
Honeysuckle Aqueous Extracts Induced let-7a Suppress EV71 Replication and Pathogenesis In Vitro and In Vivo and Is Predicted to Inhibit SARS-CoV-2

 , , ,
, , ,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Honeysuckle Aqueous Extract Preparation and Analysis of Blood miRNA Expression Profiles in Human Participants and Mice after Ingestion
2.2. Prediction of Honeysuckle-Induced miRNA and Pathway Analysis
2.3. Cell Culture and Viruses
2.4. Plaque Assay
2.5. Western Blot Analysis
2.6. Reporter Plasmids and Luciferase Assay
2.7. RT-PCR and Real-Time PCR
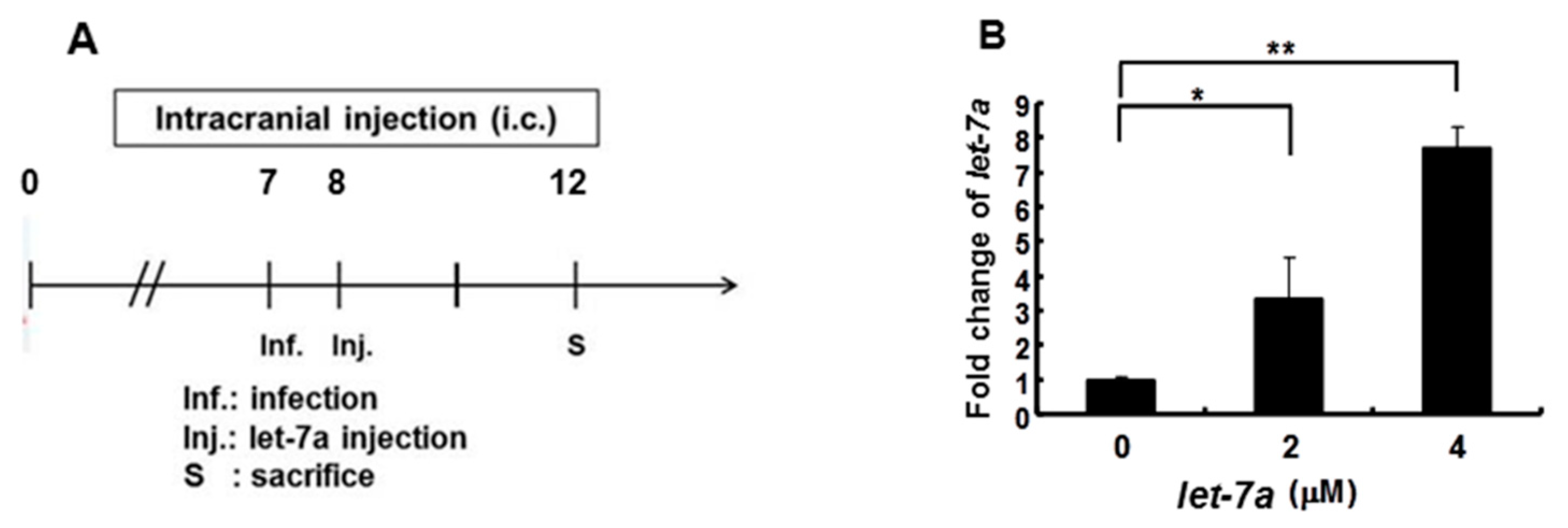
2.8. Suckling Mouse Model for EV71 Infection
3. Results
3.1. Honeysuckle-Induced Host miRNAs Both in Mice and Humans Were Predicted to Be Capable of Targeting Genome Sequences of SARS-CoV-2, DENV and EV71
3.2. In Silico Prediction of Honeysuckle-Related Pathways Are Highly Correlated to Diverse Virus Infections and Regulation of Cytokines
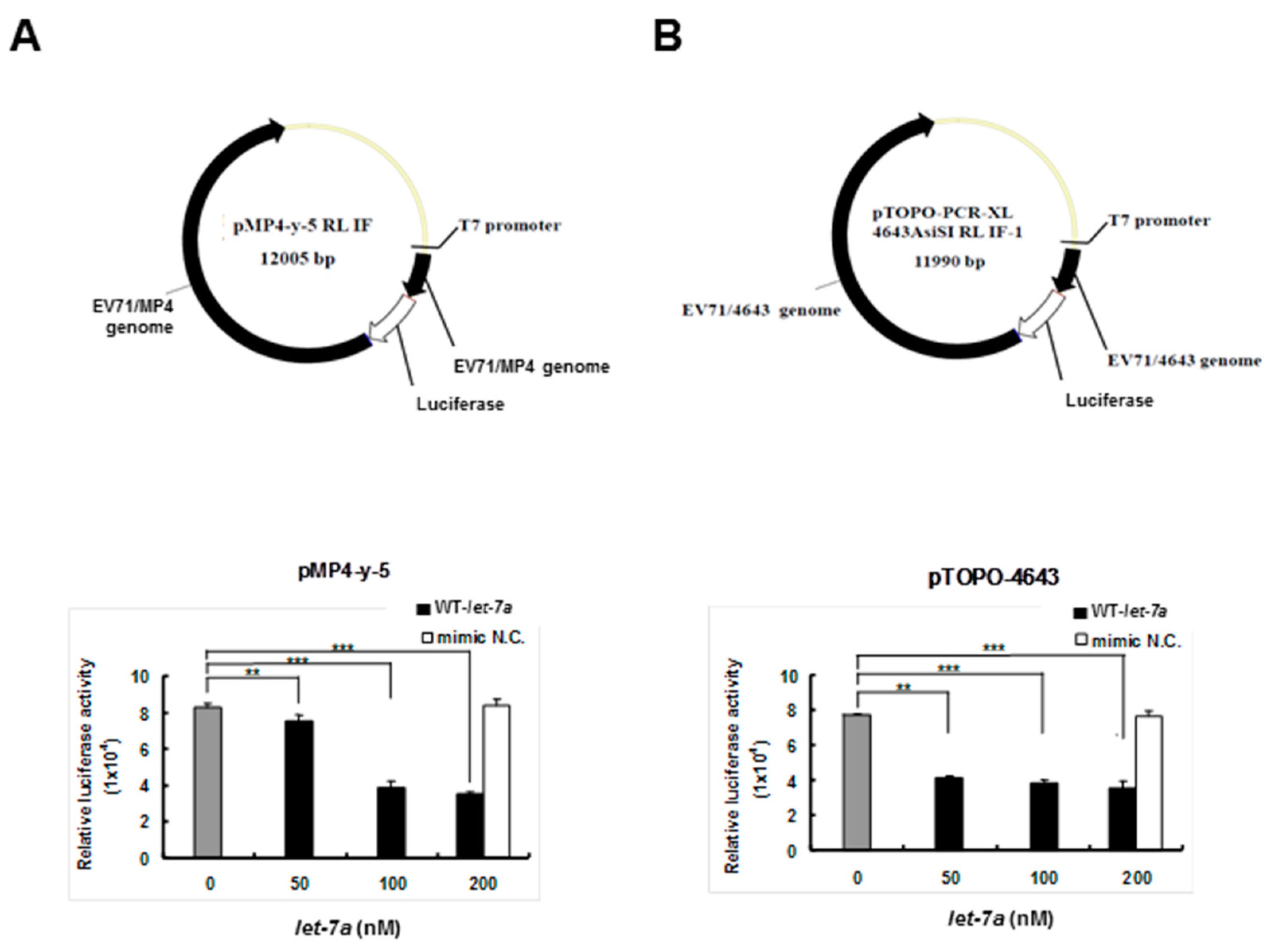
3.3. Let-7a Bound with Two EV71 Replicon Clones and Attenuated Luciferase Activity
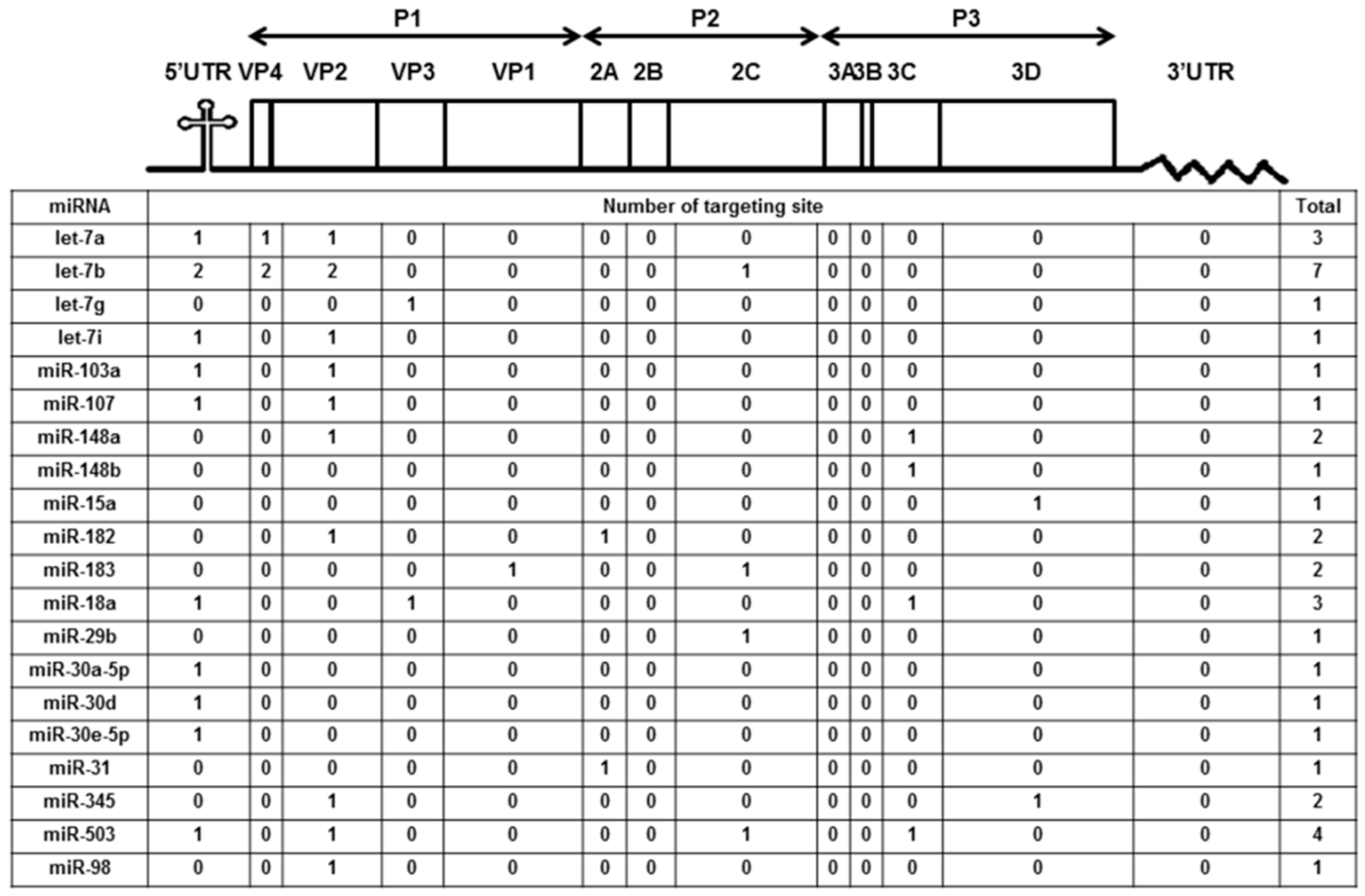
3.4. Let-7a Was Confirmed to Specifically Target the EV71 Genome at Two Regions, 5′UTR 233–254 and VP2 1234–1256
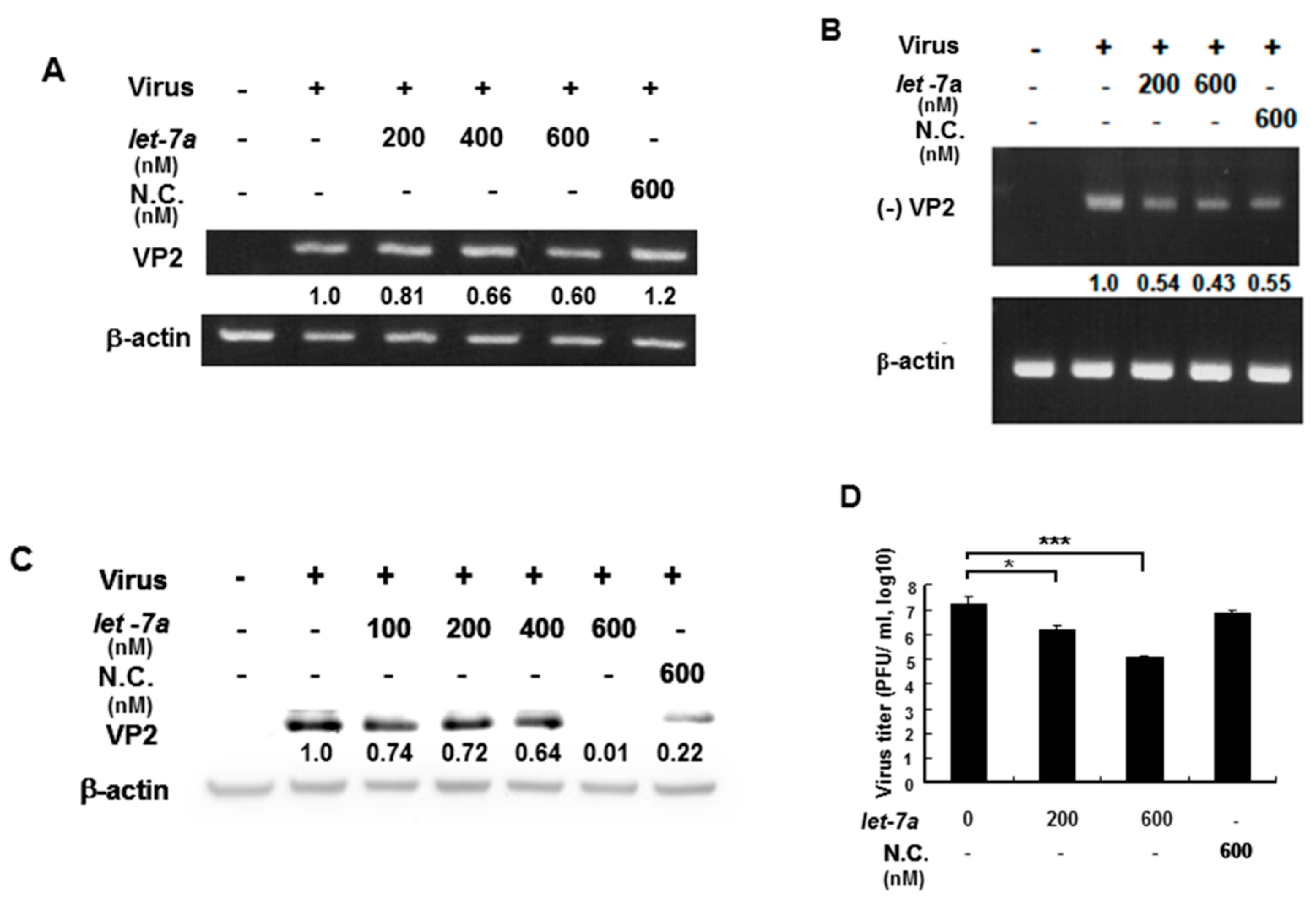
3.5. Let-7a Suppressed EV71 RNA Expression and Viral Titer in Various Infected Cell Lines
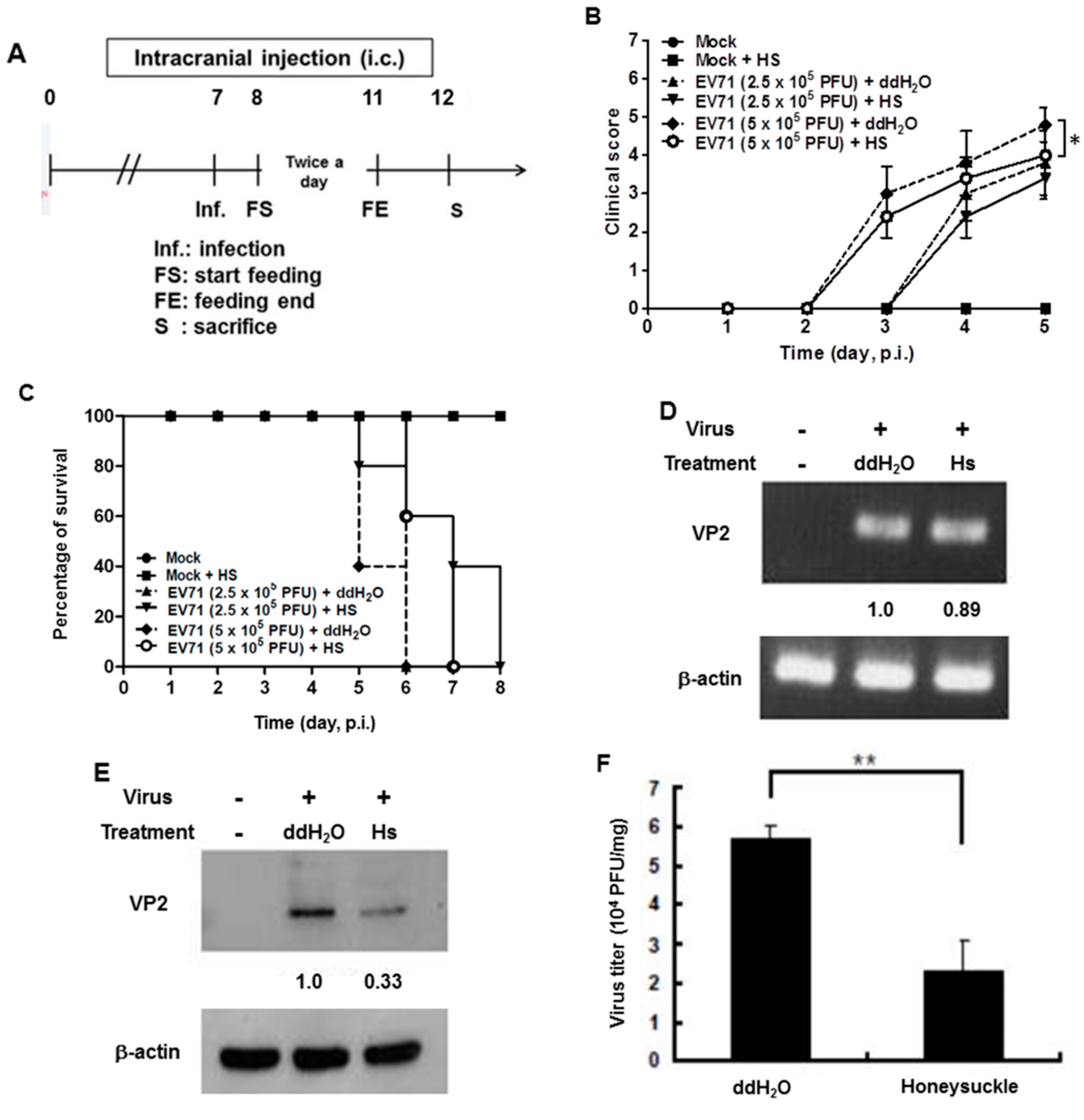
3.6. Honeysuckle Ingestion Attenuated Disease Symptoms, Prolonged Survival Time and Suppressed Viral Replication as Well as Viral Titer in EV71 MP4-Infected Suckling Mice
3.7. Let-7a Treatment Alleviated Clinical Scores, Prolonged Survival Time and Suppressed Viral Replication and Viral Titer in EV71 MP4-Infected Suckling Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shang, X.; Pan, H.; Li, M.; Miao, X.; Ding, H. Lonicera japonica thunb.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional chinese medicine. J. Ethnopharmacol. 2011, 138, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Yeh, S.F.; Ruan, X.M.; Zhang, H.; Hsu, S.D.; Huang, H.D.; Hsieh, C.C.; Lin, Y.S.; Yeh, T.M.; Liu, H.S.; et al. Honeysuckle aqueous extract and induced let-7a suppress dengue virus type 2 replication and pathogenesis. J. Ethnopharmacol. 2017, 198, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. Rna silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Chapman, E.J.; Carrington, J.C. Specialization and evolution of endogenous small rna pathways. Nat. Rev. Genet. 2007, 8, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Stefani, G.; Slack, F.J. Small non-coding rnas in animal development. Nat. Rev. Mol. Cell Biol. 2008, 9, 219–230. [Google Scholar] [CrossRef]
- Lu, L.F.; Liston, A. Microrna in the immune system, microrna as an immune system. Immunology 2009, 127, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Scaria, V.; Hariharan, M.; Maiti, S.; Pillai, B.; Brahmachari, S.K. Host-virus interaction: A new role for micrornas. Retrovirology 2006, 3, 68. [Google Scholar]
- Hsu, P.W.; Lin, L.Z.; Hsu, S.D.; Hsu, J.B.; Huang, H.D. Vita: Prediction of host micrornas targets on viruses. Nucleic Acids Res. 2007, 35, D381–D385. [Google Scholar] [CrossRef]
