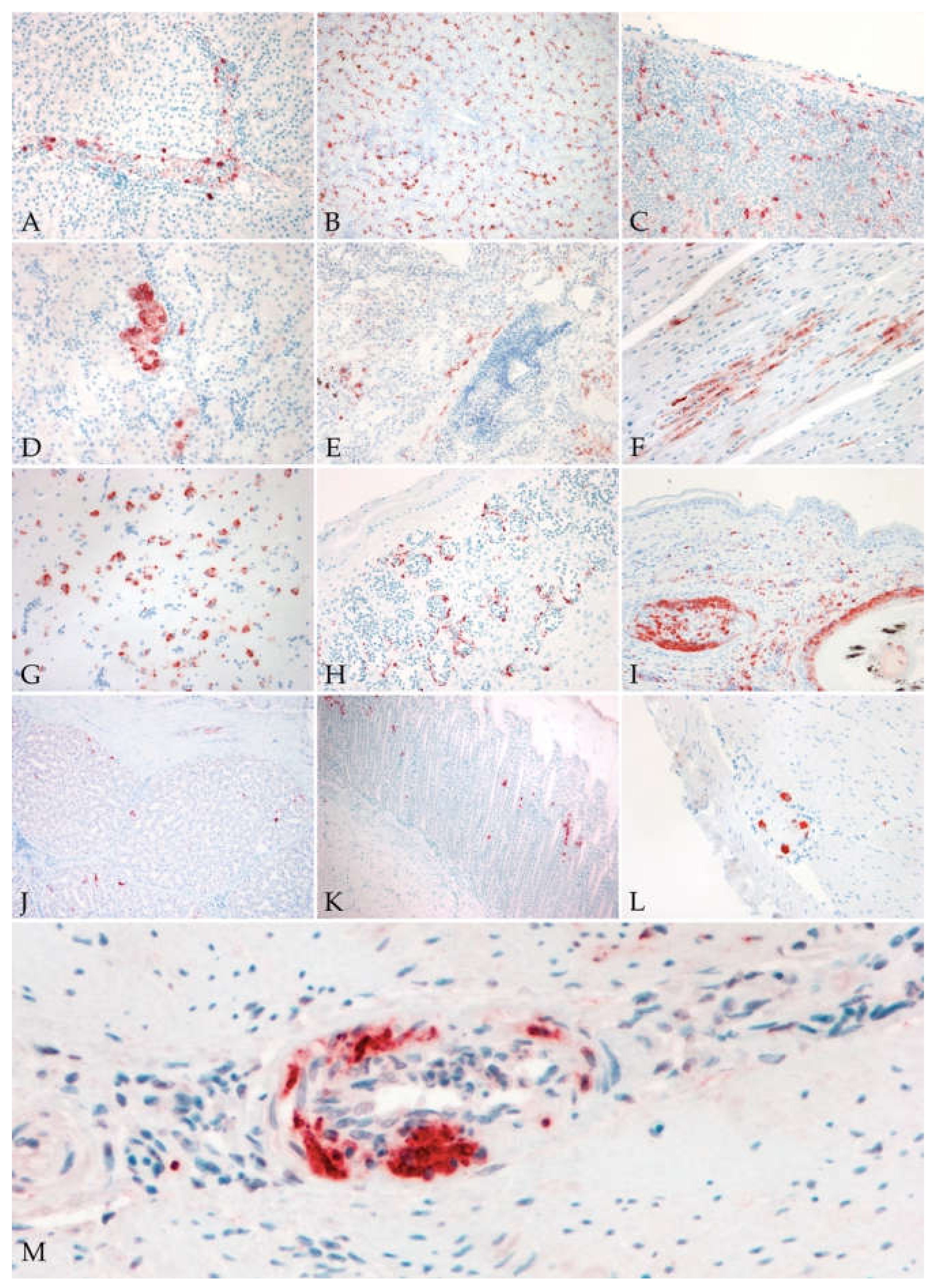
3.3.2. Immunohistochemistry
3.3.3. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rijks, J.M.; Kik, M.; Slaterus, R.; Foppen, R.; Stroo, A.; Ijzer, J.; Stahl, J.; Gröne, A.; Koopmans, M.; van der Jeugd, H.; et al. Widespread Usutu virus outbreak in birds in The Netherlands. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Gaibani, P.; Rossini, G. An overview of Usutu virus. Microbes Infect. 2017, 19, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Benzarti, E.; Garigliany, M. In Vitro and In Vivo Models to Study the Zoonotic Mosquito-Borne Usutu Virus. Viruses 2020, 12, 1116. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-bauder, B.; Nowotny, N. Emergence of Usutu virus, an African Mosquito-Borne Flavivirus of the Japanese Encephalitis Virus Group, Central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Manarolla, G.; Bakonyi, T.; Gallazzi, D.; Crosta, L.; Weissenböck, H.; Dorrestein, G.M.; Nowotny, N. Usutu virus in wild birds in northern Italy. Vet. Microbiol. 2010, 141, 159–163. [Google Scholar] [CrossRef]
- Ziegler, U.; Jöst, H.; Müller, K.; Fischer, D.; Rinder, M.; Tietze, D.T.; Danner, K.-J.; Becker, N.; Skuballa, J.; Hamann, H.-P.; et al. Epidemic Spread of Usutu Virus in Southwest Germany in 2011 to 2013 and Monitoring of Wild Birds for Usutu and West Nile Viruses. Vector-Borne Zoonotic Dis. 2015, 15, 481–488. [Google Scholar] [CrossRef]
- Nikolay, B.; Diallo, M.; Faye, O.; Boye, C.S.; Sall, A.A. Vector competence of Culex neavei (Diptera: Culicidae) for Usutu virus. Am. J. Trop. Med. Hyg. 2012, 86, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Pacenti, M.; Sinigaglia, A.; Martello, T.; De Rui, M.E.; Franchin, E.; Pagni, S.; Peta, E.; Riccetti, S.; Milani, A.; Montarsi, F.; et al. Clinical and virological findings in patients with Usutu virus infection, northern Italy, 2018. Eurosurveillance 2019, 24, 1900180. [Google Scholar] [CrossRef]
- Allering, L.; Jöst, H.; Emmerich, P.; Günther, S.; Lattwein, E.; Schmidt, M.; Seifried, E.; Sambri, V.; Hourfar, K.; Schmidt-Chanasit, J. Detection of Usutu virus infection in a healthy blood donor from south-west Germany, 2012. Eurosurveillance 2012, 17. [Google Scholar] [CrossRef]
- Aberle, S.W.; Kolodziejek, J.; Jungbauer, C.; Stiasny, K.; Aberle, J.H.; Zoufaly, A.; Hourfar, M.K.; Weidner, L.; Nowotny, N. Increase in human west nile and usutu virus infections, Austria, 2018. Eurosurveillance 2018, 23, 3–8. [Google Scholar] [CrossRef]
