
Virus Etiology, Diversity and Clinical Characteristics in South African Children Hospitalised with Gastroenteritis

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Study Population
2.3. Stool Specimen Preparation and Nucleic Acid Extraction
2.4. Enteric Virus Detection
2.5. Genotyping of Enteric Viruses
2.5.1. Complementary DNA Synthesis
2.5.2. Calicivirus Genotyping
2.5.3. Rotavirus Genotyping
2.6. Phylogenetic Analyses
2.7. Total Genomic DNA Extraction from Whole Blood Samples
2.8. Nonsense Variation Detection Using Real-Time PCR and SNP Assay
2.9. FUT2 Genotyping of Non-Secretor Samples
2.10. Statistical Analyses
3. Results
3.1. Study Population
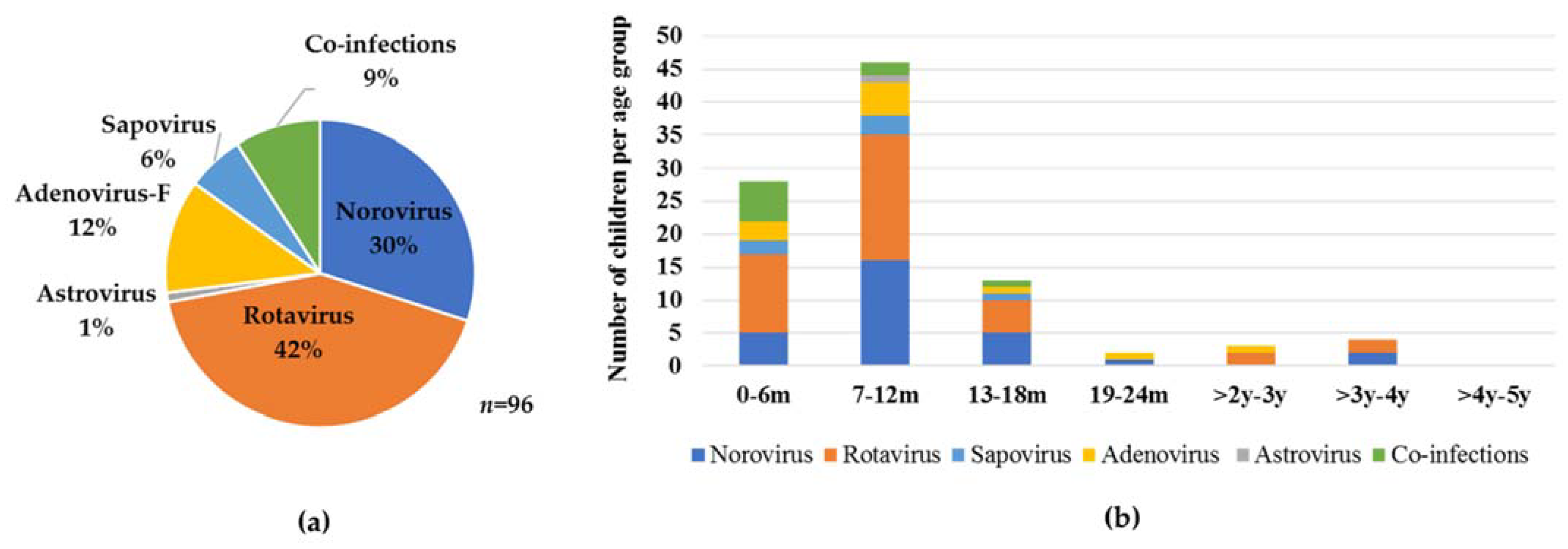
3.2. Virus Detection
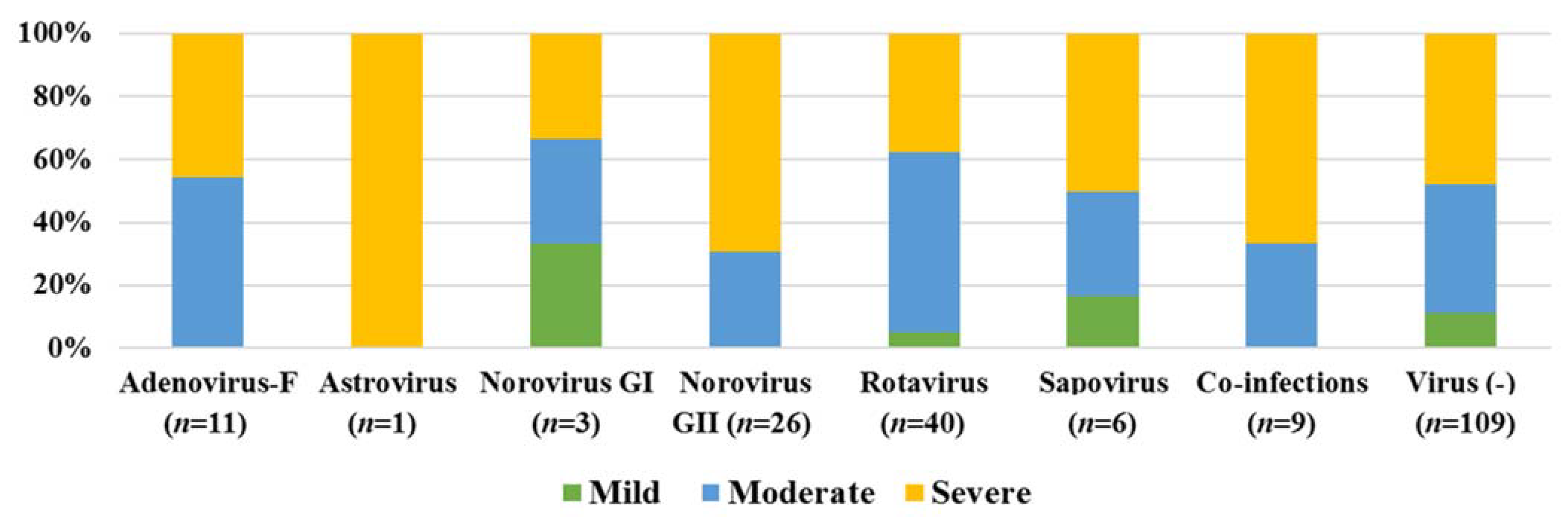
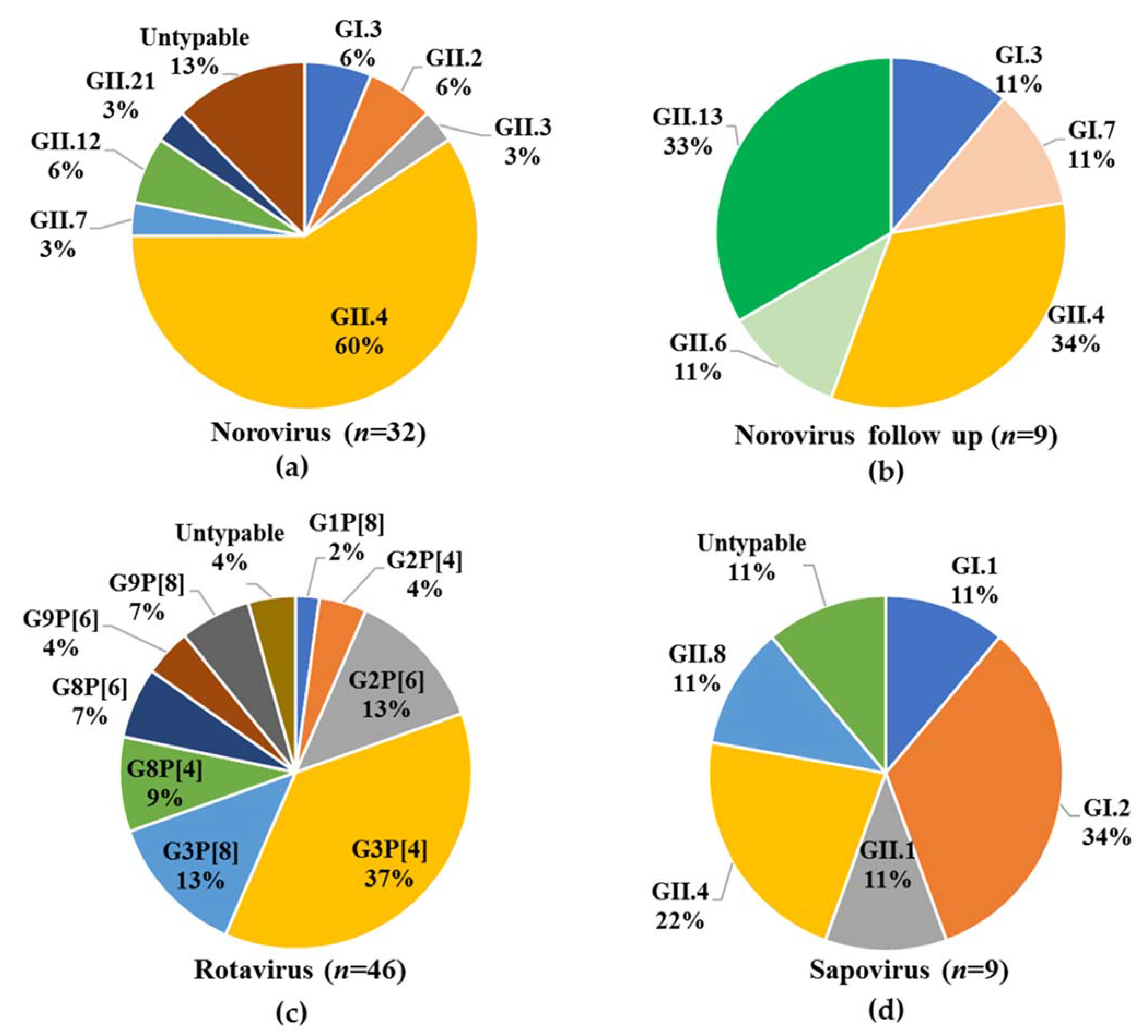
