
The Comparison of Honeybee Viral Loads for Six Honeybee Viruses (ABPV, BQCV, CBPV, DWV, LSV3 and SBV) in Healthy and Clinically Affected Honeybees with TaqMan Quantitative Real-Time RT-PCR Assays


Abstract
:1. Introduction
2. Materials and Methods
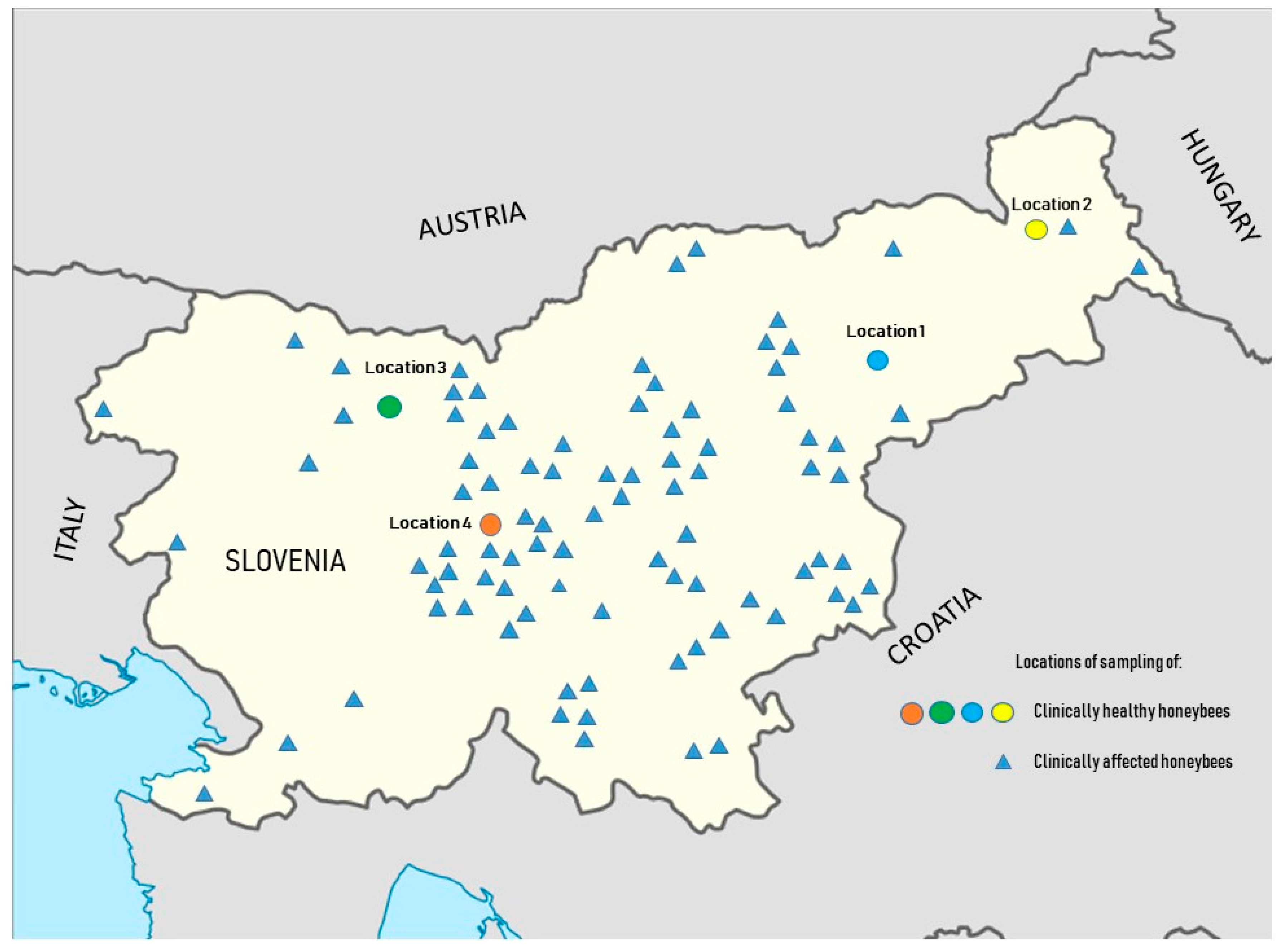
2.1. Sample Collection
2.2. Sample Preparation
2.3. Primers, TaqMan Probes, and Standards Design
2.4. RNA Extraction and RT-qPCR Assays
2.5. Statistical Analysis
3. Results
3.1. The Performance of RT-qPCR Assays for Six Honeybee Viruses
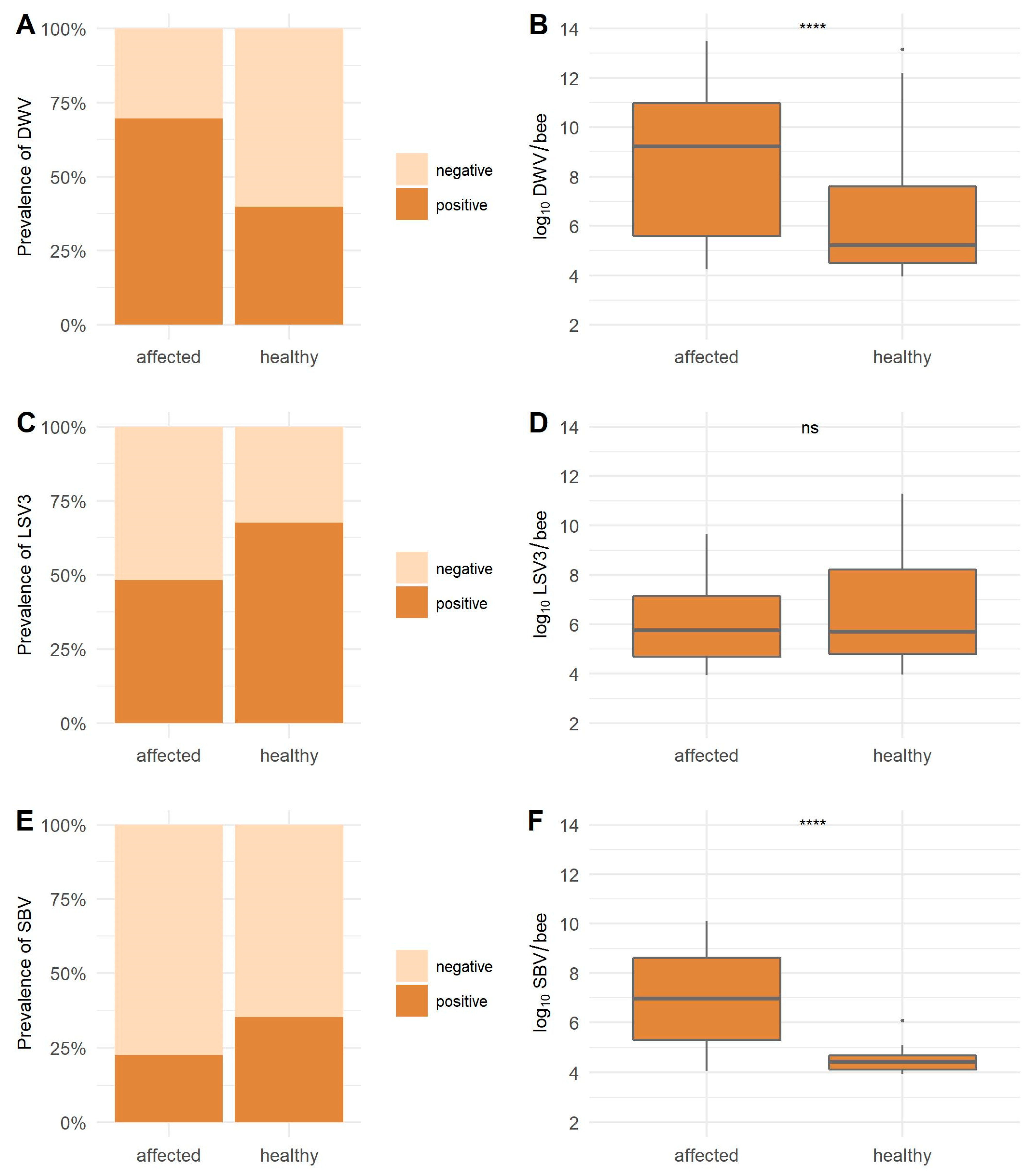
3.2. Viral Prevalence and Comparison of the Viral Loads from Samples of Healthy and Clinically Affected Honeybee Colonies
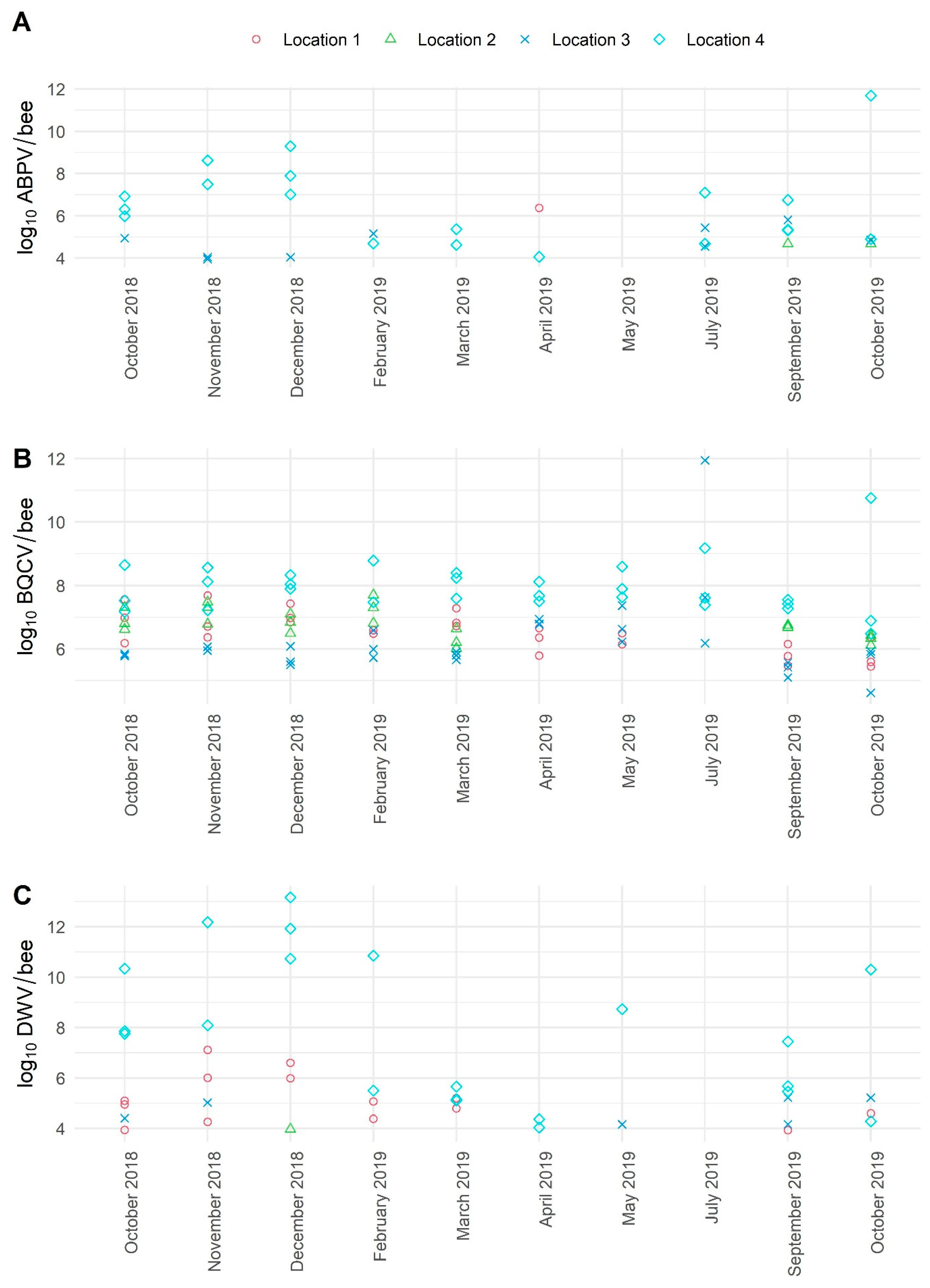
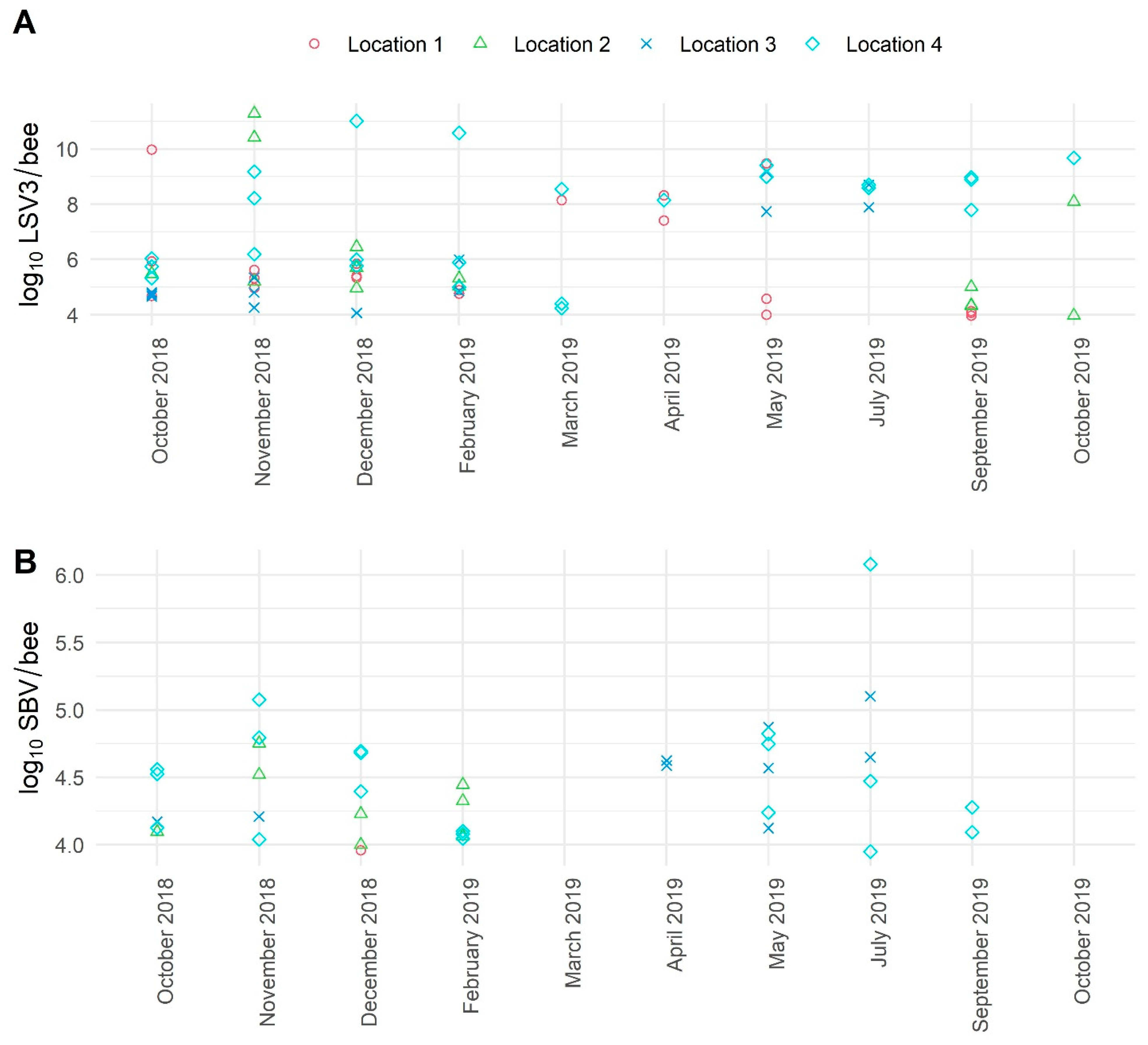
3.3. The Average Viral Load Variations in Positive Samples of Healthy Honeybees Collected in a 10-Month Period
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, J.D.; Schwarz, R.S. Bees brought to their knees: Microbes affecting honey bee health. Trends Microbiol. 2011, 19, 614–620. [Google Scholar] [CrossRef]
- Fürst, M.A.; McMahon, D.P.; Osborne, J.L.; Paxton, R.J.; Brown, M.J.F. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef]
- Manley, R.; Boots, M.; Wilfert, L. Condition-dependent virulence of slow bee paralysis virus in Bombus terrestris: Are the impacts of honeybee viruses in wild pollinators underestimated? Oecologia 2017, 184, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Toplak, I.; Šimenc, L.; Pislak Ocepek, M.; Bevk, D. Determination of genetically identical strains of four honeybee viruses in bumblebee positive samples. Viruses 2020, 12, 1310. [Google Scholar] [CrossRef]