- Zaaijer, H.L.; Slot, E.; Molier, M.; Reusken, C.B.E.M.; Koppelman, M.H.G.M. Usutu virus infection in Dutch blood donors. Transfusion 2019, 59, 2931–2937. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Jungbauer, C.; Aberle, S.W.; Kolodziejek, J.; Dimmel, K.; Stiasny, K.; Allerberger, F.; Nowotny, N. Usutu virus infections among blood donors, Austria, July and August 2017—Raising awareness for diagnostic challenges. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Domanović, D.; Gossner, C.M.; Lieshout-Krikke, R.; Mayr, W.; Baroti-Toth, K.; Dobrota, A.M.; Escoval, M.A.; Henseler, O.; Jungbauer, C.; Liumbruno, G.; et al. West Nile and Usutu Virus Infections and Challenges to Blood Safety in the European Union. Emerg. Infect. Dis. 2019, 25. [Google Scholar] [CrossRef]
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef]
- Llorente, F.; García-Irazábal, A.; Pérez-Ramírez, E.; Cano-Gómez, C.; Sarasa, M.; Vázquez, A.; Jiménez-Clavero, M.Á. Influence of flavivirus co-circulation in serological diagnostics and surveillance: A model of study using West Nile, Usutu and Bagaza viruses. Transbound. Emerg. Dis. 2019, tbed.13262. [Google Scholar] [CrossRef]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu Virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Oude Munnink, B.B.; Münger, E.; Nieuwenhuijse, D.F.; Kohl, R.; van der Linden, A.; Schapendonk, C.M.E.; van der Jeugd, H.; Kik, M.; Rijks, J.M.; Reusken, C.B.E.M.; et al. Genomic monitoring to understand the emergence and spread of Usutu virus in the Netherlands, 2016–2018. Sci. Rep. 2020, 10, 2798. [Google Scholar] [CrossRef]
- Chvala, S.; Kolodziejek, J.; Nowotny, N.; Weissenböck, H. Pathology and viral distribution in fatal Usutu virus infections of birds from the 2001 and 2002 outbreaks in Austria. J. Comp. Pathol. 2004, 131, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Erdélyi, K.; Brunthaler, R.; Dán, Á.; Weissenböck, H.; Nowotny, N. Usutu virus, Austria and Hungary, 2010–2016. Emerg. Microbes Infect. 2017, 6, 1–7. [Google Scholar] [CrossRef]
- Steinmetz, H.W.; Bakonyi, T.; Weissenböck, H.; Hatt, J.M.; Eulenberger, U.; Robert, N.; Hoop, R.; Nowotny, N. Emergence and establishment of Usutu virus infection in wild and captive avian species in and around Zurich, Switzerland-Genomic and pathologic comparison to other central European outbreaks. Vet. Microbiol. 2011, 148, 207–212. [Google Scholar] [CrossRef]
- Garigliany, M.; Linden, A.; Gilliau, G.; Levy, E.; Sarlet, M.; Franssen, M.; Benzarti, E.; Derouaux, A.; Francis, F.; Desmecht, D. Usutu virus, Belgium, 2016. Infect. Genet. Evol. 2017, 48, 116–119. [Google Scholar] [CrossRef]
- Ziegler, U.; Fast, C.; Eiden, M.; Bock, S.; Schulze, C.; Hoeper, D.; Ochs, A.; Schlieben, P.; Keller, M.; Zielke, D.E.; et al. Evidence for an independent third Usutu virus introduction into Germany. Vet. Microbiol. 2016, 192, 60–66. [Google Scholar] [CrossRef]
- Bakonyi, T.; Lussy, H.; Weissenböck, H.; Hornyák, Á.; Nowotny, N. In vitro host-cell susceptibility to usutu virus. Emerg. Infect. Dis. 2005, 11, 298–301. [Google Scholar] [CrossRef]