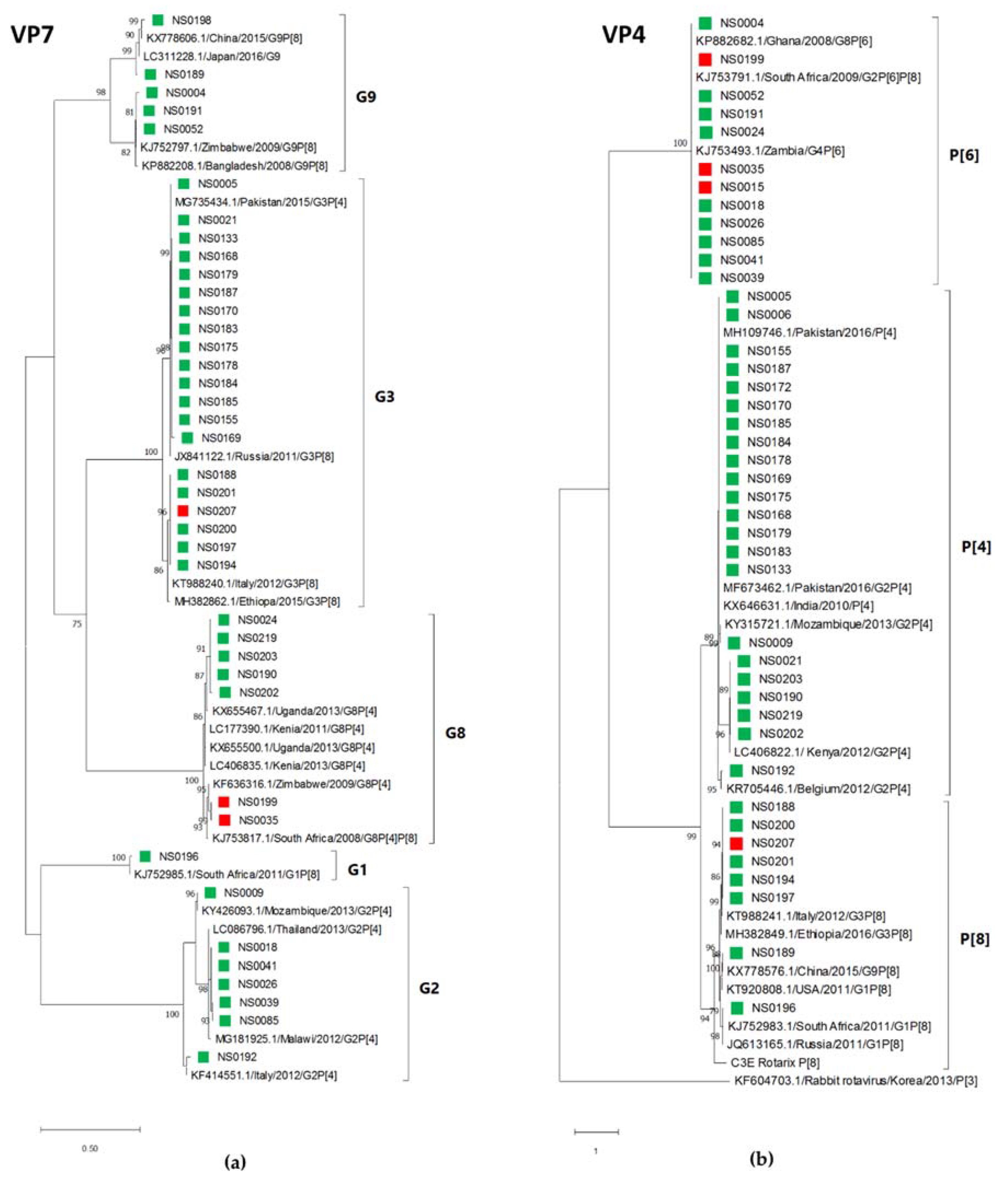
3.3. Virus Genotyping
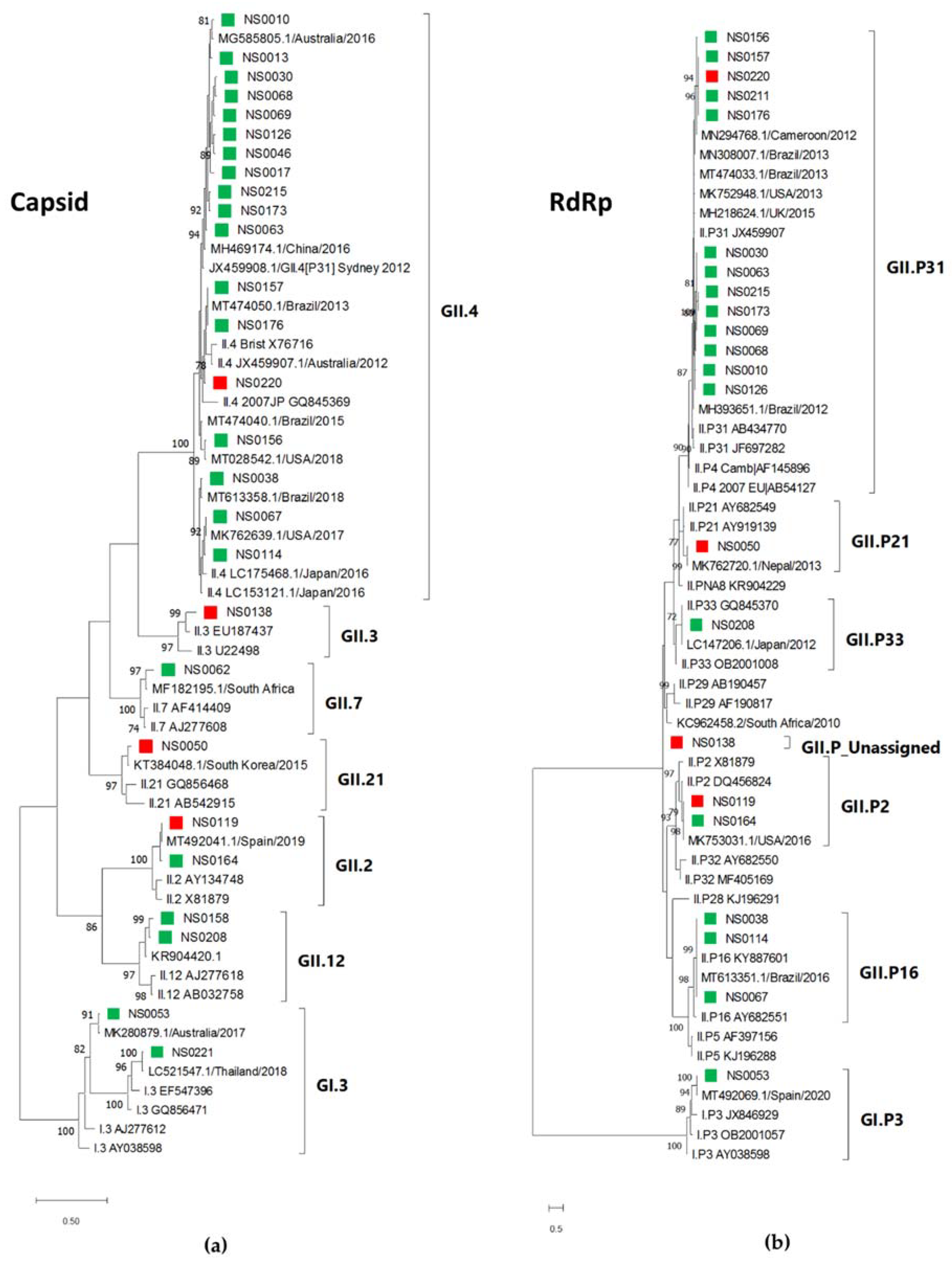
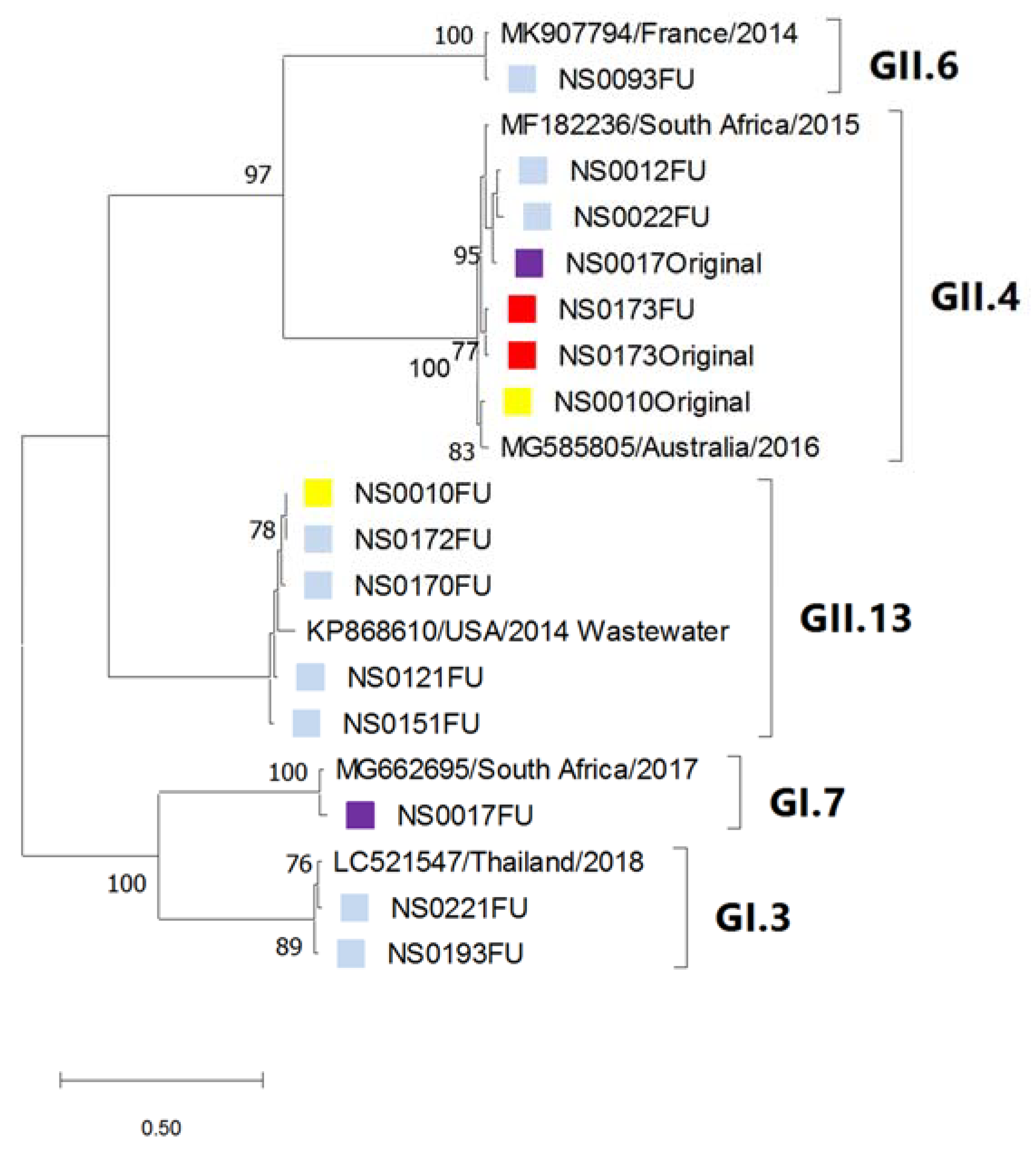
3.4. Phylogenetic Analysis
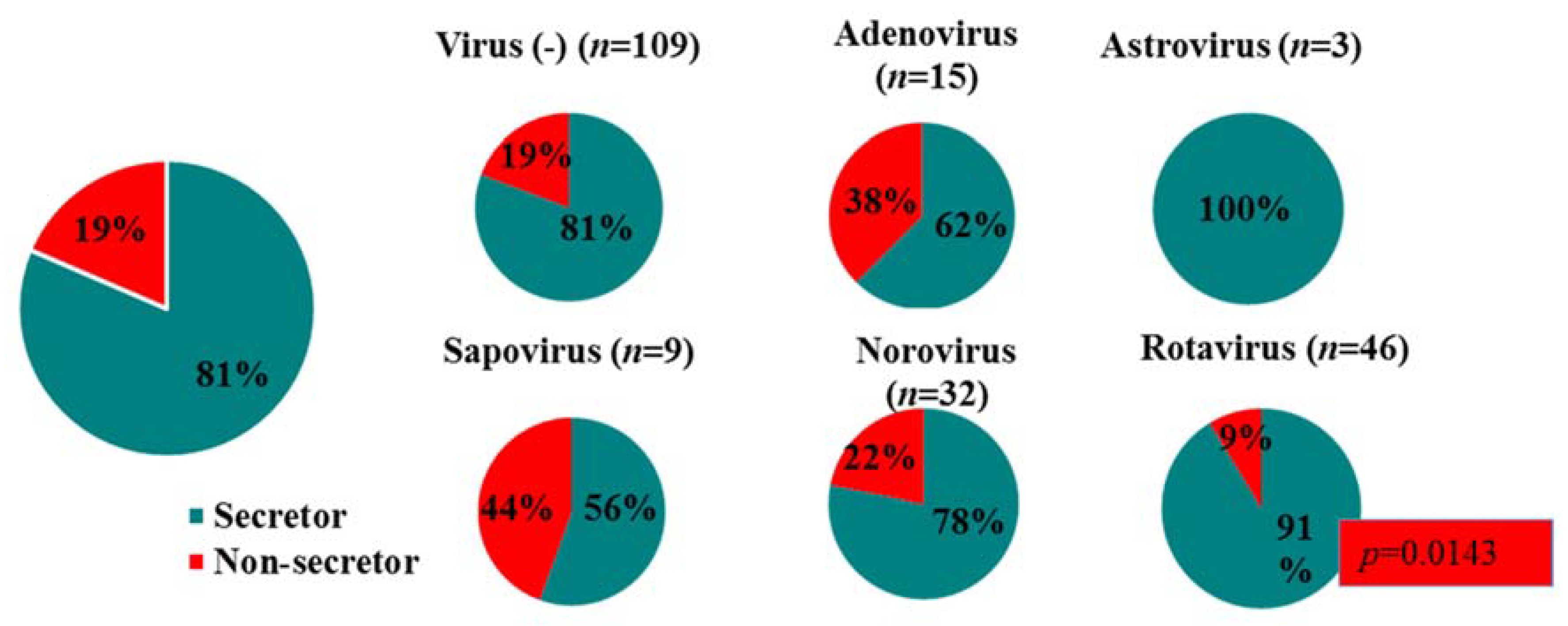
3.5. Secretor Status and Virus Genotypes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Institute for Health Metrics and Evaluation (IHME). Findings from the Global Burden of Disease Study 2017; IHME: Seattle, WA, USA, 2018. [Google Scholar]
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, G.; Bines, J.E.; Brewer, T.G.; Colombara, D.V. Rotavirus vaccination and the global burden of rotavirus diarrhea among children younger than 5 years. JAMA Pediatr. 2018, 172, 958–965. [Google Scholar] [CrossRef]
