
Modeling Abundance of Culicoides stellifer, a Candidate Orbivirus Vector, Indicates Nonrandom Hemorrhagic Disease Risk for White-Tailed Deer (Odocoileus virginianus)

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
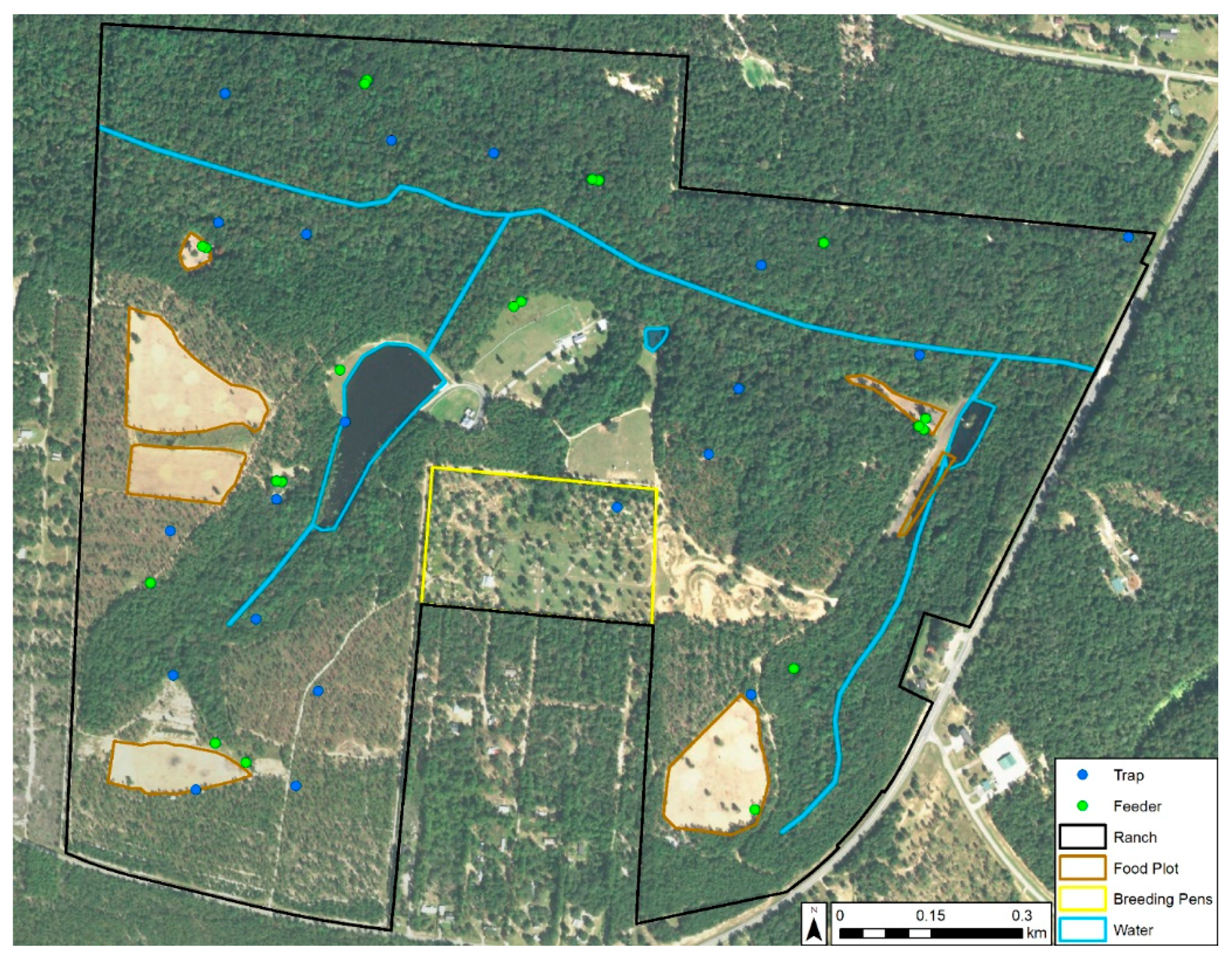
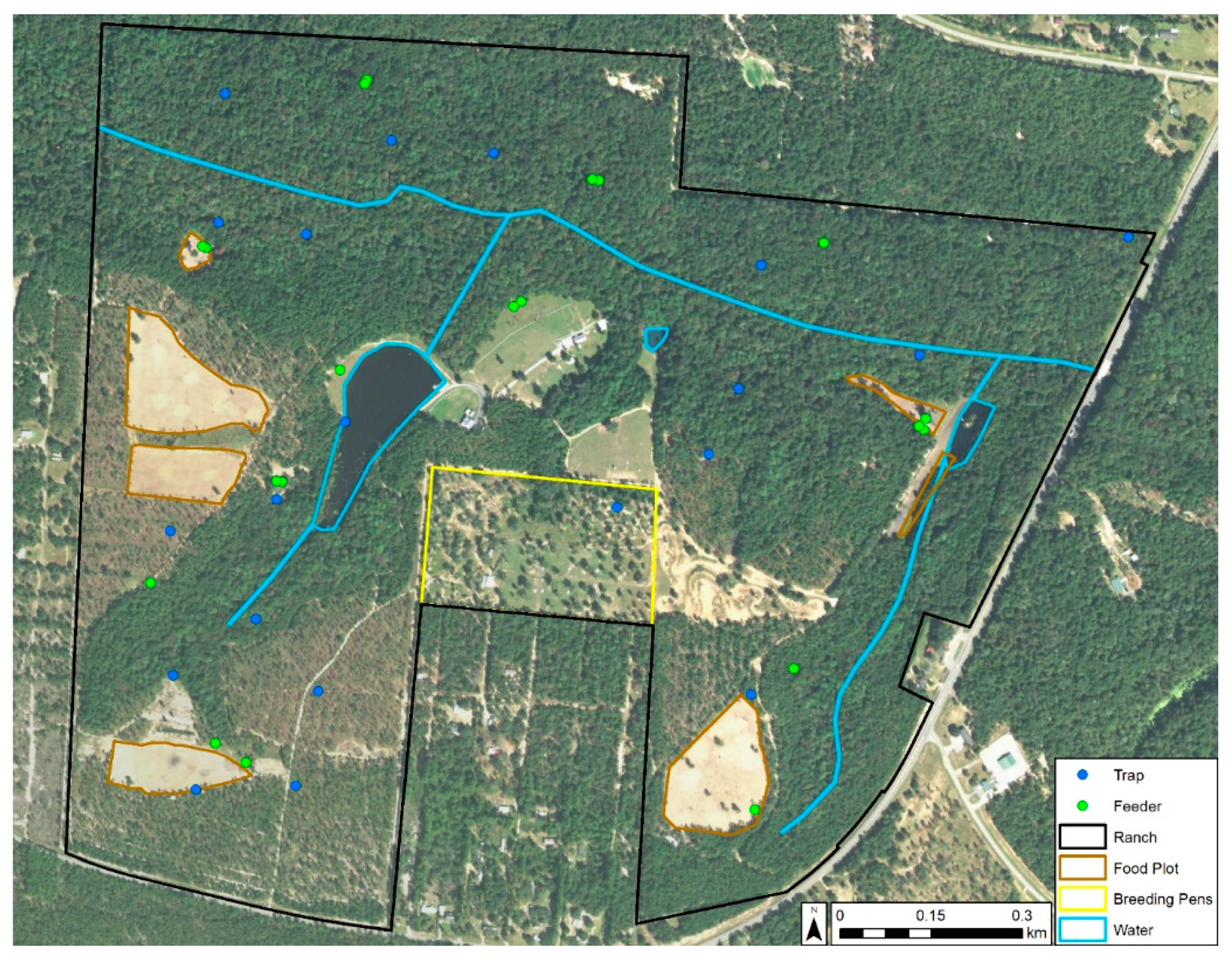
2.1. Study Area
Entomological Sampling
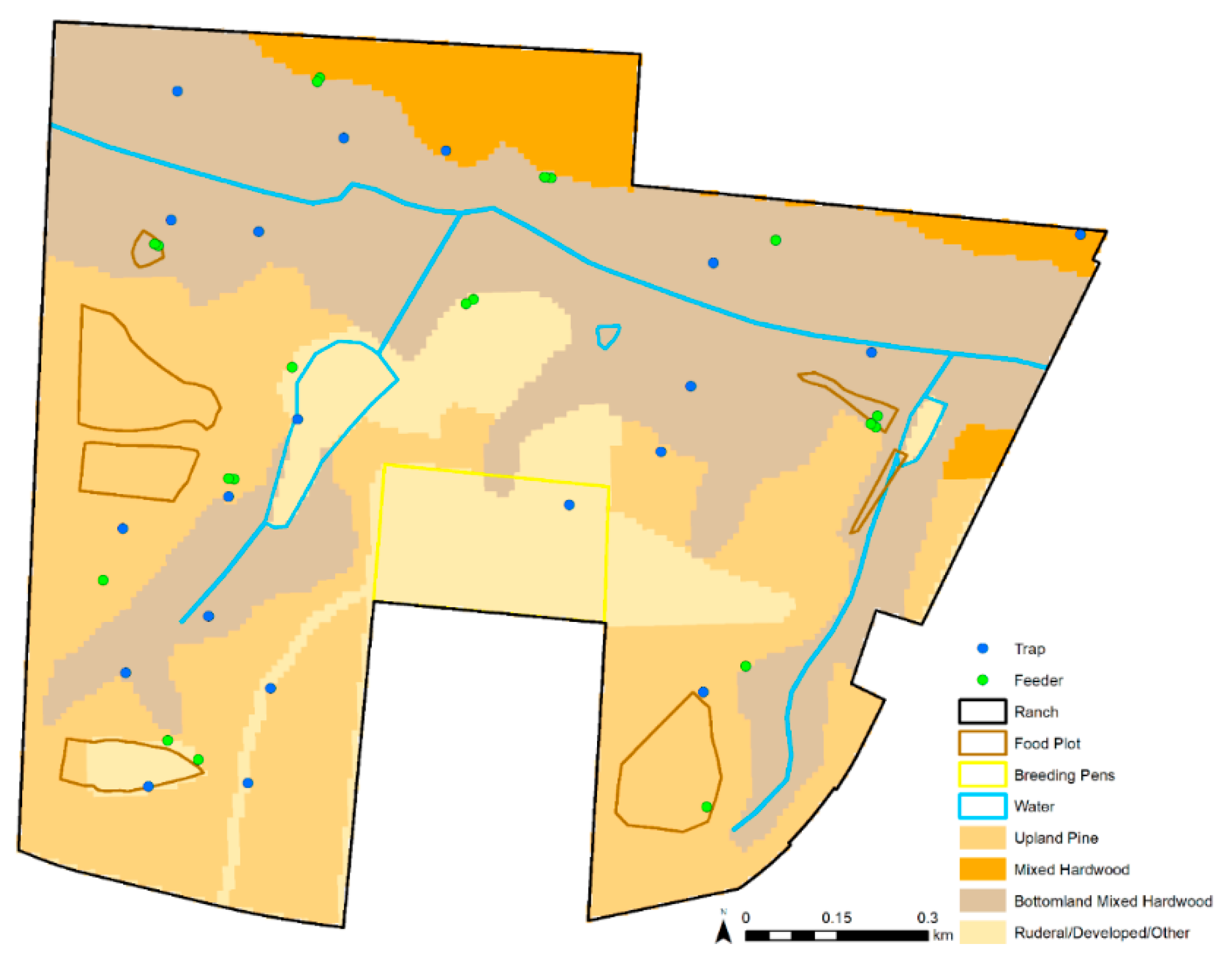
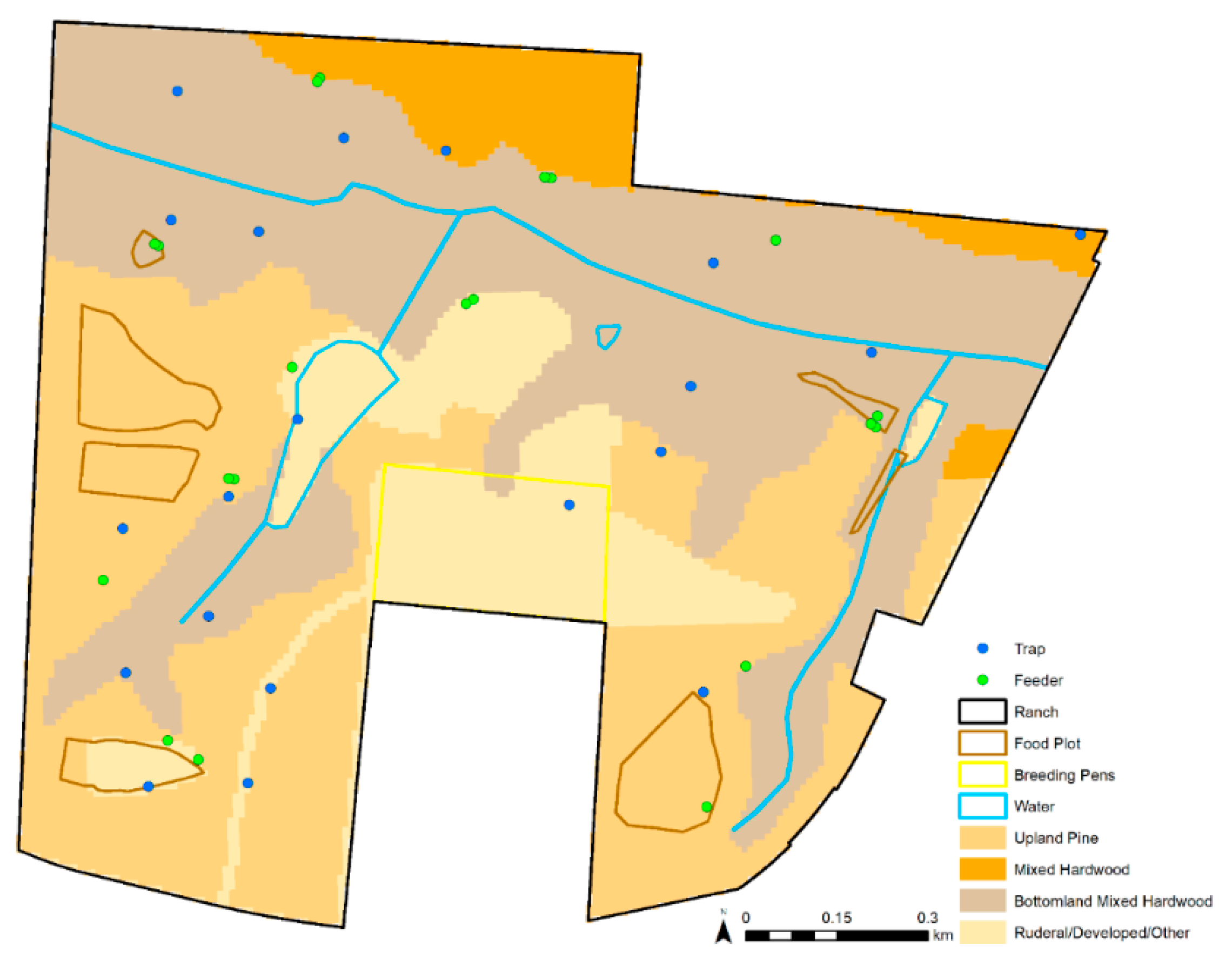
2.2. Environmental Data
2.3. Model Construction
3. Results
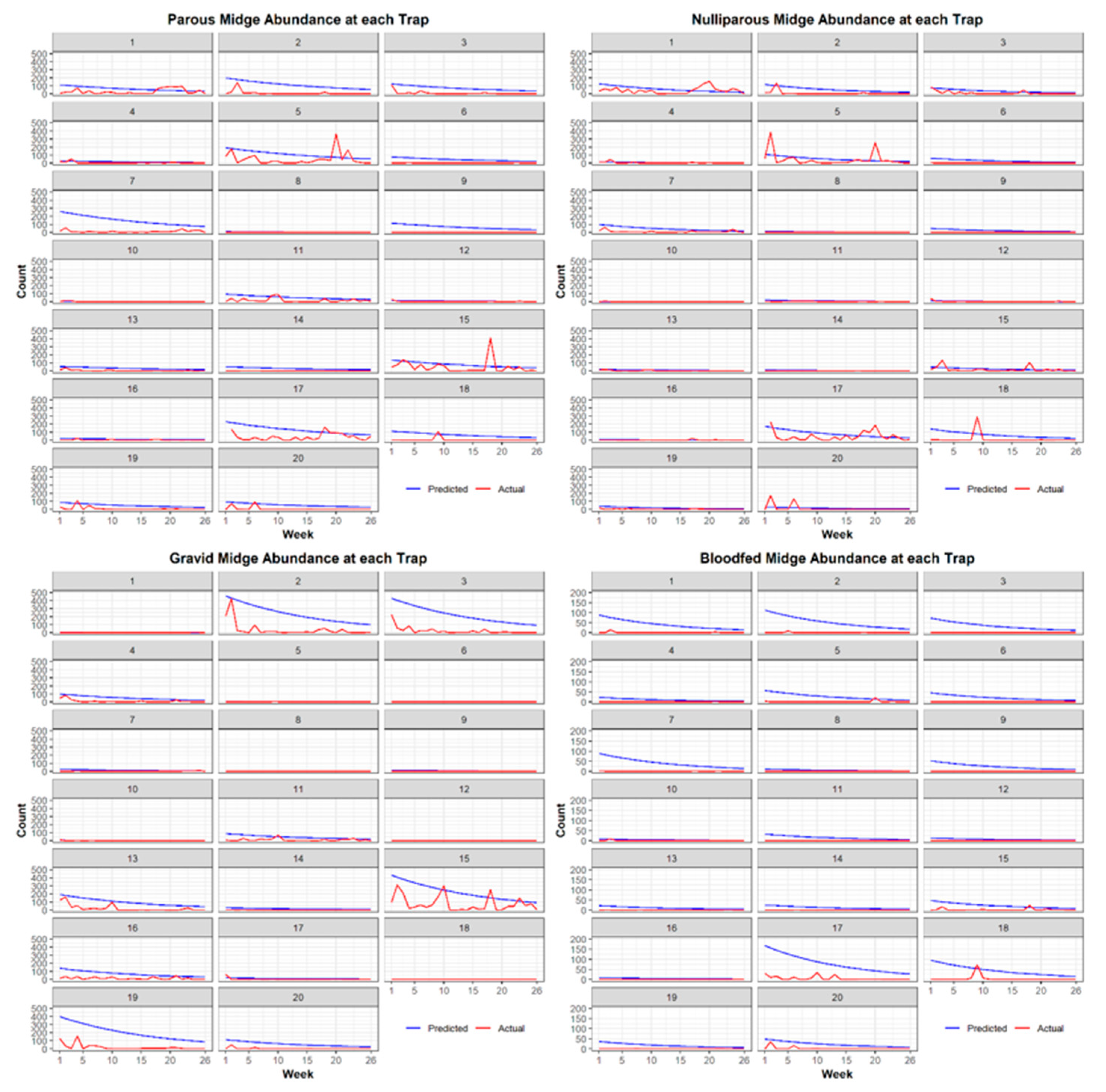
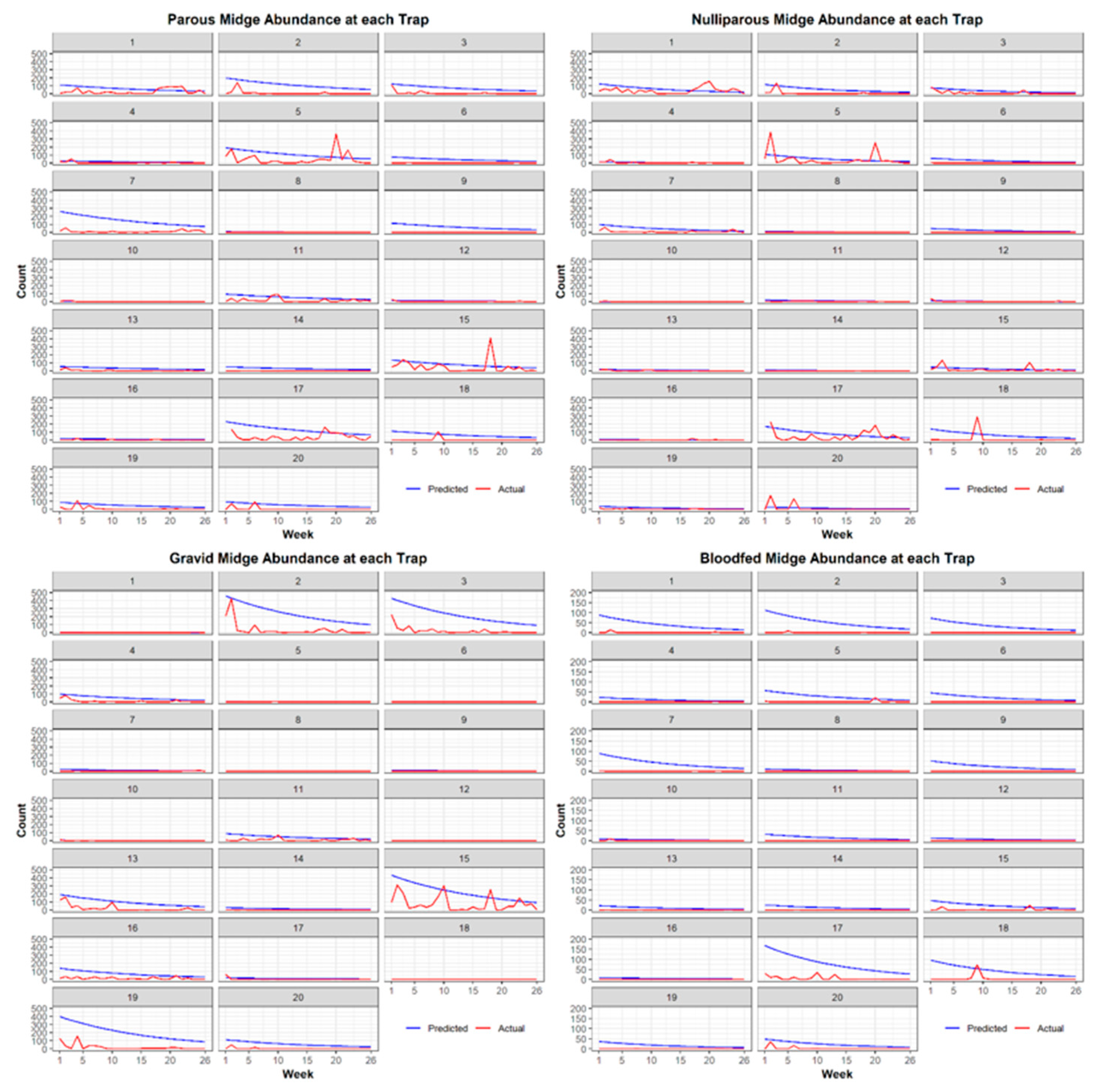
3.1. Entomological Sampling
Best Models for Each Physiological Stage of C. stellifer
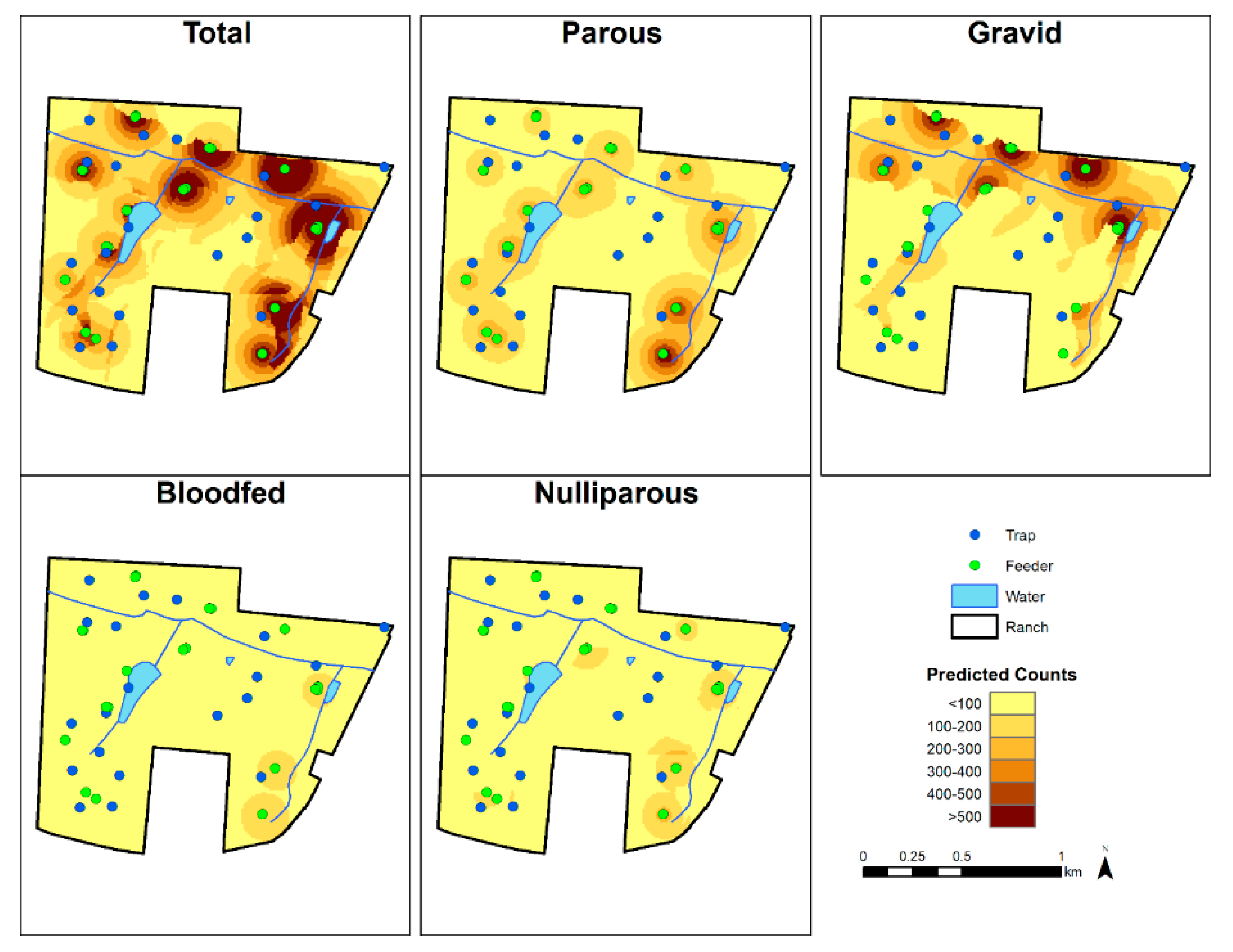
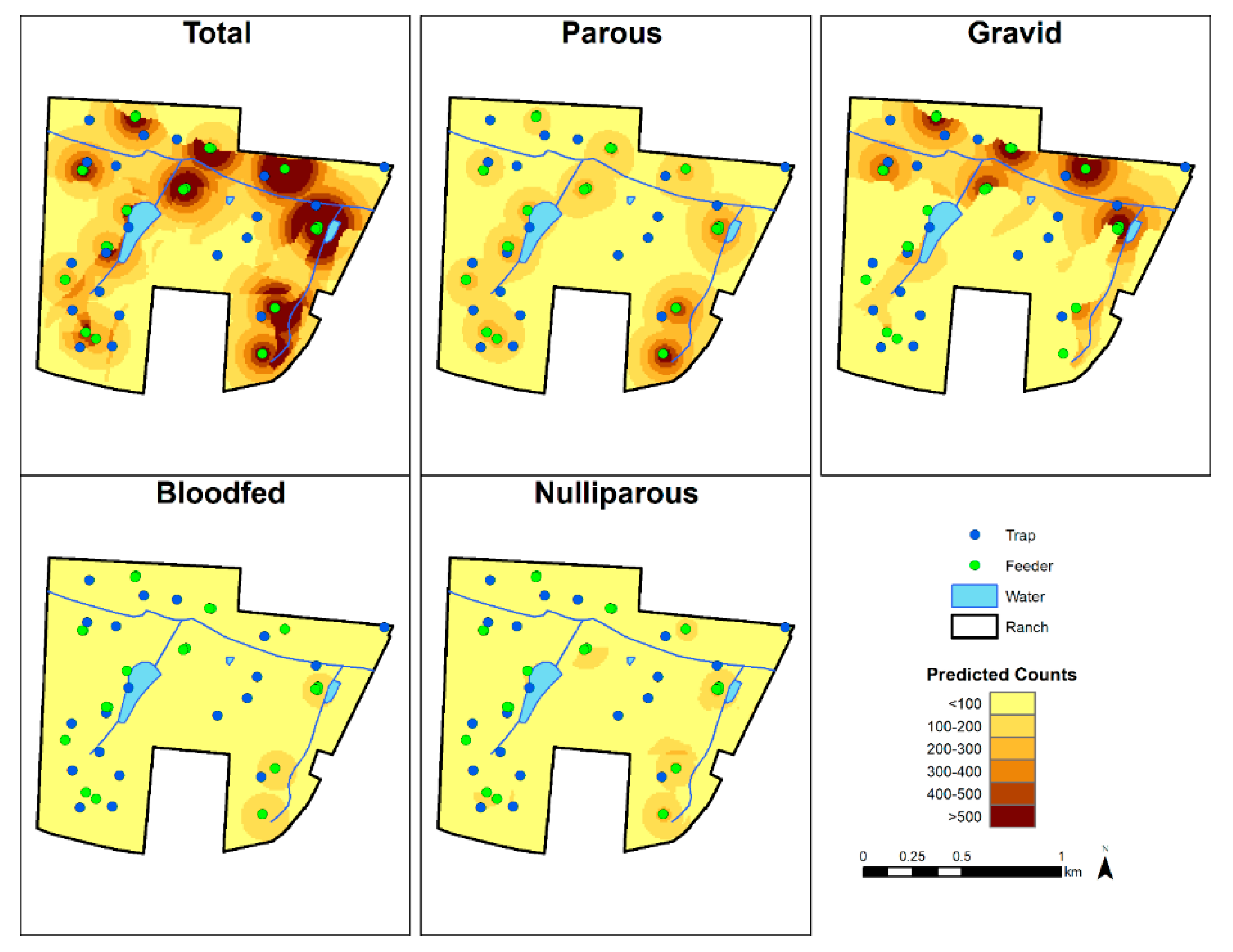
3.2. Spatial Predictions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, D.P.; Outlaw, J.L.; Earle, M.; Richardson, J.W. Economic Impact of U.S. Deer