
Infectivity of an Infectious Clone of Banana Streak CA Virus in A-Genome Bananas (Musa acuminata ssp.)

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Rolling Circle Amplification
2.3. PCR Detection of BSCAV
2.4. Cloning and Sanger Sequencing of the BSCAV-Brisbane Genomic Sequence
2.5. Construction of the Infectious Clone
2.6. Plant Inoculation and Screening for Virus Infection
2.7. Transmission Electron Microscopy
3. Results and Discussion
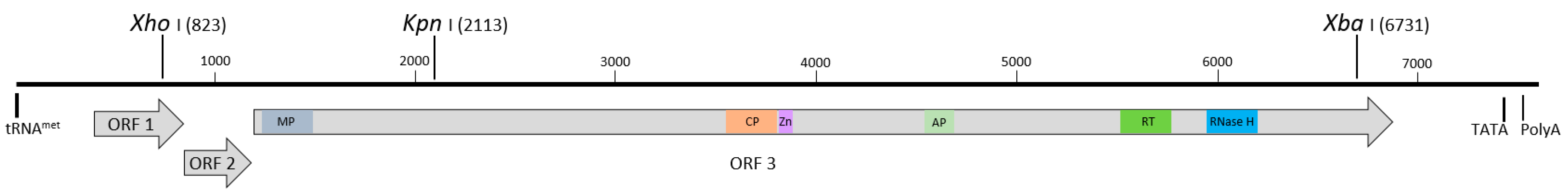
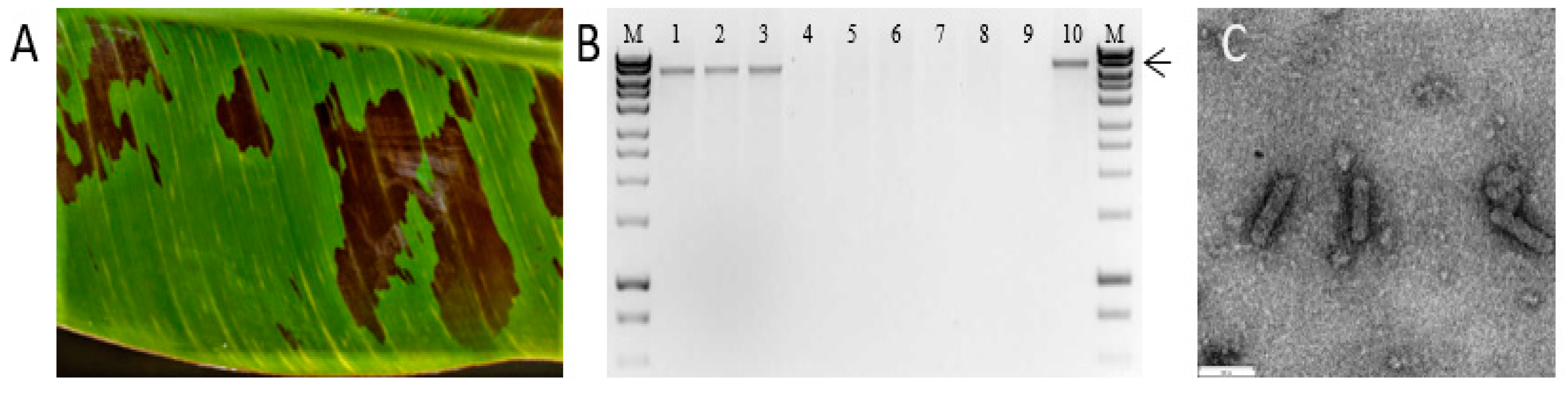
3.1. Identification of BSCAV and Characterization of the Full-Length Sequence
3.2. Development of an Infectious Clone and Assessment of Infectivity in M. Acuminata Bananas
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org/taxonomy/w/ictv-taxonomy (accessed on 12 August 2020).