- Kuchinsky, S.C.; Hawks, S.A.; Mossel, E.C.; Coutermarsh-Ott, S.; Duggal, N.K. Differential pathogenesis of Usutu virus isolates in mice. PLoS Negl. Trop. Dis. 2020, 14, e0008765. [Google Scholar] [CrossRef]
- Clé, M.; Constant, O.; Barthelemy, J.; Desmetz, C.; Martin, M.F.; Lapeyre, L.; Cadar, D.; Savini, G.; Teodori, L.; Monaco, F.; et al. Differential neurovirulence of Usutu virus lineages in mice and neuronal cells. J. Neuroinflamm. 2021, 18, 11. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Bakonyi, T.; Chvala, S.; Nowotny, N. Experimental Usutu virus infection of suckling mice causes neuronal and glial cell apoptosis and demyelination. Acta Neuropathol. 2004, 108, 453–460. [Google Scholar] [CrossRef]
- Chvala, S.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; Weissenböck, H. Limited pathogenicity of Usutu virus for the domestic chicken (Gallus domesticus). Avian Pathol. 2005, 34, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Chvala, S.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; Weissenböck, H. Limited pathogenicity of usutu virus for the domestic goose (Anser anser f. domestica) following experimental inoculation. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 2006, 53, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, B.; Weidmann, M.; Dupressoir, A.; Faye, O.; Boye, C.S.; Diallo, M.; Sall, A.A. Development of a Usutu virus specific real-time reverse transcription PCR assay based on sequenced strains from Africa and Europe. J. Virol. Methods 2014, 197, 51–54. [Google Scholar] [CrossRef]
- Jöst, H.; Bialonski, A.; Maus, D.; Sambri, V.; Eiden, M.; Groschup, M.H.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation of Usutu Virus in Germany. Am. J. Trop. Med. Hyg. 2011, 85, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Oude Munnink, B.B.; Kik, M.; de Bruijn, N.D.; Kohl, R.; van der Linden, A.; Reusken, C.B.E.M.; Koopmans, M. Towards high quality real-time whole genome sequencing during outbreaks using Usutu virus as example. Infect. Genet. Evol. 2019, 73, 49–54. [Google Scholar] [CrossRef]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for Valid Histopathologic Scoring in Research. Vet. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Van Riel, D.; Van Den Brand, J.M.A.; Munster, V.J.; Besteboer, T.M.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Pathology and Virus Distribution in Chickens Naturally Infected with Highly Pathogenic Avian Influenza A Virus (H7N7) During the 2003 Outbreak in The Netherlands. Vet. Pathol. 2009, 46, 971–976. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using IBM SPSS Statistics; Sage Publications Ltd.: London, UK, 2009; ISBN 9781446249185. [Google Scholar]
- Beaman, M.; Madge, S. The Handbook of Bird Identification; A&C Black: London, UK, 1998; ISBN 978-0-8117-3196-6. [Google Scholar]