- Runckel, C.; Flenniken, M.L.; Engel, J.C.; Ruby, J.G.; Ganem, D.; Andino, R.; DeRisi, J.L. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia. PLoS ONE 2011, 6, e20656. [Google Scholar] [CrossRef] [Green Version]
- Remnant, E.J.; Shi, M.; Buchmann, G.; Blacquière, T.; Holmes, E.C.; Beekman, M.; Ashe, A. A diverse range of novel RNA viruses in geographically distinct honey bee populations. J. Virol. 2017, 91, e00158-17. [Google Scholar] [CrossRef] [Green Version]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, pathogenicity, and impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L.; Ball, B.V. Viruses. In Honey Bee Pathology, 2nd ed.; Academic Press: Cambridge, MA, USA, 1991; pp. 10–34. [Google Scholar] [CrossRef]
- Ribière, M.; Triboulot, C.; Mathieu, L.; Aurières, C.; Faucon, J.P.; Pépin, M. Molecular diagnosis of chronic bee paralysis virus infection. Apidologie 2002, 33, 339–351. [Google Scholar] [CrossRef]
- Daughenbaugh, K.F.; Martin, M.; Brutscher, L.M.; Cavigli, I.; Garcia, E.; Lavin, M.; Flenniken, M.L. Honey bee infecting lake sinai viruses. Viruses 2015, 7, 3285–3309. [Google Scholar] [CrossRef] [Green Version]
- Bacandritsos, N.; Granato, A.; Budge, G.; Papanastasiou, I.; Roinioti, E.; Caldon, M.; Falcaro, C.; Gallina, A.; Mutinelli, F. Sudden deaths and colony population decline in Greek honey bee colonies. J. Inverteb. Pathol. 2010, 105, 335–340. [Google Scholar] [CrossRef]
- Blanchard, P.; Regnault, J.; Schurr, F.; Dubois, E.; Ribière, M. Intra-laboratory validation of chronic bee paralysis virus quantitation using an accredited standardised real-time quantitative RT-PCR method. J. Virol. Methods 2012, 180, 26–31. [Google Scholar] [CrossRef]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [Green Version]