Breeding and Hunting Operations; Agricultural and Food Policy Center, Texas A&M University: College Station, TX, USA, 2017. [Google Scholar]
- Jones, R.H.; Roughton, R.D.; Foster, N.M.; Bando, B.M. Culicoides, the Vector of Epizootic Hemorrhagic Disease in White-Tailed Deer in Kentucky in 1971. J. Wildl. Dis. 1977, 13, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P.J. Transmission and Epidemiology of Bluetongue and Epizootic Hemorrhagic Disease in North America: Current Perspectives, Research Gaps, and Future Directions. Vector-Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Blanton, F.S.; Wirth, W.W. The sand flies (Culicoides) of Florida (Diptera: Ceratopogonidae). In Arthropods of Florida and Neighboring Land Areas; Florida Department of Agriculture and Consumer Affairs, Division of Plant Industry: Gainesville, FL, USA, 1979; Volume 10. [Google Scholar]
- Pfannenstiel, R.S.; Mullens, B.A.; Ruder, M.G.; Zurek, L.; Cohnstaedt, L.W.; Nayduch, D. Management of North American Culicoides Biting Midges: Current Knowledge and Research Needs. Vector-Borne Zoonotic Dis. 2015, 15, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Mellor, P.; Torr, S. Control Techniques for Culicoides Biting Midges and Their Application in the UK and Northwestern Palaearctic. Med. Vet. Entomol. 2008, 22, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Harrup, L.E.; Miranda, M.A.; Carpenter, S. Advances in Control Techniques for Culicoides and Future Prospects. Vet. Ital. 2016, 52, 247–264. [Google Scholar] [PubMed]
- McGregor, B.; Stenn, T.; Sayler, K.; Blosser, E.; Blackburn, J.; Wisely, S.; Burkett-Cadena, N. Host Use Patterns of Culicoides Spp. Biting Midges at a Big Game Preserve in Florida, USA, and Implications for the Transmission of Orbiviruses. Med. Vet. Entomol. 