- Becker, N.; Jöst, H.; Ziegler, U.; Eiden, M.; Höper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic Emergence of Usutu Virus in Wild and Captive Birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Meister, T.; Lussy, H.; Bakonyi, T.; Šikutová, S.; Rudolf, I.; Vogl, W.; Winkler, H.; Frey, H.; Hubálek, Z.; Nowotny, N. Serological evidence of continuing high Usutu virus (Flaviviridae) activity and establishment of herd immunity in wild birds in Austria. Vet. Microbiol. 2008, 127, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Höfle, U.; Gamino, V.; Fernández de Mera, I.G.; Mangold, A.J.; Ortíz, J.A.; de la Fuente, J. Usutu virus in migratory song thrushes, Spain. Emerg. Infect. Dis. 2013, 19, 1173–1175. [Google Scholar] [CrossRef]
- Gamino, V.; Höfle, U. Pathology and tissue tropism of natural West Nile virus infection in birds: A review. Vet. Res. 2013, 44, 39. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.; Gould, D.; Bowen, R.; Komar, N. Natural and experimental West Nile virus infection in five raptor species. J. Wildl. Dis. 2006, 42, 1–13. [Google Scholar] [CrossRef]
- Ernest, H.B.; Woods, L.W.; Hoar, B.R. Pathology associated with west nile virus infections in the yellow-billed magpie (PICA nuttalli): A california endemic bird. J. Wildl. Dis. 2010, 46, 401–408. [Google Scholar] [CrossRef]
- Harroud, A.; Almutlaq, A.; Pellerin, D.; Paz, D.; Linnell, G.J.; Gendron, D. West Nile virus-associated vasculitis and intracranial hemorrhage. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e641. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, B.A.; Newman, T.L.; Tresley, D.J. Occlusive retinal vasculitis in a patient with West Nile virus. Clin. Exp. Optom. 2007, 90, 463–467. [Google Scholar] [CrossRef]
- Medina, L. Do birds and reptiles possess homologues of mammalian visual, somatosensory, and motor cortices? Evol. Nerv. Syst. 2007, 2, 163–194. [Google Scholar] [CrossRef]
- Stacho, M.; Herold, C.; Rook, N.; Wagner, H.; Axer, M.; Amunts, K.; Güntürkün, O. A cortex-like canonical circuit in the avian forebrain. Science 2020, 369. [Google Scholar] [CrossRef]
- Reiner, A.; Yamamoto, K.; Karten, H.J. Organization and evolution of the avian forebrain. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2005, 287, 1080–1102. [Google Scholar] [CrossRef]
- Marszalek, R.; Chen, A.Y.-Y.; Gjede, J. Psoriasiform eruption in the setting of West Nile virus. J. Am. Acad. Dermatol. 2014, 70, AB4. [Google Scholar] [CrossRef]
- Palmieri, C.; Franca, M.; Uzal, F.; Anderson, M.; Barr, B.; Woods, L.; Moore, J.; Woolcock, P.; Shivaprasad, H.L.L. Pathology and immunohistochemical findings of west nile virus infection in psittaciformes. Vet. Pathol. 2011, 48, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Redfern, C.P.F. Brood-patch development and female body mass in passerines. Ringing Migr. 2010, 25, 33–41. [Google Scholar] [CrossRef]