- Jamnikar Ciglenečki, U.; Toplak, I. Genetic diversity of acute bee paralysis virus in Slovenian honeybee samples. Acta Vet. Hung. 2013, 61, 244–256. [Google Scholar] [CrossRef]
- Doublet, V.; Labarussias, M.; de Miranda, J.R.; Moritz, R.F.; Paxton, R.J. Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 2015, 17, 969–983. [Google Scholar] [CrossRef]
- Faucon, J.P.; Mathieu, L.; Ribiere, M.P.; Drajnudel, A.C.; Zeggane, S.; Aurieres, C.; Aubert, M.F.A. Honey bee winter mortality in France in 1999 and 2000. Bee World 2002, 83, 14–23. [Google Scholar] [CrossRef]
- Antúnez, K.; Alessandro, B.D.; Corbella, E.; Zunino, P. Detection of chronic bee paralysis virus and acute bee paralysis virus in Uruguayan honeybees. J. Invertebr. Pathol. 2005, 90, 69–72. [Google Scholar] [CrossRef]
- Schurr, F.; Cougoule, N.; Rivière, M.P.; Ribière-Chabert, M.; Achour, H.; Ádám, D.; Castillo, C.; de Graaf, D.C.; Forsgren, E.; Granato, A.; et al. Trueness and precision of the real-time RT-PCR method for quantifying the chronic bee paralysis virus genome in bee homogenates evaluated by a comparative inter-laboratory study. J. Virol. Methods 2017, 248, 217–225. [Google Scholar] [CrossRef]
- Toplak, I.; Jamnikar-Ciglenečki, U.; Aronstein, K.; Gregorc, A. Chronic bee paralysis virus and Nosema ceranae experimental co-infection of winter honey bee workers (Apis mellifera L.). Viruses 2013, 5, 2282–2297. [Google Scholar] [CrossRef] [Green Version]
- de Miranda, J.R.; Cordoni, G.; Budge, G. The Acute bee paralysis virus-Kashmir bee virus-Israeli acute paralysis virus complex. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S30–S34. [Google Scholar] [CrossRef]