2019, 33, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisen, W.K. Epidemiology of vector-borne diseases. In Medical Veterinary Entomology; Academic Press: San Diego, CA, USA, 2002; pp. 15–29. ISBN 978-0-12-510451-7. [Google Scholar]
- Royle, J.A. N-Mixture Models for Estimating Population Size from Spatially Replicated Counts. Biometrics 2004, 60, 108–115. [Google Scholar] [CrossRef]
- Abad-Franch, F.; Ferraz, G.; Campos, C.; Palomeque, F.S.; Grijalva, M.J.; Aguilar, H.M.; Miles, M.A. Modeling Disease Vector Occurrence When Detection Is Imperfect: Infestation of Amazonian Palm Trees by Triatomine Bugs at Three Spatial Scales. PLOS Negl. Trop. Dis. 2010, 4, e620. [Google Scholar] [CrossRef] [Green Version]
- Drolet, B.S.; van Rijn, P.; Howerth, E.W.; Beer, M.; Mertens, P.P. A Review of Knowledge Gaps and Tools for Orbivirus Research. Vector Borne Zoonotic Dis. 2015, 15, 339–347. [Google Scholar] [CrossRef]
- Padilla-Torres, S.D.; Ferraz, G.; Luz, S.L.B.; Zamora-Perea, E.; Abad-Franch, F. Modeling Dengue Vector Dynamics under Imperfect Detection: Three Years of Site-Occupancy by Aedes Aegypti and Aedes Albopictus in Urban Amazonia. PLoS ONE 2013, 8, e58420. [Google Scholar] [CrossRef]
- Cauvin, A.; Dinh, E.T.N.; Orange, J.P.; Shuman, R.M.; Blackburn, J.K.; Wisely, S.M. Antibodies to Epizootic Hemorrhagic Disease Virus (EHDV) in Adjacent Farmed and Wild Florida White-Tailed Deer (Odocoileus virginianus). J. Wildl. Dis. 2020, 56, 208–213. [Google Scholar] [CrossRef]
- ESRI. ArcGIS 10.3.1 for Desktop, ESRI: Redlands, CA, USA, 2014.
- Erram, D.; Burkett-Cadena, N. Laboratory Studies on the Oviposition Stimuli of Culicoides Stellifer (Diptera: Ceratopogonidae), a Suspected Vector of Orbiviruses in the United States. Parasites Vectors 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Erram, D.; Blosser, E.M.; Burkett-Cadena, N. Habitat Associations of Culicoides Species (Diptera: Ceratopogonidae) Abundant on a Commercial Cervid Farm in Florida, USA. Parasites Vectors 2019, 12, 367. [Google Scholar] [CrossRef] [Green Version]
- Natural Resources Conservation Service National Hydrography Dataset 1:24,000. 2017. Available online: https://www.usgs.gov/core-science-systems/ngp/national-hydrography/national-hydrography-dataset?qt-science_support_page_related_con=0#qt-science_support_page_related_con (accessed on 15 August 2019).
- Florida Fish and Wildlife Conservation Commission Tallahassee Florida. Florida Natural Areas Inventory Florida Cooperative Land Cover Map; Version 3.1. 2019. Available online: https://myfwc.com/research/gis/regional-projects/cooperative-land-cover/ (accessed on 15 August 2019).