- Docherty, D.E.; Long, R.R.; Griffin, K.M.; Saito, E.K. Corvidae Feather Pulp and West Nile Virus Detection. Emerg. Infect. Dis. 2004, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.M.; Young, G.R.; Burkhalter, K.L.; Brault, A.C.; Reisen, W.K.; Komar, N. West Nile virus detection in nonvascular feathers from avian carcasses. J. Vet. Diagn. Investig. 2009, 21, 616–622. [Google Scholar] [CrossRef]
- Benzarti, E.; Rivas, J.; Sarlet, M.; Franssen, M.; Desmecht, D.; Schmidt-Chanasit, J.; Savini, G.; Lorusso, A.; Van Laere, A.S.; Garigliany, M.M. Experimental usutu virus infection in domestic canaries serinus canaria. Viruses 2020, 12, 164. [Google Scholar] [CrossRef]
- Nuradji, H.; Bingham, J.; Payne, J.; Harper, J.; Lowther, S.; Wibawa, H.; Long, N.T.; Meers, J. Highly Pathogenic Avian Influenza (H5N1) Virus in Feathers: Tropism and Pathology of Virus-Infected Feathers of Infected Ducks and Chickens. Vet. Pathol. 2017, 54, 226–233. [Google Scholar] [CrossRef]
- Mercer, J.; Greber, U.F. Virus interactions with endocytic pathways in macrophages and dendritic cells. Trends Microbiol. 2013, 21, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, E.A.; Bowen, R.A.; Brault, A.C. An ex vivo avian leukocyte culture model for West Nile virus infection. J. Virol. Methods 2015, 218, 19–22. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rios, M.; Zhang, M.J.; Grinev, A.; Srinivasan, K.; Daniel, S.; Wood, O.; Hewlett, I.K.; Dayton, A.I. Monocytes-macrophages are a potential target in human infection with West Nile virus through blood transfusion. Transfusion 2006, 46, 659–667. [Google Scholar] [CrossRef]
- Nikitina, E.; Larionova, I.; Choinzonov, E.; Kzhyshkowska, J. Monocytes and macrophages as viral targets and reservoirs. Int. J. Mol. Sci. 2018, 19, 2821. [Google Scholar] [CrossRef] [PubMed]
- Suen, W.W.; Prow, N.A.; Hall, R.A.; Bielefeldt-Ohmann, H. Mechanism of west nile virus neuroinvasion: A critical appraisal. Viruses 2014, 6, 2796–2825. [Google Scholar] [CrossRef]
- Luo, H.; Winkelmann, E.R.; Zhu, S.; Ru, W.; Mays, E.; Silvas, J.A.; Vollmer, L.L.; Gao, J.; Peng, B.H.; Bopp, N.E.; et al. Peli1 facilitates virus replication and promotes neuroinflammation during West Nile virus infection. J. Clin. Invest. 2018, 128, 4980–4991. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Kolodziejek, J.; Fragner, K.; Kuhn, R.; Pfeffer, M.; Nowotny, N. Usutu virus activity in Austria, 2001–2002. Microbes Infect. 2003, 5, 1132–1136. [Google Scholar] [CrossRef]
- Simonin, Y.; Sillam, O.; Carles, M.J.; Gutierrez, S.; Gil, P.; Constant, O.; Martin, M.F.; Grard, G.; Van de Perre, P.; Salinas, S.; et al. Human Usutu Virus Infection with Atypical Neurologic Presentation, Montpellier, France, 2016. Emerg. Infect. Dis. 2018, 24, 875–878. [Google Scholar] [CrossRef]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef]