- Ahluwalia, J.K.; Khan, S.Z.; Soni, K.; Rawat, P.; Gupta, A.; Hariharan, M.; Scaria, V.; Lalwani, M.; Pillai, B.; Mitra, D.; et al. Human cellular microrna hsa-mir-29a interferes with viral nef protein expression and hiv-1 replication. Retrovirology 2008, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Breving, K.; Esquela-Kerscher, A. The complexities of microrna regulation: Mirandering around the rules. Int. J. Biochem. Cell Biol. 2010, 42, 1316–1329. [Google Scholar] [CrossRef]
- Ciesla, M.; Skrzypek, K.; Kozakowska, M.; Loboda, A.; Jozkowicz, A.; Dulak, J. Micrornas as biomarkers of disease onset. Anal. Bioanal. Chem. 2011, 401, 2051–2061. [Google Scholar] [CrossRef] [PubMed]
- Ajit, S.K. Circulating micrornas as biomarkers, therapeutic targets, and signaling molecules. Sensors 2012, 12, 3359–3369. [Google Scholar] [CrossRef]
- Kren, B.T.; Wong, P.Y.; Sarver, A.; Zhang, X.; Zeng, Y.; Steer, C.J. Micrornas identified in highly purified liver-derived mitochondria may play a role in apoptosis. RNA Biol. 2009, 6, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mrnas and micrornas is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar]
- Xu, J.; Qian, Y.; Wang, S.; Serrano, J.M.; Li, W.; Huang, Z.; Lu, S. Ev71: An emerging infectious disease vaccine target in the far east? Vaccine 2010, 28, 3516–3521. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.L.; Chiang, P.S.; Chia, M.Y.; Luo, S.T.; Chang, L.Y.; Lin, T.Y.; Ho, M.S.; Lee, M.S. Cross-reactive neutralizing antibody responses to enterovirus 71 infections in young children: Implications for vaccine development. PLoS Negl. Trop. Dis. 2013, 7, e2067. [Google Scholar] [CrossRef]
- Shih, S.R.; Stollar, V.; Li, M.L. Host factors in enterovirus 71 replication. J. Virol. 2011, 85, 9658–9666. [Google Scholar] [CrossRef] [PubMed]
- McMinn, P.C. Recent advances in the molecular epidemiology and control of human enterovirus 71 infection. Curr. Opin. Virol. 2012, 2, 199–205. [Google Scholar] [CrossRef]
- Solomon, T.; Lewthwaite, P.; Perera, D.; Cardosa, M.J.; McMinn, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet. Infect. Dis. 2010, 10, 778–790. [Google Scholar]
- Liu, Q.; Huang, X.; Ku, Z.; Wang, T.; Liu, F.; Cai, Y.; Li, D.; Leng, Q.; Huang, Z. Characterization of enterovirus 71 capsids using subunit protein-specific polyclonal antibodies. J. Virol. Methods 2013, 187, 127–131. [Google Scholar] [CrossRef]
- Denizot, M.; Neal, J.W.; Gasque, P. Encephalitis due to emerging viruses: Cns innate immunity and potential therapeutic targets. J. Infect. 2012, 65, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fowlkes, A.L.; Honarmand, S.; Glaser, C.; Yagi, S.; Schnurr, D.; Oberste, M.S.; Anderson, L.; Pallansch, M.A.; Khetsuriani, N. Enterovirus-associated encephalitis in the california encephalitis project, 1998–2005. J. Infect. Dis. 2008, 198, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.Y. Enterovirus 71 in taiwan. Pediatrics Neonatol. 2008, 49, 103–112. [Google Scholar] [CrossRef]
- Huang, H.I.; Weng, K.F.; Shih, S.R. Viral and host factors that contribute to pathogenicity of enterovirus 71. Future Microbiol. 2012, 7, 467–479. [Google Scholar] [CrossRef]
- Weng, K.F.; Chen, L.L.; Huang, P.N.; Shih, S.R. Neural pathogenesis of enterovirus 71 infection. Microbes Infect. 2010, 12, 505–510. [Google Scholar]