- Parashar, U.D.; Burton, A.; Lanata, C.; Boschi-Pinto, C.; Shibuya, K.; Steele, D.; Birmingham, M.; Glass, R.I. Global mortality associated with rotavirus disease among children in 2004. J. Infect. Dis. 2009, 200 (Suppl. 1), S9–S15. [Google Scholar] [CrossRef] [PubMed]
- Burnett, E.; Jonesteller, C.L.; Tate, J.E.; Yen, C.; Parashar, U.D. Global impact of rotavirus vaccination on childhood hospitalizations and mortality from diarrhea. J. Infect. Dis. 2017, 215, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Hallowell, B.D.P.; Parashar, U.D.; Curns, A.; DeGroote, N.P.; Tate, J.E. Trends in the laboratory detection of rotavirus before and after implementation of routine rotavirus vaccination—United States, 2000–2018. MMWR 2019, 68, 539–543. [Google Scholar] [PubMed]
- Govender, N.; Perovic, O.; Meiring, S.; Kleynhans, J.; von Gottberg, A.; Thomas, J.; Ismail, F.; Ismail, N.; Page, N.A.; Hunt, G.; et al. GERMS-SA Annual Report 2017; National Institute for Communicable Diseases: Johannesburg, South Africa, 2017; pp. 41–42. [Google Scholar]
- Govender, N.; Mathebula, R.; Perovic, O.; Meiring, S.; Kleynhans, J.; von Gottberg, A.; Thomas, J.; Ismail, F.; Ismail, N.; Page, N.A.; et al. GERMS-SA Annual Report 2018; National Institute for Communicable Diseases: Johannesburg, South Africa, 2018; pp. 37–38. [Google Scholar]
- Mans, J.; Armah, G.E.; Steele, A.D.; Taylor, M.B. Norovirus epidemiology in Africa: A review. PLoS ONE 2016, 11, e0146280. [Google Scholar] [CrossRef]
- Page, N.; Groome, M.; Nadan, S.; Netshikweta, R.; Keddy, K.; Poonsamy, B.; Moyes, J.; Walaza, S.; Kahn, K.; Madhi, S.; et al. Norovirus epidemiology in South African children <5 years hospitalised for diarrhoeal illness between 2009 and 2013. Epidemiol. Infect. 