- McLean, R.G.; Ubico, S.R.; Docherty, D.E.; Hansen, W.R.; Sileo, L.; McNamara, T.S. West Nile virus transmission and ecology in birds. Ann. N. Y. Acad. Sci. 2001, 951, 54–57. [Google Scholar] [CrossRef]
- Sbrana, E.; Tonry, J.H.; Xiao, S.Y.; Travassos Da Rosa, A.P.A.; Higgs, S.; Tesh, R.B. Oral transmission of West Nile virus in a hamster model. Am. J. Trop. Med. Hyg. 2005, 72, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Reperant, L.A.; van de Bildt, M.W.G.; van Amerongen, G.; Leijten, L.M.E.; Watson, S.; Palser, A.; Kellam, P.; Eissens, A.C.; Frijlink, H.W.; Osterhaus, A.D.M.E.; et al. Marked Endotheliotropism of Highly Pathogenic Avian Influenza Virus H5N1 following Intestinal Inoculation in Cats. J. Virol. 2012, 86, 1158–1165. [Google Scholar] [CrossRef] [PubMed]
- Bertelsen, M.F.; Ølberg, R.A.; Cramshaw, G.J.; Dibernardo, A.; Lindsay, L.R.; Drebot, M.; Barker, I.K. West Nile virus infection in the eastern loggerhead shrike (Lanius ludovicianus migrans): Pathology, epidemiology, and immunization. J. Wildl. Dis. 2004, 40, 538–542. [Google Scholar] [CrossRef][Green Version]
- Maria Gershovitz, A.B. Sub Acute Thyroiditis in a Case of West Nile Virus (WNV) Infection. J. Clin. Case Rep. 2013, 03. [Google Scholar] [CrossRef]
- Bakonyi, T.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Csörgo, T.; Lussy, H.; Chvala, S.; Bukovsky, C.; Meister, T.; Weissenböck, H.; et al. Emergence of usutu virus in Hungary. J. Clin. Microbiol. 2007, 45, 3870–3874. [Google Scholar] [CrossRef] [PubMed]
- Rouffaer, L.O.; Steensels, M.; Verlinden, M.; Vervaeke, M.; Boonyarittichaikij, R.; Martel, A.; Lambrecht, B. Usutu Virus Epizootic and Plasmodium Coinfection in Eurasian Blackbirds (Turdus merula) in Flanders, Belgium. J. Wildl. Dis. 2018, 54, 859–862. [Google Scholar] [CrossRef]
- Nemeth, N.M.; Bosco-Lauth, A.M.; Williams, L.M.; Bowen, R.A.; Brown, J.D. West Nile Virus Infection in Ruffed Grouse (Bonasa umbellus): Experimental Infection and Protective Effects of Vaccination. Vet. Pathol. 2017, 54, 901–911. [Google Scholar] [CrossRef]
- Nemeth, N.M.; Thomsen, B.V.; Spraker, T.R.; Benson, J.M.; Bosco-Lauth, A.M.; Oesterle, P.T.; Bright, J.M.; Muth, J.P.; Campbell, T.W.; Gidlewski, T.L.; et al. Clinical and pathologic responses of american crows (corvus brachyrhynchos) and fish crows (c ossifragus) to experimental west nile virus infection. Vet. Pathol. 2011, 48, 1061–1074. [Google Scholar] [CrossRef]
- Peter Clement, R.H. Thrushes. Available online: https://books.google.nl/books?hl=en&lr=&id=WAZCBAAAQBAJ&oi=fnd&pg=PA3&dq=Thrushes+(Helm+Identification+Guides)+Clement,+Peter&ots=Hgo5dofhES&sig=aMHmaO89rC2yMpOz3eh6GVwwu4c&redir_esc=y#v=onepage&q=Thrushes(HelmIdentificationGuides)Clement%2CPeter&f=f (accessed on 26 October 2020).