- Hamaguchi, T.; Fujisawa, H.; Sakai, K.; Okino, S.; Kurosaki, N.; Nishimura, Y.; Shimizu, H.; Yamada, M. Acute encephalitis caused by intrafamilial transmission of enterovirus 71 in adult. Emerg. Infect. Dis. 2008, 14, 828–830. [Google Scholar] [CrossRef]
- Chatproedprai, S.; Theanboonlers, A.; Korkong, S.; Thongmee, C.; Wananukul, S.; Poovorawan, Y. Clinical and molecular characterization of hand-foot-and-mouth disease in thailand, 2008–2009. Jpn. J. Infect. Dis. 2010, 63, 229–233. [Google Scholar] [PubMed]
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of sars coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Ruan, Q.; Yang, K.; Wang, W.; Jiang, L.; Song, J. Clinical predictors of mortality due to covid-19 based on an analysis of data of 150 patients from wuhan, china. Intensive Care Med 2020, 46, 846–848. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in wuhan, china. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Russell, C.D.; Millar, J.E.; Baillie, J.K. Clinical evidence does not support corticosteroid treatment for 2019-ncov lung injury. Lancet 2020, 395, 473–475. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. Sars-cov-2 cell entry depends on ace2 and tmprss2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Kupferschmidt, K.; Cohen, J. Race to find covid-19 treatments accelerates. Science 2020, 367, 1412–1413. [Google Scholar] [CrossRef]
- Leng, Z.; Zhu, R.; Hou, W.; Feng, Y.; Yang, Y.; Han, Q.; Shan, G.; Meng, F.; Du, D.; Wang, S.; et al. Transplantation of ace2(-) mesenchymal stem cells improves the outcome of patients with covid-19 pneumonia. Aging Dis. 2020, 11, 216–228. [Google Scholar] [CrossRef]
- Sommerstein, R.; Kochen, M.M.; Messerli, F.H.; Grani, C. Coronavirus disease 2019 (covid-19): Do angiotensin-converting enzyme inhibitors/angiotensin receptor blockers have a biphasic effect? J. Am. Heart Assoc. 2020, 9, e016509. [Google Scholar] [PubMed]
- Koniari, I.; Kounis, N.G. Cardiac arrest caused by nafamostat mesilate: Kounis syndrome in the dialysis room? Kidney Res. Clin. Pract. 2017, 36, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Stokkermans, T.J.; Trichonas, G. Chloroquine and hydroxychloroquine toxicity. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Ding, Y.; Zeng, L.; Li, R.; Chen, Q.; Zhou, B.; Chen, Q.; Cheng, P.L.; Yutao, W.; Zheng, J.; Yang, Z.; et al. The chinese prescription lianhuaqingwen capsule exerts anti-influenza activity through the inhibition of viral propagation and impacts immune function. BMC Complement Altern. Med. 2017, 17, 130. [Google Scholar] [CrossRef]
- Poon, P.M.; Wong, C.K.; Fung, K.P.; Fong, C.Y.; Wong, E.L.; Lau, J.T.; Leung, P.C.; Tsui, S.K.; Wan, D.C.; Waye, M.M.; et al. Immunomodulatory effects of a traditional chinese medicine with potential antiviral activity: A self-control study. Am. J. Chin. Med. 2006, 34, 13–21. [Google Scholar] [CrossRef]
- Lau, J.T.; Leung, P.C.; Wong, E.L.; Fong, C.; Cheng, K.F.; Zhang, S.C.; Lam, C.W.; Wong, V.; Choy, K.M.; Ko, W.M. The use of an herbal formula by hospital care workers during the severe acute respiratory syndrome epidemic in hong kong to prevent severe acute respiratory syndrome transmission, relieve influenza-related symptoms, and improve quality of life: A prospective cohort study. J. Altern. Complement Med. 2005, 11, 49–55. [Google Scholar] [PubMed]