2017, 145, 1942–1952. [Google Scholar]
- Mans, J.; Murray, T.Y.; Nadan, S.; Netshikweta, R.; Page, N.A.; Taylor, M.B. Norovirus diversity in children with gastroenteritis in South Africa from 2009 to 2013: GII.4 variants and recombinant strains predominate. Epidemiol. Infect. 2016, 144, 907–916. [Google Scholar] [CrossRef]
- Esposito, S.; Principi, N. Norovirus Vaccine: Priorities for Future Research and Development. Front. Immunol. 2020, 11, 1383. [Google Scholar] [CrossRef]
- Phattanawiboon, B.; Nonthabenjawan, N.; Boonyos, P.; Jetsukontorn, C.; Towayunanta, W.; Chuntrakool, K.; Ngaopravet, K.; Ruchusatsawat, K.; Uppapong, B.; Sangkitporn, S.; et al. Norovirus transmission mediated by asymptomatic family members in households. PLoS ONE 2020, 15, e0236502. [Google Scholar] [CrossRef]
- Lopman, B.A.; Trivedi, T.; Vicuña, Y.; Costantini, V.; Collins, N.; Gregoricus, N.; Parashar, U.; Sandoval, C.; Broncano, N.; Vaca, M. Norovirus infection and disease in an Ecuadorian birth cohort: Association of certain norovirus genotypes with host FUT2 secretor status. J. Infect. Dis. 2014, 211, 1813–1821. [Google Scholar] [CrossRef]
- Payne, D.C.; Currier, R.L.; Staat, M.A.; Sahni, L.C.; Selvarangan, R.; Halasa, N.B.; Englund, J.A.; Weinberg, G.A.; Boom, J.A.; Szilagyi, P.G. Epidemiologic association between FUT2 secretor status and severe rotavirus gastroenteritis in children in the United States. JAMA Pediatr. 