{kind=link}
{kind=link}
| USUV RT-PCR | Ct Values | |
|---|---|---|
| USUV Positive * | 118/160 | 21.62 ± 4.63 |
| Europe 3 | 24/118 | 20.84 ± 4.14 |
| Africa 3 | 80/118 | 20.99 ± 3.65 |
| Organ | Lesion | N° | % |
|---|---|---|---|
| Liver | Necrosis | 118/143 | 82 |
| Hepatitis | 118/143 | 82 | |
| Spleen | Necrosis | 118/142 | 83 |
| Splenitis | 85/142 | 59 | |
| Heart | Necrosis | 97/145 | 66 |
| Myocarditis | 126/145 | 86 | |
| Vasculitis | 3/145 | 2 | |
| Lung | Pneumonia | 128/139 | 92 |
| Cerebrum | Necrosis | 59/145 | 40 |
| Perivascular cuffs | 104/145 | 71 | |
| Vasculitis | 69/145 | 47 |
| Organ | Grade 0 * | Grade 1 | Grade 2 | Grade 3 | |||||
|---|---|---|---|---|---|---|---|---|---|
| N° | % | N° | % | N° | % | N° | % | ||
| Liver | Necrosis | 25/143 | 17 | 77/143 | 53 | 35/143 | 24 | 6/143 | 4 |
| Hepatitis | 25/143 | 17 | 69/143 | 48 | 46/143 | 32 | 3/143 | 2 | |
| Spleen | Necrosis | 24/142 | 16 | 35/142 | 24 | 61/142 | 43 | 22/142 | 15 |
| Splenitis | 57/142 | 40 | 41/142 | 28 | 40/142 | 28 | 4/142 | 2 | |
| Heart | Necrosis | 48/145 | 33 | 62/145 | 42 | 32/145 | 22 | 3/145 | 2 |
| Myocarditis | 19/145 | 13 | 58/145 | 40 | 60/145 | 41 | 8/145 | 5 | |
| Lung | Pneumonia | 11/139 | 7 | 69/139 | 49 | 51/139 | 36 | 8/139 | 5 |
| Brain | Encephalitis | 41/145 | 28 | 52/145 | 35 | 27/145 | 18 | 3/145 | 17 |
| Vasculitis | 76/145 | 52 | 37/145 | 25 | 26/145 | 17 | 6/145 | 4 | |
| Necrosis | 86/145 | 53 | 48/145 | 33 | 10/145 | 6 | 1/145 | 1 | |
| Organ | Tissue/Cell | Immunolabeling * |
|---|---|---|
| All organs | Endothelial cells Vascular wall Mononucleated cells Intravascular leukocytes | +++ ++ ++ + |
| Liver | Kupffer’s cells Hepatocytes | +++ + |
| Spleen | Capsular stromal cells High walled veins | ++ + |
| Heart | Cardiomyocytes Endocardium | +++ ++ |
| Lung | Type I pneumocytes | + |
| Air sacs | Spindle stromal cells | + |
| Kidney | Tubular epithelium Glomeruli | +++ + |
| Skin | Keratinocytes Feathers | + ++ |
| Cerebrum | Neurons Glial cells | +++ + |
| Cerebellum | Neurons Glial cells | +++ + |
| Proventriculus | Mucosal epithelium Smooth muscle cells Peripheral ganglion | ++ + + |
| Gizzard | Mucosal epithelium Smooth muscle cells Peripheral ganglion | ++ + + |
| Intestine | Mucosal epithelium Smooth muscle cells Peripheral ganglion | ++ + + |
| Gonads | Sertoli cells Ovary follicular epithelium Germinal cells | + + + |
| Thyroid | Follicular epithelium | ++ |
| Skeletal muscle | Stromal cells | + |
| Organ | Grade 0 | Grade 1 | Grade 2 | Grade 3 | ||||
|---|---|---|---|---|---|---|---|---|