- Benaets, K.; Van Geystelen, A.; Cardoen, D.; De Smet, L.; de Graaf, D.C.; Schoofs, L.; Larmuseau, M.H.; Brettell, L.E.; Martin, S.J.; Wenseleers, T. Covert deformed wing virus infections have long-term deleterious effects on honeybee foraging and survival. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed]
- Brettell, L.E.; Mordecai, G.J.; Schroeder, D.C.; Jones, I.M.; da Silva, J.R.; Vicente-Rubiano, M.; Martin, S.J. A Comparison of deformed wing virus in deformed and asymptomatic honey bees. Insects 2017, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamnikar-Ciglenečki, U.; Pislak Ocepek, M.; Toplak, I. Genetic diversity of deformed wing virus rom Apis mellifera carnica (Hymenoptera: Apidae) and Varroa Mite (Mesostigmata: Varroidae). J. Econ. Entomol. 2018, 112, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Cornman, R.S. Relative abundance and molecular evolution of Lake Sinai Virus (Sinaivirus) clades. PeerJ 2019, 7, e6305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigot, D.; Dalmon, A.; Roy, B.; Hou, C.; Germain, M.; Romary, M.; Deng, S.; Diao, Q.; Weinert, L.A.; Cook, J.M.; et al. The discovery of Halictivirus resolves the Sinaivirus phylogeny. J. Gen. Virol. 2017, 98, 2864–2875. [Google Scholar] [CrossRef]
- Traynor, K.S.; Rennich, K.; Forsgren, E.; Rose, R.; Pettis, J.; Kunkel, G.; Madella, S.; Evans, J.; Lopez, D.; van Engelsdorp, D. Multiyear survey targeting disease incidence in US honey bees. Apidologie 2016, 47, 325–347. [Google Scholar] [CrossRef] [Green Version]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; van Engelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef] [Green Version]