- Yang, Y.; Islam, M.S.; Wang, J.; Li, Y.; Chen, X. Traditional chinese medicine in the treatment of patients infected with 2019-new coronavirus (sars-cov-2): A review and perspective. Int. J. Biol. Sci. 2020, 16, 1708–1717. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhou, Q.; Li, Y.; Garner, L.V.; Watkins, S.P.; Carter, L.J.; Smoot, J.; Gregg, A.C.; Daniels, A.D.; Jervey, S.; et al. Research and development on therapeutic agents and vaccines for covid-19 and related human coronavirus diseases. ACS Cent. Sci. 2020, 6, 315–331. [Google Scholar] [CrossRef]
- Harrison, C. Coronavirus puts drug repurposing on the fast track. Nat. Biotechnol. 2020, 38, 379–381. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. Mirbase: Integrating microrna annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, F.; Yang, K.; Fang, S.; Bu, D.; Li, H.; Sun, L.; Hu, H.; Gao, K.; Wang, W.; et al. Symmap: An integrative database of traditional chinese medicine enhanced by symptom mapping. Nucleic Acids Res. 2019, 47, D1110–D1117. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. Mirtarbase update 2018: A resource for experimentally validated microrna-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Kamburov, A.; Stelzl, U.; Lehrach, H.; Herwig, R. The consensuspathdb interaction database: 2013 update. Nucleic Acids Res. 2013, 41, D793–D800. [Google Scholar] [CrossRef]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Wang, Y.F.; Chou, C.T.; Lei, H.Y.; Liu, C.C.; Wang, S.M.; Yan, J.J.; Su, I.J.; Wang, J.R.; Yeh, T.M.; Chen, S.H.; et al. A mouse-adapted enterovirus 71 strain causes neurological disease in mice after oral infection. J. Virol. 2004, 78, 7916–7924. [Google Scholar] [CrossRef]
- Huang, S.W.; Wang, Y.F.; Yu, C.K.; Su, I.J.; Wang, J.R. Mutations in vp2 and vp1 capsid proteins increase infectivity and mouse lethality of enterovirus 71 by virus binding and rna accumulation enhancement. Virology 2012, 422, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Wang, P.S.; Wang, J.R.; Liu, H.S. Enterovirus 71-induced autophagy increases viral replication and pathogenesis in a suckling mouse model. J. Biomed. Sci. 2014, 21, 80. [Google Scholar] [CrossRef]
- Kung, Y.H.; Huang, S.W.; Kuo, P.H.; Kiang, D.; Ho, M.S.; Liu, C.C.; Yu, C.K.; Su, I.J.; Wang, J.R. Introduction of a strong temperature-sensitive phenotype into enterovirus 71 by altering an amino acid of virus 3d polymerase. Virology 2010, 396, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.R.; Tuan, Y.C.; Tsai, H.P.; Yan, J.J.; Liu, C.C.; Su, I.J. Change of major genotype of enterovirus 71 in outbreaks of hand-foot-and-mouth disease in taiwan between 1998 and 2000. J. Clin. Microbiol. 2002, 40, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Poh, C.L.; Chow, V.T. Complete sequence analyses of enterovirus 71 strains from fatal and non-fatal cases of the hand, foot and mouth disease outbreak in singapore (2000). Microbiol. Immunol. 2002, 46, 801–808. [Google Scholar] [CrossRef]
- Tan, S.H.; Ong, K.C.; Wong, K.T. Enterovirus 71 can directly infect the brainstem via cranial nerves and infection can be ameliorated by passive immunization. J. Neuropathol. Exp. Neurol. 2014, 73, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.M.; Chen, I.C.; Su, L.Y.; Huang, K.J.; Lei, H.Y.; Liu, C.C. Enterovirus 71 infection of monocytes with antibody-dependent enhancement. Clin. Vaccine Immunol. 2010, 17, 1517–1523. [Google Scholar] [CrossRef]