- Iskra-Caruana, M.L.; Thomas, J.E.; Chabannes, M. Banana Streak. In Handbook of Diseases of Banana, Abacá and Enset; Jones, D.R., Ed.; CABI: Boston, MA, USA, 2018; pp. 393–409. ISBN 9781780647197. [Google Scholar]
- Bhat, A.I.; Hohn, T.; Selvarajan, R. Badnaviruses: The Current Global Scenario. Viruses 2016, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Geering, A.D. Caulimoviridae (Plant Pararetroviruses). In eLS; John Wiley & Sons, Ltd: Chichester, UK, 2014; ISBN 9780470015902. [Google Scholar] [CrossRef]
- Medberry, S.L.; Lockhart, B.E.L.; Olszewski, N.E. Properties of Commelina yellow mottle virus’s complete DNA sequence, genomic discontinuities and transcript suggest that it is a pararetrovirus. Nucleic Acids Res. 1990, 18, 5505–5513. [Google Scholar] [CrossRef] [PubMed]
- Bouhida, M.; Lockhart, B.E.L.; Olszewski, N.E. An analysis of the complete sequence of a sugarcane bacilliform virus genome infectious to banana and rice. J. Gen. Virol. 1993, 74, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, E.; Hagen, L.S.; Michler, P.; Rohfritsch, O.; Stussi-Garaud, C.; Keller, M.; Jacquemond, M.; Yot, P. In situ localization of cacao swollen shoot virus in agroinfected Theobroma cacao. Arch. Virol. 1999, 144, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Hartung, J.S. Cloning and sequence analysis of an infectious clone of Citrus yellow mosaic virus that can infect sweet orange via Agrobacterium-mediated inoculation. J. Gen. Virol. 2001, 82, 2549–2558. [Google Scholar] [CrossRef] [PubMed]
- Kidanemariam, D.B.; Sukal, A.C.; Crew, K.; Jackson, G.V.H.; Abraham, A.D.; Dale, J.L.; Harding, R.M.; James, A.P. Characterization of an Australian isolate of taro bacilliform virus and development of an infectious clone. Arch. Virol. 2018, 163, 1677–1681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.; Han, R. Advances in the Application of Virus-Induced Gene Silencing in Plants. Am. J. Plant Sci. 2019, 10, 1649–1661. [Google Scholar] [CrossRef]
- Abrahamian, P.; Hammond, R.W.; Hammond, J. Plant Virus–Derived Vectors: Applications in Agricultural and Medical Biotechnology. Annu. Rev. Virol. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- D’Hont, A.; Denoeud, F.; Aury, J.-M.; Baurens, F.-C.; Carreel, F.; Garsmeur, O.; Noel, B.; Bocs, S.; Droc, G.; Rouard, M.; et al. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature 2012, 488, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Perrier, X.; Langhe, E.D.; Donohue, M.; Lentfer, C.; Vrydaghs, L.; Bakry, F.; Carreel, F.; Hippolyte, I.; Horry, J.-P.; Jenny, C.; et al. Multidisciplinary perspectives on banana (Musa spp.) domestication. Proc. Nalt. Acad. Sci. USA 2011, 108, 11311–11318. [Google Scholar] [CrossRef] [PubMed]
- Onsarigo, M.N. Assessment of Banana Streak MY Virus-Based Infectious Clone Vectors in Musa ssp. Ph.D. Thesis, Queensland University of Technology, Brisbane, QLD, Australia, 2017. [Google Scholar]
- James, A.P.; Geijskes, R.J.; Dale, J.L.; Harding, R.M. Development of a Novel Rolling-Circle Amplification Technique to Detect Banana streak virus that also Discriminates Between Integrated and Episomal Virus Sequences. Plant Dis. 2011, 95, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Geering, A.D.W.; McMichael, L.A.; Dietzgen, R.G.; Thomas, J.E. Genetic Diversity Among Banana streak virus Isolates from Australia. Phytopathology 2000, 90, 921–927. [Google Scholar] [CrossRef] [PubMed]
- James, A.P.; Geijskes, R.J.; Dale, J.L.; Harding, R.M. Molecular characterisation of six badnavirus species associated with leaf streak disease of banana in East Africa. Ann. Appl. Biol. 2011, 158, 346–353. [Google Scholar] [CrossRef]
- Das Bhowmik, S.S.; Cheng, A.Y.; Long, H.; Tan, G.Z.H.; Hoang, T.M.L.; Karbaschi, M.R.; Williams, B.; Higgins, T.J.V.; Mundree, S.G. Robust genetic transformation system to obtain non-chimeric transgenic chickpea. Front. Plant Sci. 2019, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Ieki, H.; Matsuda, I. PCR detection of Ti and Ri plasmids from phytopathogenic Agrobacterium strains. Appl. Environ. Microbiol. 1995, 61, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, B.E.L. Purification and serology of a bacilliform virus associated with banana streak disease. Phytopathology 1986, 76, 995–999. [Google Scholar] [CrossRef]
- Rajeswaran, R.; Seguin, J.; Chabannes, M.; Duroy, P.-O.; Laboureau, N.; Farinelli, L.; Iskra-Caruana, M.-L.; Pooggin, M.M. Evasion of short interfering RNA-directed antiviral silencing in Musa acuminata persistently infected with six distinct banana streak pararetroviruses. J. Virol. 2014, 88, 11516–11528. [Google Scholar] [CrossRef]
- Geering, A.D.W.; Pooggin, M.M.; Olszewski, N.E.; Lockhart, B.E.L.; Thomas, J.E. Characterisation of Banana streak Mysore virus and evidence that its DNA is integrated in the B genome of cultivated Musa. Arch. Virol. 2005, 150, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.C.; Hafner, G.J.; Revill, P.A.; Dale, J.L.; Harding, R.M. Sequence diversity of South Pacific isolates of Taro bacilliform virus and the development of a PCR-based diagnostic test. Arch. Virol. 2003, 148, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Muller, E.; Dupuy, V.; Blondin, L.; Bauffe, F.; Daugrois, J.-H.; Nathalie, L.; Iskra-Caruana, M.-L. High molecular variability of sugarcane bacilliform viruses in Guadeloupe implying the existence of at least three new species. Virus Res. 2011, 160, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Musa Genome Information Service (MGIS). Available online: http://www.promusa.org/MGIS (accessed on 12 August 2020).