- Šimenc, L.; Kuhar, U.; Jamnikar-Ciglenečki, U.; Toplak, I. First Complete Genome of Lake Sinai virus lineage 3 and genetic diversity of Lake Sinai virus strains from honey bees and bumblebees. J. Econ. Entomol. 2020, 113, 1055–1061. [Google Scholar] [CrossRef]
- Bailey, L. The multiplication and spread of sacbrood virus of bees. Ann. Appl. Biol. 1969, 63, 483–491. [Google Scholar] [CrossRef]
- Schurr, F.; Tison, A.; Militano, L.; Cheviron, N.; Sircoulomb, F.; Rivière, M.P.; Ribière-Chabert, M.; Thiéry, R.; Dubois, E. Validation of quantitative real-time RT-PCR assays for the detection of six honeybee viruses. J. Virol. Methods 2019, 270, 70–78. [Google Scholar] [CrossRef]
- Jamnikar-Ciglenečki, U.; Pislak-Ocepek, M.; Toplak, I. Seasonal variations of four honey bee viruses in pupae, hive and forager bees of Carniolan gray bee (Apis mellifera carnica). Slo. Vet. Res. 2014, 51, 131–140. [Google Scholar]
- Jamnikar-Ciglenečki, U.; Toplak, I. Development of a real-time RT-PCR assay with TaqMan probe for specific detection of acute bee paralysis virus. J. Virol. Methods 2012, 184, 63–68. [Google Scholar] [CrossRef]
- Forsgren, E.; Locke, B.; Semberg, E.; Laugen, A.T.; Miranda, J.R. Sample preservation, transport and processing strategies for honeybee RNA extraction: Influence on RNA yield, quality, target quantification and data normalisation. J. Virol. Methods 2017, 246, 81–89. [Google Scholar] [CrossRef]
- Chantawannakul, P.; Ward, L.; Boonham, N.; Brown, M. A scientific note on the detection of honeybee viruses using real-time PCR (TaqMan) in Varroa mites collected from a Thai honeybee (Apis mellifera) apiary. J. Invertebr. Pathol. 2006, 91, 69–73. [Google Scholar] [CrossRef]