- Dinh, E.T.N. Epizootic Hemorrhagic Disease Virus Transmission in North Florida Free-Ranging Ranched and Wild White-Tailed Deer. Ph.D. Dissertation, University of Florida, Gainesville, FL, USA, 2019. [Google Scholar]
- Worton, B.J. Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Getz, W.M.; Wilmers, C.C. A Local Nearest-Neighbor Convex-Hull Construction of Home Ranges and Utilization Distributions. Ecography 2004, 27, 489–505. [Google Scholar] [CrossRef] [Green Version]
- Calenge, C. Home Range Estimation in R: The AdehabitatHR Package; CRAN: Auffargis, France, 2015. [Google Scholar]
- Kennedy, C.M.; Grant, E.H.C.; Neel, M.C.; Fagan, W.F.; Marra, P.P. Landscape Matrix Mediates Occupancy Dynamics of Neotropical Avian Insectivores. Ecol. Appl. 2011, 21, 1837–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazerolle, M.J.; Desrochers, A.; Rochefort, L. Landscape Characteristics Influence Pond Occupancy by Frogs after Accounting for Detectability. Ecol. Appl. 2005, 15, 824–834. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.E.; Swihart, R.K. Modeling Patch Occupancy by Forest Rodents: Incorporating Detectability and Spatial Autocorrelation with Hierarchically Structured Data. J. Wildl. Manag. 2005, 69, 933–949. [Google Scholar] [CrossRef]
- Fiske, I.; Chandler, R. Overview of Unmarked: An R Package for the Analysis of Data from Unmarked Animals; CRAN: Auffargis, France, 2017. [Google Scholar]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43. [Google Scholar] [CrossRef] [Green Version]
- Fiske, I.; Chandler, R.; Miller, D.; Royle, A.; Kery, M.; Hostetler, J.; Hutchinson, R. Package ‘Unmarked’; CRAN: Auffargis, France, 2017. [Google Scholar]
- Sileshi, G. A Method for Estimating Insect Abundance and Patch Occupancy with Potential Applications in Large-Scale Monitoring Programmes. Afr. Entomol. 2007, 15, 89–101. [Google Scholar] [CrossRef]
- Manly, B.F.J.; McDonald, L.L.; Thomas, D.L.; McDonald, T.L.; Erickson, W. Resource Selection by Animals: Statistical Design and Analysis for Field Studies, 2nd ed.; Springer: Dordrecht, The Netherlands, 2002; ISBN 978-1-4020-0677-7. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Thompson, W.L.; Lee, D.C. Modeling Relationships between Landscape-Level Attributes and Snorkel Counts of Chinook Salmon and Steelhead Parr in Idaho. Can. J. Fish. Aquat. Sci. 2000, 57, 1834–1842. [Google Scholar] [CrossRef]
- Miller, R.; Kaneene, J.B.; Fitzgerald, S.D.; Schmitt, S.M. Evaluation of the Influence of Supplemental Feeding of White-Tailed Deer (Odocoileus virginianus) on the Prevalence of Bovine Tuberculosis in the Michigan Wild Deer Population. J. Wildl. Dis. 2003, 39, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Cross, P.C.; Edwards, W.H.; Scurlock, B.M.; Maichak, E.J.; Rogerson, J.D. Effects of Management and Climate on Elk Brucellosis in the Greater Yellowstone Ecosystem. Ecol. Appl. 2007, 17, 957–964. [Google Scholar] [CrossRef]
- Yaremych, S.A.; Novak, R.J.; Raim, A.J.; Mankin, P.C.; Warner, R.E. Home Range and Habitat Use by American Crows in Relation to Transmission of West Nile Virus. Wilson Bull. 2004, 116, 232–239. [Google Scholar] [CrossRef]
- Rustand, M. Home-Range Fidelity and the Effect of Supplemental Feeding on Contact Rates between White-Tailed Deer (Odocoileus virginianus) in Southern Illinois. Master’s Thesis, Southern Illinois University at Carbondale, Ann Arbor, MI, USA, 2010. [Google Scholar]
- Brown, L.M.; Hall, R.J. Consequences of Resource Supplementation for Disease Risk in a Partially Migratory Population. Philos. Trans. R. Soc. B 2018, 373, 20170095. [Google Scholar] [CrossRef] [Green Version]
- Calvo, J.H.; Berzal, B.; Calvete, C.; Miranda, M.A.; Estrada, R.; Lucientes, J. Host Feeding Patterns of Culicoides Species (Diptera: Ceratopogonidae) within the Picos de Europa National Park in Northern Spain. Bull. Entomol. Res. 2012, 102, 692–697. [Google Scholar] [CrossRef]
- McGregor, B.L.; Sloyer, K.E.; Sayler, K.A.; Goodfriend, O.; Krauer, J.M.C.; Acevedo, C.; Zhang, X.; Mathias, D.; Wisely, S.M.; Burkett-Cadena, N.D. Field Data Implicating Culicoides Stellifer and Culicoides Venustus (Diptera: Ceratopogonidae) as Vectors of Epizootic Hemorrhagic Disease Virus. Parasites Vectors 2019, 12, 258. [Google Scholar] [CrossRef]
- Orange, J.; Dinh, E.; Shuman, R.; Wisely, S.M.; Blackburn, J.K.; Blackburn, J.K. Inter-Annual Site Fidelity of Wild and Farmed White-Tailed Deer in Florida: Implications for Disease Intervention. Vector-Borne Zoonotic Dis. 2020. submitted. [Google Scholar]
- Vanderhoof, R.; Jacobson, H. Production and Use of Agricultural Food Planting by Deer on Marion County Wildlife Management Area, Mississippi Completion Report: 1984–1989; Mississippi Department of Wildlife, Fisheries and Parks: Jackson, MS, USA, 1990. [Google Scholar]
- Vanderhoof, R.E.; Jacobson, H.A. Food Plots and Deer Home Range Movements in the Southern Coastal Plain. In Proceedings of the Annual Conference of the Southeastern Association of Fish and Wildlife Agencies, Atlanta, GA, USA, 9–13 October 1993; Volume 47, pp. 33–42. [Google Scholar]