| N° | % | N° | % | N° | % | N° | % | |
| Liver | 59/137 | 43 | 35/137 | 25 | 10/137 | 7 | 33/137 | 24 |
| Spleen | 58/125 | 46 | 26/125 | 20 | 12/125 | 9 | 29/125 | 23 |
| Heart | 45/133 | 33 | 25/133 | 18 | 10/133 | 7 | 53/133 | 35 |
| Kidney | 57/138 | 41 | 30/138 | 21 | 15/138 | 10 | 36/138 | 26 |
| Lung | 69/128 | 53 | 25/128 | 19 | 9/128 | 7 | 25/128 | 19 |
| Air sacs | 7/14 | 50 | 3/14 | 21 | 0/14 | 0 | 4/14 | 28 |
| Cerebrum | 60/136 | 44 | 17/136 | 12 | 8/136 | 5 | 51/136 | 37 |
| Cerebellum | 60/98 | 61 | 14/98 | 14 | 12/98 | 12 | 12/98 | 12 |
| Skin | 27/82 | 32 | 6/82 | 7 | 4/82 | 4 | 45/82 | 54 |
| Gizzard | 88/126 | 69 | 10/126 | 7 | 9/126 | 7 | 19/126 | 15 |
| Proventriculus | 79/105 | 75 | 8/105 | 7 | 7/105 | 6 | 11/105 | 10 |
| Intestine | 65/104 | 62 | 4/104 | 3 | 11/104 | 10 | 24/104 | 23 |
| Pancreas | 22/31 | 70 | 6/31 | 19 | 0/31 | 0 | 3/31 | 9 |
| Gonads | 44/65 | 67 | 5/65 | 7 | 4/65 | 6 | 12/65 | 18 |
| Thyroids | 47/68 | 69 | 5/68 | 7 | 4/68 | 5 | 12/68 | 17 |
| Bursa of Fabricius | 11/15 | 73 | 1/15 | 6 | 0/15 | 0 | 3/15 | 20 |
| Thymus | 0/2 | 0 | 0/2 | 0 | 0/2 | 0 | 0/2 | 0 |
| Skeletal muscle | 0/1 | 0 | 0/1 | 0 | 1/1 | 100 | 0/1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giglia, G.; Agliani, G.; Munnink, B.B.O.; Sikkema, R.S.; Mandara, M.T.; Lepri, E.; Kik, M.; Ijzer, J.; Rijks, J.M.; Fast, C.; et al. Pathology and Pathogenesis of Eurasian Blackbirds (Turdus merula) Naturally Infected with Usutu Virus. Viruses 2021, 13, 1481. https://doi.org/10.3390/v13081481
Giglia G, Agliani G, Munnink BBO, Sikkema RS, Mandara MT, Lepri E, Kik M, Ijzer J, Rijks JM, Fast C, et al. Pathology and Pathogenesis of Eurasian Blackbirds (Turdus merula) Naturally Infected with Usutu Virus. Viruses. 2021; 13(8):1481. https://doi.org/10.3390/v13081481
Chicago/Turabian StyleGiglia, Giuseppe, Gianfilippo Agliani, Bas B. Oude Munnink, Reina S. Sikkema, Maria Teresa Mandara, Elvio Lepri, Marja Kik, Jooske Ijzer, Jolianne M. Rijks, Christine Fast, and et al. 2021. "Pathology and Pathogenesis of Eurasian Blackbirds (Turdus merula) Naturally Infected with Usutu Virus" Viruses 13, no. 8: 1481. https://doi.org/10.3390/v13081481
APA StyleGiglia, G., Agliani, G., Munnink, B. B. O., Sikkema, R. S., Mandara, M. T., Lepri, E., Kik, M., Ijzer, J., Rijks, J. M., Fast, C., Koopmans, M. P. G., Verheije, M. H., Gröne, A., Reusken, C. B. E. M., & van den Brand, J. M. A. (2021). Pathology and Pathogenesis of Eurasian Blackbirds (Turdus merula) Naturally Infected with Usutu Virus. Viruses, 13(8), 1481. https://doi.org/10.3390/v13081481