- Bakonyi, T.; Grabensteiner, E.; Kolodziejek, J.; Rusvai, M.; Topolska, G.; Ritter, W.; Nowotny, N. Phylogenetic analysis of acute bee paralysis virus strains. Appl. Environ. Microbiol. 2002, 68, 6446–6450. [Google Scholar] [CrossRef] [Green Version]
- Benjeddou, M.; Leat, N.; Allsopp, M.; Davison, S. Detection of acute bee paralysis virus and black queen cell virus from honeybees by reverse transcriptase pcr. Appl. Environ. Microbiol. 2001, 67, 2384–2387. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, P.; Ribiere, M.; Celle, O.; Lallemand, P.; Schurr, F.; Olivier, V.; Iscache, A.L.; Faucon, J.P. Evaluation of a real-time two-step RT-PCR assay for quantitation of chronic bee paralysis virus (CBPV) genome in experimentally-infected bee tissues and in life stages of a symptomatic colony. J. Virol. Methods 2007, 141, 7–13. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2019; Available online: http://www.r-project.org/index.html (accessed on 10 July 2021).
- Sprent, P. Fisher Exact Test. In International Encyclopaedia of Statistical Science; Lovric, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef] [Green Version]
- Haynes, W. Wilcoxon Rank Sum Test. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.H., Yokota, H., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef] [Green Version]
- AFNOR. NF U47-600, Part 2: Animal Health Analysis Methods-PCR-Part2: Requirements and recommendations for the Development and the VALIDATION of veterinary PCR, Norme Françaises. 2015, pp. 1–51. Available online: https://www.boutique.afnor.org (accessed on 10 June 2021).