- Hussain, K.M.; Leong, K.L.; Ng, M.M.; Chu, J.J. The essential role of clathrin-mediated endocytosis in the infectious entry of human enterovirus 71. J. Biol. Chem. 2011, 286, 309–321. [Google Scholar]
- Lin, J.Y.; Shih, S.R.; Pan, M.; Li, C.; Lue, C.F.; Stollar, V.; Li, M.L. Hnrnp a1 interacts with the 5′ untranslated regions of enterovirus 71 and sindbis virus rna and is required for viral replication. J. Virol. 2009, 83, 6106–6114. [Google Scholar] [CrossRef]
- Michlewski, G.; Caceres, J.F. Antagonistic role of hnrnp a1 and ksrp in the regulation of let-7a biogenesis. Nat. Struct. Mol. Biol. 2010, 17, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- De Vito, C.; Riggi, N.; Suva, M.L.; Janiszewska, M.; Horlbeck, J.; Baumer, K.; Provero, P.; Stamenkovic, I. Let-7a is a direct ews-fli-1 target implicated in ewing’s sarcoma development. PLoS ONE 2011, 6, e23592. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 rna regulates developmental timing in caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Muller, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory rna. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific micrornas from mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression profiling of mammalian micrornas uncovers a subset of brain-expressed micrornas with possible roles in murine and human neuronal differentiation. Genome Biol. 2004, 5, R13. [Google Scholar] [CrossRef]
- Johnson, S.M.; Grosshans, H.; Shingara, J.; Byrom, M.; Jarvis, R.; Cheng, A.; Labourier, E.; Reinert, K.L.; Brown, D.; Slack, F.J. Ras is regulated by the let-7 microrna family. Cell 2005, 120, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Akao, Y.; Nakagawa, Y.; Naoe, T. Let-7 microrna functions as a potential growth suppressor in human colon cancer cells. Biol. Pharm. Bull. 2006, 29, 903–906. [Google Scholar]
- Sampson, V.B.; Rong, N.H.; Han, J.; Yang, Q.; Aris, V.; Soteropoulos, P.; Petrelli, N.J.; Dunn, S.P.; Krueger, L.J. Microrna let-7a down-regulates myc and reverts myc-induced growth in burkitt lymphoma cells. Cancer Res. 2007, 67, 9762–9770. [Google Scholar] [CrossRef]
- He, X.Y.; Chen, J.X.; Zhang, Z.; Li, C.L.; Peng, Q.L.; Peng, H.M. The let-7a microrna protects from growth of lung carcinoma by suppression of k-ras and c-myc in nude mice. J. Cancer Res. Clin. Oncol. 2010, 136, 1023–1028. [Google Scholar] [CrossRef]
- Lee, Y.S.; Dutta, A. The tumor suppressor microrna let-7 represses the hmga2 oncogene. Genes Dev. 2007, 21, 1025–1030. [Google Scholar] [CrossRef]
- Motoyama, K.; Inoue, H.; Nakamura, Y.; Uetake, H.; Sugihara, K.; Mori, M. Clinical significance of high mobility group a2 in human gastric cancer and its relationship to let-7 microrna family. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 2334–2340. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Fu, M.; Bookout, A.L.; Kliewer, S.A.; Mangelsdorf, D.J. Microrna let-7 regulates 3t3-l1 adipogenesis. Mol. Endocrinol. 2009, 23, 925–931. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Duan, C.; Chen, J.; Ou-Yang, X.; Zhang, Z.; Li, C.; Peng, H. Let-7a elevates p21(waf1) levels by targeting of nirf and suppresses the growth of a549 lung cancer cells. FEBS Lett. 2009, 583, 3501–3507. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Meng, P.; Wang, T.; Qin, W.; Qin, W.; Wang, F.; Yuan, J.; Chen, Z.; Yang, A.; Wang, H. Microrna let-7a inhibits proliferation of human prostate cancer cells in vitro and in vivo by targeting e2f2 and ccnd2. PLoS ONE 2010, 5, e10147. [Google Scholar] [CrossRef]