{kind=link}
{kind=link}
{kind=link}
| Accession Name (Taxonomic Subgroup) 1 | No. of Plants Testing Positive by PCR at 8 Weeks Post-Inoculation | No. of Plants with Symptoms at 8 Weeks (Typical Symptoms) | No. of Plants with Symptoms at 28 Weeks (Typical Symptoms) |
|---|---|---|---|
| Williams (Musa AAA Cavendish subgroup) | 5/5 | 5/5 (mild yellow-green streaks) | 5/5 (mild yellow flecking or asymptomatic) |
| Gros Michel (Musa AAA Gros Michel subgroup) | 5/5 | 5/5 (mild yellow-green streaks) | 5/5 (mild yellow flecking or asymptomatic) |
| Malaccensis (M. acuminata AA subsp. malaccensis) | 5/5 | 5/5 (mild yellow-green streaks) | 5/5 (mild yellow flecking or asymptomatic) |
| Khae Phrae (M. acuminata AA subsp. siamea) | 4/5 | 4/5 (yellow/chlorotic streaks; cigar leaf necrosis) | 5/5 (cigar leaf necrosis; pseudostem splitting; plant death) |
| Truncata (M. acuminata AA subsp. truncata) | 5/5 | 5/5 (continuous yellow and yellow-brown streaks) | 5/5 (mild yellow-brown flecking or asymptomatic) |
| Zebrina (M. acuminata AA subsp. zebrina) | 5/5 | No symptoms | No symptoms |
| Paka (Musa AA subgroup) | 5/5 | 5/5 (mild yellow-green streaks) | 5/5 (yellow flecking and streaks) |
| Igisahira Gisanzwe (Musa AAA EAHB subgroup) 2 | 4/5 | 3/5 (yellow-brown chlorotic patches; white, yellow and brown streaks) | 5/5 (chlorosis/necrosis of the leaf lamina; pseudostem splitting) |
| Pisang Madu (Musa AA subgroup) | 5/5 | 5/5 (yellow/chlorotic flecking) | 5/5 (yellow flecking and streaks) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, A.P.; Kidanemariam, D.B.; Hamill, S.D.; Dale, J.L.; Harding, R.M. Infectivity of an Infectious Clone of Banana Streak CA Virus in A-Genome Bananas (Musa acuminata ssp.). Viruses 2021, 13, 1071. https://doi.org/10.3390/v13061071
James AP, Kidanemariam DB, Hamill SD, Dale JL, Harding RM. Infectivity of an Infectious Clone of Banana Streak CA Virus in A-Genome Bananas (Musa acuminata ssp.). Viruses. 2021; 13(6):1071. https://doi.org/10.3390/v13061071
Chicago/Turabian StyleJames, Anthony P., Dawit B. Kidanemariam, Sharon D. Hamill, James L. Dale, and Robert M. Harding. 2021. "Infectivity of an Infectious Clone of Banana Streak CA Virus in A-Genome Bananas (Musa acuminata ssp.)" Viruses 13, no. 6: 1071. https://doi.org/10.3390/v13061071
APA StyleJames, A. P., Kidanemariam, D. B., Hamill, S. D., Dale, J. L., & Harding, R. M. (2021). Infectivity of an Infectious Clone of Banana Streak CA Virus in A-Genome Bananas (Musa acuminata ssp.). Viruses, 13(6), 1071. https://doi.org/10.3390/v13061071