- Desai, S.D.; Kumar, S.; Currie, R. Occurrence, detection, and quantification of economically important viruses in healthy and unhealthy honey bee (Hymenoptera: Apidae) colonies in Canada. Can. Entomol. 2015, 148, 22–35. [Google Scholar] [CrossRef]
- Molineri, A.I.; Pacini, A.; Giacobino, A.; Bulacio-Cagnolo, A.; Aignasse, A.; Zago, L.; Fondevila, N.; Ferrufino, C.; Merke, J.; Orellano, E.; et al. Prevalence of honey bee (Apis mellifera) viruses in temperate and subtropical regions from Argentina. Rev. Argent. Microbiol. 2017, 49, 166–173. [Google Scholar] [CrossRef]
- Blanchard, P.; Guillot, S.; Antùnez, K.; Köglberger, H.; Kryger, P.; de Miranda, J.R.; Franco, S.; Chauzat, M.P.; Thiéry, R.; Ribière, M. Development and validation of a real-time two-step RT-qPCR TaqMan(®) assay for quantitation of Sacbrood virus (SBV) and its application to a field survey of symptomatic honey bee colonies. J. Virol Methods 2014, 197, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Reid, S.M.; Ferris, N.P.; Hutchings, G.H.; Zhang, Z.; Belsham, G.J.; Alexandersen, S. Detection of all seven serotyes of foot-and-mouth disease virus by real-time, fluorogenic reverse transcription polymerase chain reaction assay. J. Virol. Methods 2002, 105, 67–80. [Google Scholar] [CrossRef]
- Lakshmi, I.K.; Putty, K.; Raut, S.S. Standardization and application of real-time polymerase chain reaction for rapid detection of bluetongue virus. Vet. World 2018, 11, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, B.; Beer, M.; Schelp, C.; Schirrmeier, H.; Depner, K. Validation of a real-time RT-PCR assay for sensitive and specific detection of classical swine fever. J. Virol. Methods 2005, 130, 36–44. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Probe (5′-3′) | Reference | Primer Concentration | Probe Concentration |
|---|---|---|---|---|---|---|
| ABPV | CATATTGGCGAGCCACTATG | CTACCAGGTTCAAAGAAAATTTC | (6-Fam) ATAGTTAAAACAGCTTTTCACACTGG (Tamra) | [32] | 800 mM | 100 nM |
| BQCV | GGTGCGGGAGATGATATATGGA | GCCGTCTGAGATGCATGAATAC | (6-Fam) TTTCCATCTTTATCGGTACGCCGCC (Tamra) | [34] | 320 nM | 200 nM |
| CBPV | CGCAAGTACGCCTTGATAAAGAAC | ACTACTAGAAACTCGTCGCTTCG | (6-Fam) TCAAGAACGAGACCACCGCCAAGTTC (Tamra) | [12] | 320 nM | 200 nM |
| DWV | GCGGCTAAGATTGTAAATGTC | GTGACTAGCATAACCATGATTA | (6-Fam) CCTTGACCAGTAGACAGCATC (Tamra) | [30] | 350 nM | 100 nM |
| LSV3 | GTACCTACACTCTTCCATGCAC | CCAGCTAAGGAGGCGTAAC | (Cy5) TCACCCACATCATTGAGCCAGGT (BHQ2) | This study | 200 nM | 100 nM |
| SBV | AGCCAGTGATAGATGCTC | AAATACTCCCGCCAAATCAC | (6-Fam) TGGCTCATCTGGGATCACAATTTCC (Tamra) | This study | 200 nM | 50 nM |
| Standard Name | Plasmid Vector (Supplier) | Cloned Sequence: Nucleotide Position | Sequence Length | Virus (GenBank Accession Number) | Reference |
|---|---|---|---|---|---|
| pB2 | pCR II Topo (Invitrogen, Waltham, MA, USA) | 8115 to 8512 | 397 | ABPV (AF126050) | [35] |
| pNC1-4 | pGEM-T Easy (Promega, Madison, WI, USA) | 7850 to 8550 | 700 | BQCV (AF183905) | [36] |
| pAb2 | pGEM-T Easy (Promega, Madison, WI, USA) | 2260 to 3059 | 799 | CBPV (EU122229) | [37] |
| pC1 | pCR II Topo (Invitrogen, Waltham, MA, USA) | 4240 to 4659 | 419 | DWV (AY292384) | [37] |
| LSV3 | LSV3-SI (Sigma-Aldrich, Steinheim, Germany) | 2021 to 2265 | 244 | LSV3 (KY465717) | This study |
| SBV | SBV-SI (Sigma-Aldrich, Steinheim, Germany) | 5030 to 5368 | 338 | SBV (MG545287) | This study |