- Muller, D.W.; Bosserhoff, A.K. Integrin beta 3 expression is regulated by let-7a mirna in malignant melanoma. Oncogene 2008, 27, 6698–6706. [Google Scholar]
- Tsang, W.P.; Kwok, T.T. Let-7a microrna suppresses therapeutics-induced cancer cell death by targeting caspase-3. Apoptosis Int. J. Program. Cell Death 2008, 13, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Tang, Q.L.; Shang, Y.X.; Liang, S.B.; Yang, M.; Robinson, N.; Liu, J.P. Can chinese medicine be used for prevention of corona virus disease 2019 (covid-19)? A review of historical classics, research evidence and current prevention programs. Chin. J. Integr. Med. 2020, 26, 243–250. [Google Scholar] [CrossRef]
- Zhou, L.K.; Zhou, Z.; Jiang, X.M.; Zheng, Y.; Chen, X.; Fu, Z.; Xiao, G.; Zhang, C.Y.; Zhang, L.K.; Yi, Y. Absorbed plant mir2911 in honeysuckle decoction inhibits sars-cov-2 replication and accelerates the negative conversion of infected patients. Cell Discov. 2020, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Runfeng, L.; Yunlong, H.; Jicheng, H.; Weiqi, P.; Qinhai, M.; Yongxia, S.; Chufang, L.; Jin, Z.; Zhenhua, J.; Haiming, J.; et al. Lianhuaqingwen exerts anti-viral and anti-inflammatory activity against novel coronavirus (sars-cov-2). Pharm. Res. 2020, 104761. [Google Scholar] [CrossRef]
- Gao, Y.; Fang, L.; Cai, R.; Zong, C.; Chen, X.; Lu, J.; Qi, Y. Shuang-huang-lian exerts anti-inflammatory and anti-oxidative activities in lipopolysaccharide-stimulated murine alveolar macrophages. Phytomedicine 2014, 21, 461–469. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, H.; Sun, X.; Ding, M.; Tao, G.; Li, X. Honeysuckle-derived microrna2911 directly inhibits varicella-zoster virus replication by targeting ie62 gene. J. Neurovirol. 2019, 25, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microrna2911 directly targets influenza a viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J.; Hlh Across Speciality Collaboration, U.K. Covid-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Moshiashvili, G.; Tabatadze, N.; Mshvildadze, V. The genus daphne: A review of its traditional uses, phytochemistry and pharmacology. Fitoterapia 2020, 143, 104540. [Google Scholar] [CrossRef]
- Zhong, T.; Zhang, L.Y.; Wang, Z.Y.; Wang, Y.; Song, F.M.; Zhang, Y.H.; Yu, J.H. Rheum emodin inhibits enterovirus 71 viral replication and affects the host cell cycle environment. Acta Pharmacol. Sin. 2017, 38, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tao, L.; Xu, H. Chinese herbal medicines as a source of molecules with anti-enterovirus 71 activity. Chin. Med. 2016, 11, 2. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enriched Pathway-Based Sets | q-Value # | Source |
|---|---|---|
| Interleukin-4 and Interleukin-13 signaling | 4.82E-29 | Wikipathways |
| Human cytomegalovirus infection | 2.40E-22 | KEGG |
| Kaposi sarcoma-associated herpesvirus infection | 6.30E-20 | KEGG |
| Human T-cell leukemia virus 1 infection | 1.94E-18 | KEGG |