- Garver, L.W. Effects of Supplemental Feed on Foraging Behavior and Activity of White-Tailed Deer. Master’s Thesis, Texas A&M University, Kingsville, TX, USA, 2011. [Google Scholar]
- Wiemers, D.W.; Fulbright, T.E.; Wester, D.B.; Ortega-S, J.A.; Rasmussen, G.A.; Hewitt, D.G.; Hellickson, M.W. Role of Thermal Environment in Habitat Selection by Male White-Tailed Deer during Summer in Texas, USA. Wildl. Biol. 2014, 20, 47–56. [Google Scholar] [CrossRef]
- Zimmerman, R.H.; Turner, E.C., Jr. Host-Feeding Patterns of Culicoides (Diptera: Ceratopogonidae) Collected from Livestock in Virginia, USA. J. Med. Entomol. 1983, 20, 514–519. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description |
|---|---|
| Feeder | Euclidian distance from each feeder |
| Water | Euclidian distance from major water bodies |
| Habitat | Discrete habitat category: upland pine, mixed hardwood pine, mixed bottomland hardwood, or rural/developed/other |
| Utilization distribution (UD) | Weekly probability density of deer presence |
| Latitude | Insect trap location latitude |
| Longitude | Insect trap location longitude |
| Week | Week in which samples were collected |
| Latitude | Longitude | Feeder | Water | Weekly UD | |
|---|---|---|---|---|---|
| Latitude | 1.0000 | 0.2570 | 0.5661 | −0.0282 | −0.0789 |
| Longitude | 0.2570 | 1.0000 | 0.2170 | −0.5426 | −0.0789 |
| Feeder | 0.5661 | 0.2170 | 1.0000 | 0.3105 | −0.0451 |
| Water | −0.0282 | −0.5426 | 0.3105 | 1.0000 | −0.0331 |
| Weekly UD | −0.0789 | −0.2556 | −0.0451 | −0.0331 | 1.0000 |
| Variables | p |
|---|---|
| Feeder and habitat | <2.0 × 10−16 |
| Water and habitat | <2.0 × 10−16 |
| UD and habitat | 0.0002 |
| Status | Variable | Estimate | SE | p-Value |
|---|---|---|---|---|
| All | Intercept | 4.8848 | 0.2365 | 8.11 × 10−95 |
| Week | −0.0569 | 0.0090 | 3.00 × 10−10 | |
| Latitude | 0.3853 | 0.1088 | 3.99 × 10−4 | |
| Longitude | −0.0207 | 0.0910 | 0.8200 | |
| Mixed hardwood pine | −0.5807 | 0.3770 | 0.1230 | |
| Mixed bottomland hardwood | 0.6705 | 0.2476 | 0.0068 | |
| Rural/developed/other | 0.4843 | 0.2452 | 0.0482 | |
| Feeder | −0.3005 | 0.0962 | 5.29 × 10−25 | |
| Parous | Intercept | 4.2716 | 0.1666 | 6.37 × 10−145 |
| Week | −0.0507 | 0.0106 | 1.72 × 10−6 | |
| Latitude | 0.1608 | 0.1095 | 0.1420 | |
| Longitude | −0.3355 | 0.1046 | 0.0013 | |
| Feeder | −0.9647 | 0.1188 | 4.59 × 10−16 | |
| Water | −0.3005 | 0.0962 | 0.0018 | |
| Gravid | Intercept | 2.5957 | 0.2455 | 3.94 × 10−26 |
| Week | −0.0613 | 0.0095 | 9.62 × 10−11 | |
| Latitude | 0.2636 | 0.0997 | 0.0082 | |
| Longitude | 0.3327 | 0.1067 | 0.0018 | |
| Mixed hardwood pine | 0.3859 | 0.4426 | 0.3830 | |
| Mixed bottomland hardwood | 2.5626 | 0.2541 | 3.66 × 10−24 | |
| Rural/developed/other | −1.0642 | 0.2944 | 0.0003 | |
| Feeder | −0.8350 | 0.10922 | 2.08 × 10−14 | |
| Bloodfed | Intercept | 3.7417 | 0.3205 | 1.76 × 10−31 |
| Week | −0.0721 | 0.0191 | 0.0002 | |
| Latitude | 0.4233 | 0.1777 | 0.0172 | |
| Longitude | −0.3422 | 0.1377 | 0.0130 | |
| Feeder | −0.8664 | 0.1795 | 1.38 × 10−6 | |
| Nulliparous | Intercept | 3.7178 | 0.2757 | 1.93 × 10−41 |
| Week | −0.0714 | 0.0108 | 3.69 × 10−11 | |
| Latitude | 0.5067 | 0.1268 | 6.43 × 10−5 | |
| Longitude | −0.1940 | 0.1072 | 0.0703 | |
| Mixed hardwood pine | −1.1957 | 0.4497 | 0.0078 | |
| Mixed bottomland hardwood | −0.2292 | 0.2918 | 0.4320 | |
| Rural/developed/other | 0.5516 | 0.2857 | 0.0535 | |
| Feeder | −1.0703 | 0.1176 | 9.08 × 10−20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinh, E.T.N.; Gomez, J.P.; Orange, J.P.; Morris, M.A.; Sayler, K.A.; McGregor, B.L.; Blosser, E.M.; Burkett-Cadena, N.D.; Wisely, S.M.; Blackburn, J.K. Modeling Abundance of Culicoides stellifer, a Candidate Orbivirus Vector, Indicates Nonrandom Hemorrhagic Disease Risk for White-Tailed Deer (Odocoileus virginianus). Viruses 2021, 13, 1328. https://doi.org/10.3390/v13071328
Dinh ETN, Gomez JP, Orange JP, Morris MA, Sayler KA, McGregor BL, Blosser EM, Burkett-Cadena ND, Wisely SM, Blackburn JK. Modeling Abundance of Culicoides stellifer, a Candidate Orbivirus Vector, Indicates Nonrandom Hemorrhagic Disease Risk for White-Tailed Deer (Odocoileus virginianus). Viruses. 2021; 13(7):1328. https://doi.org/10.3390/v13071328
Chicago/Turabian StyleDinh, Emily T. N., Juan Pablo Gomez, Jeremy P. Orange, Max A. Morris, Katherine A. Sayler, Bethany L. McGregor, Erik M. Blosser, Nathan D. Burkett-Cadena, Samantha M. Wisely, and Jason K. Blackburn. 2021. "Modeling Abundance of Culicoides stellifer, a Candidate Orbivirus Vector, Indicates Nonrandom Hemorrhagic Disease Risk for White-Tailed Deer (Odocoileus virginianus)" Viruses 13, no. 7: 1328. https://doi.org/10.3390/v13071328
APA StyleDinh, E. T. N., Gomez, J. P., Orange, J. P., Morris, M. A., Sayler, K. A., McGregor, B. L., Blosser, E. M., Burkett-Cadena, N. D., Wisely, S. M., & Blackburn, J. K. (2021). Modeling Abundance of Culicoides stellifer, a Candidate Orbivirus Vector, Indicates Nonrandom Hemorrhagic Disease Risk for White-Tailed Deer (Odocoileus virginianus). Viruses, 13(7), 1328. https://doi.org/10.3390/v13071328