| ABPV | Repeat 1 | Repeat 2 | Repeat 3 | Mean | SD |
|---|---|---|---|---|---|
| Intercept | 43.96 | 48.47 | 45.35 | 45.93 | ±2.54 |
| Slope | −3.084 | −3.345 | −3.150 | −3.193 | ±0.152 |
| Efficiency | 111.0 | 99.0 | 107.7 | 105.9 | ±6.9 |
| R2 | 0.995 | 0.978 | 0.982 | 0.985 | ±0.010 |
| BQCV | Repeat 1 | Repeat 2 | Repeat 3 | Mean | SD |
| Intercept | 42.77 | 41.80 | 41.77 | 42.11 | ±0.66 |
| Slope | −3.362 | −3.102 | −3.036 | −3.167 | ±0.195 |
| Efficiency | 99.2 | 110.1 | 113.5 | 107.6 | ±8.4 |
| R2 | 0.993 | 0.994 | 1.000 | 0.996 | ±0.004 |
| CBPV | Repeat 1 | Repeat 2 | Repeat 3 | Mean | SD |
| Intercept | 48.35 | 48.04 | 46.05 | 47.48 | ±1.43 |
| Slope | −3.579 | −3.432 | −3.358 | −3.456 | ±0.123 |
| Efficiency | 90.3 | 95.9 | 98.5 | 94.9 | ±4.6 |
| R2 | 0.995 | 0.996 | 1.000 | 0.997 | ±0.003 |
| DWV | Repeat 1 | Repeat 2 | Repeat 3 | Mean | SD |
| Intercept | 42.74 | 42.65 | 40.38 | 41.92 | ±1.54 |
| Slope | −3.334 | −3.059 | −3.011 | −3.135 | ±0.199 |
| Efficiency | 99.5 | 112.3 | 114.8 | 108.9 | ±9.37 |
| R2 | 0.981 | 0.998 | 0.997 | 0.992 | ±0.011 |
| LSV 3 | Repeat 1 | Repeat 2 | Repeat 3 | Mean | SD |
| Intercept | 42.52 | 41.04 | 39.58 | 41.05 | ±1.47 |
| Slope | −3.395 | −3.607 | −3.460 | −3.487 | ±0.120 |
| Efficiency | 97.0 | 89.9 | 94.5 | 93.8 | ±3.9 |
| R2 | 0.997 | 0.998 | 0.995 | 0.997 | ±0.002 |
| SBV | Repeat 1 | Repeat 2 | Repeat 3 | Mean | SD |
| Intercept | 41.08 | 40.29 | 37.68 | 39.68 | ±2.00 |
| Slope | −3.616 | −3.517 | −3.264 | −3.466 | ±0.202 |
| Efficiency | 89.0 | 92.5 | 102.5 | 94.7 | ±7.8 |
| R2 | 0.997 | 0.999 | 1.000 | 0.999 | ±0.002 |
| Location No. No. of Samples | ABPV | BQCV | CBPV | DWV | LSV3 | SBV |
|---|---|---|---|---|---|---|
| Location 1—healthy n = 27 | 3.70 | 96.30 | 0.00 | 55.56 | 74.07 | 3.70 |
| Location 2—healthy n = 21 | 9.52 | 100 | 0.00 | 4.76 | 71.43 | 38.10 |
| Location 3—healthy n = 30 | 30.00 | 100 | 0.00 | 20.00 | 46.67 | 30.00 |
| Location 4—healthy n = 30 | 66.67 | 96.67 | 0.00 | 70.00 | 80.00 | 66.67 |
| Total—healthy n = 108 | 29.63 | 98.15 | 0.00 | 39.81 | 67.59 | 35.19 |
| Total—affected n = 89 | 79.78 | 96.63 | 19.10 | 69.66 | 48.31 | 22.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šimenc, L.; Knific, T.; Toplak, I. The Comparison of Honeybee Viral Loads for Six Honeybee Viruses (ABPV, BQCV, CBPV, DWV, LSV3 and SBV) in Healthy and Clinically Affected Honeybees with TaqMan Quantitative Real-Time RT-PCR Assays. Viruses 2021, 13, 1340. https://doi.org/10.3390/v13071340
Šimenc L, Knific T, Toplak I. The Comparison of Honeybee Viral Loads for Six Honeybee Viruses (ABPV, BQCV, CBPV, DWV, LSV3 and SBV) in Healthy and Clinically Affected Honeybees with TaqMan Quantitative Real-Time RT-PCR Assays. Viruses. 2021; 13(7):1340. https://doi.org/10.3390/v13071340
Chicago/Turabian StyleŠimenc, Laura, Tanja Knific, and Ivan Toplak. 2021. "The Comparison of Honeybee Viral Loads for Six Honeybee Viruses (ABPV, BQCV, CBPV, DWV, LSV3 and SBV) in Healthy and Clinically Affected Honeybees with TaqMan Quantitative Real-Time RT-PCR Assays" Viruses 13, no. 7: 1340. https://doi.org/10.3390/v13071340
APA StyleŠimenc, L., Knific, T., & Toplak, I. (2021). The Comparison of Honeybee Viral Loads for Six Honeybee Viruses (ABPV, BQCV, CBPV, DWV, LSV3 and SBV) in Healthy and Clinically Affected Honeybees with TaqMan Quantitative Real-Time RT-PCR Assays. Viruses, 13(7), 1340. https://doi.org/10.3390/v13071340