| Epstein–Barr virus infection | 2.43E-18 | KEGG |
| IL-17 signaling pathway | 4.20E-17 | KEGG |
| IL-2 receptor beta chain in t cell activation | 4.41E-12 | BioCarta |
| Validated transcriptional targets of AP1 family members Fra1 and Fra2 | 5.32E-12 | PID |
| Human immunodeficiency virus 1 infection | 8.74E-11 | KEGG |
| Human papillomavirus infection | 1.41E-10 | KEGG |
| IL-2-mediated signaling events | 3.35E-10 | PID |
| IL-2 Signaling Pathway | 4.95E-10 | Wikipathways |
| Interleukin-10 signaling | 4.18E-09 | Wikipathways |
| IL-5 Signaling Pathway | 6.59E-09 | Wikipathways |
| IL-6 | 8.49E-09 | NetPath |
| IL-3 Signaling Pathway | 3.56E-08 | Wikipathways |
| IL-11 | 4.30E-08 | NetPath |
| IL-23-mediated signaling events | 6.36E-08 | PID |
| IL-7 Signaling Pathway | 7.65E-08 | Wikipathways |
| IL-4 Signaling Pathway | 7.99E-08 | Wikipathways |
| Interleukin-11 Signaling Pathway | 2.44E-07 | Wikipathways |
| IL-2 signaling events mediated by STAT5 | 2.80E-07 | PID |
| IL-6-mediated signaling events | 3.97E-07 | PID |
| Regulation of Telomerase | 5.31E-07 | PID |
| IL-1 | 1.08E-06 | NetPath |
| IL-2 signaling events mediated by PI3K | 1.11E-06 | PID |
| Signal transduction through il1r | 1.11E-06 | BioCarta |
| IL-1 signaling pathway | 1.22E-06 | Wikipathways |
| IL-2 | 1.29E-06 | NetPath |
| Ras signaling in the CD4+ TCR pathway | 1.91E-06 | PID |
| IL-27-mediated signaling events | 2.20E-06 | PID |
| Ras signaling pathway | 2.82E-06 | KEGG |
| Fc-epsilon receptor I signaling in mast cells | 2.83E-06 | PID |
| IL-4 | 3.16E-06 | NetPath |
| Human cytomegalovirus and map kinase pathways | 3.74E-06 | BioCarta |
| Ras Signaling | 7.25E-06 | Wikipathways |
| Signaling by Interleukins | 7.27E-06 | Reactome |
| Natural killer cell-mediated cytotoxicity | 1.17E-05 | KEGG |
| Activation of the AP-1 family of transcription factors | 1.39E-05 | Reactome |
| IL-6 signaling pathway | 3.54E-05 | Wikipathways |
| IL-4-mediated signaling events | 3.66E-05 | PID |
| IL-12-mediated signaling events | 3.66E-05 | PID |
| IL-7 signaling | 3.76E-05 | INOH |
| IL-10 anti-inflammatory signaling pathway | 4.00E-05 | BioCarta |
| Fc epsilon RI signaling pathway | 4.30E-05 | KEGG |
| IL8- and CXCR1-mediated signaling events | 5.36E-05 | PID |
| Fc epsilon receptor i signaling in mast cells | 6.24E-05 | BioCarta |
| Nfkb activation by nontypeable hemophilus influenzae | 6.24E-05 | BioCarta |
| IL-3 | 8.49E-05 | NetPath |
| IL-12 signaling mediated by STAT4 | 9.82E-05 | PID |
| IL-8- and CXCR2-mediated signaling events | 0.000129 | PID |
| West Nile virus | 0.000231 | BioCarta |
| MAPK family signaling cascades | 0.000287 | Reactome |
| Ccr3 signaling in eosinophils | 0.000346 | BioCarta |
| Ebola Virus Pathway on Host | 0.00037 | Wikipathways |
| Structural Pathway of Interleukin 1 (IL-1) | 0.000392 | Wikipathways |
| IL-1 and megakaryocytes in obesity | 0.0004 | Wikipathways |
| Signaling pathway from g-protein families | 0.000528 | BioCarta |
| IL-5 | 0.000562 | NetPath |
| IL-7 | 0.000607 | NetPath |
| IL-17 signaling pathway | 0.000985 | Wikipathways |
| Transcriptional regulation by the AP-2 (TFAP2) family of transcription factors | 0.00113 | Wikipathways |
| Regulation of Ras family activation | 0.00123 | PID |
| Calcium signaling by hbx of hepatitis b virus | 0.00162 | BioCarta |
| IL-1-mediated signaling events | 0.00162 | PID |
| IL-1 NFkB | 0.00181 | INOH |
| IL-9 Signaling Pathway | 0.00187 | Wikipathways |
| Transcriptional regulation by the AP-2 (TFAP2) family of transcription factors | 0.00228 | Reactome |
| TFAP2 (AP-2) family regulates transcription of cell cycle factors | 0.00246 | Reactome |
| Ras signaling pathway | 0.0028 | BioCarta |
| Fc epsilon receptor (FCERI) signaling | 0.00348 | Reactome |
| Interleukin-1 processing | 0.00467 | Reactome |
| FoxO family signaling | 0.00467 | PID |
| IPA Canonical Pathways | −log (p-Value) |
|---|---|
| IL-12 signaling and production in macrophages | 17.4 |
| IL-8 signaling | 14.8 |
| IL-6 signaling | 12.7 |
| Role of PKR in interferon induction and antiviral response | 12.2 |
| IL-10 signaling | 12 |
| Role of pattern recognition receptors in recognition of bacteria and viruses | 12 |
| IL-7 signaling pathway | 10.1 |
| TNFR1 signaling | 9.79 |
| ILK signaling | 9.45 |
| IL-17A signaling in fibroblasts | 8.9 |
| IL-3 signaling | 8.23 |
| TNFR2 signaling | 7.5 |
| NF-κB activation by viruses | 6.61 |
| Differential regulation of cytokine production in macrophages and T helper cells by IL-17A and IL-17F | 6.55 |
| IL-23 signaling pathway | 6.55 |
| Differential regulation of cytokine production in intestinal epithelial cells by IL-17A and IL-17F | 6.1 |
| IL-15 signaling | 5.5 |
| IL-17 signaling | 5.25 |
| Regulation of IL-2 expression in activated and anergic T lymphocytes | 5.04 |
| IL-17A signaling in airway cells | 4.28 |
| LPS/IL-1-mediated inhibition of RXR function | 4.25 |
| IL-1 signaling | 3.69 |
| Mechanisms of viral exit from host cells | 3.52 |
| Natural killer cell signaling | 3.27 |
| IL-2 signaling | 3.01 |
| Virus entry via endocytic pathways | 2.31 |
| IL-9 signaling | 2.3 |
| IL-15 production | 2.19 |
| Role of RIG1-like receptors in antiviral innate immunity | 2.1 |
| IL-4 signaling | 1.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-R.; Chang, C.-M.; Yeh, Y.-C.; Huang, C.-Y.F.; Lin, F.-M.; Huang, J.-T.; Hsieh, C.-C.; Wang, J.-R.; Liu, H.-S. Honeysuckle Aqueous Extracts Induced let-7a Suppress EV71 Replication and Pathogenesis In Vitro and In Vivo and Is Predicted to Inhibit SARS-CoV-2. Viruses 2021, 13, 308. https://doi.org/10.3390/v13020308
Lee Y-R, Chang C-M, Yeh Y-C, Huang C-YF, Lin F-M, Huang J-T, Hsieh C-C, Wang J-R, Liu H-S. Honeysuckle Aqueous Extracts Induced let-7a Suppress EV71 Replication and Pathogenesis In Vitro and In Vivo and Is Predicted to Inhibit SARS-CoV-2. Viruses. 2021; 13(2):308. https://doi.org/10.3390/v13020308
Chicago/Turabian StyleLee, Ying-Ray, Chia-Ming Chang, Yuan-Chieh Yeh, Chi-Ying F. Huang, Feng-Mao Lin, Juan-Ting Huang, Chang-Chi Hsieh, Jen-Ren Wang, and Hsiao-Sheng Liu. 2021. "Honeysuckle Aqueous Extracts Induced let-7a Suppress EV71 Replication and Pathogenesis In Vitro and In Vivo and Is Predicted to Inhibit SARS-CoV-2" Viruses 13, no. 2: 308. https://doi.org/10.3390/v13020308
APA StyleLee, Y.-R., Chang, C.-M., Yeh, Y.-C., Huang, C.-Y. F., Lin, F.-M., Huang, J.-T., Hsieh, C.-C., Wang, J.-R., & Liu, H.-S. (2021). Honeysuckle Aqueous Extracts Induced let-7a Suppress EV71 Replication and Pathogenesis In Vitro and In Vivo and Is Predicted to Inhibit SARS-CoV-2. Viruses, 13(2), 308. https://doi.org/10.3390/v13020308