2015, 169, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Thorven, M.; Grahn, A.; Hedlund, K.-O.; Johansson, H.; Wahlfrid, C.; Larson, G.; Svensson, L. A homozygous nonsense mutation (428G→A) in the human secretor (FUT2) gene provides resistance to symptomatic norovirus (GGII) infections. J. Virol. 2005, 79, 15351–15355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-F.; Long, Y.; Tan, M.; Zhang, T.; Huang, Q.; Jiang, X.; Tan, W.-F.; Li, J.-D.; Hu, G.-F.; Tang, S. P [8] and P [4] rotavirus infection associated with secretor phenotypes among children in South China. Sci. Rep. 2016, 6, 34591. [Google Scholar] [CrossRef]
- Ferrer-Admetlla, A.; Sikora, M.; Laayouni, H.; Esteve, A.; Roubinet, F.; Blancher, A.; Calafell, F.; Bertranpetit, J.; Casals, F. A natural history of FUT2 polymorphism in humans. Mol. Biol. Evol. 2009, 26, 1993–2003. [Google Scholar] [CrossRef] [PubMed]
- Kindberg, E.; Svensson, L. Genetic basis of host resistance to norovirus infection. Future Virol. 2009, 4, 369–382. [Google Scholar] [CrossRef]
- Koda, Y.; Tachida, H.; Pang, H.; Liu, Y.; Soejima, M.; Ghaderi, A.A.; Takenaka, O.; Kimura, H. Contrasting patterns of polymorphisms at the ABO-secretor gene (FUT2) and plasma α (1, 3) fucosyltransferase gene (FUT6) in human populations. Genetics 2001, 158, 747–756. [Google Scholar] [PubMed]
- Nordgren, J.; Nitiema, L.W.; Ouermi, D.; Simpore, J.; Svensson, L. Host genetic factors affect susceptibility to norovirus infections in Burkina Faso. PLoS ONE 2013, 8, e69557. [Google Scholar] [CrossRef]
- Parker, E.P.; Ramani, S.; Lopman, B.A.; Church, J.A.; Iturriza-Gomara, M.; Prendergast, A.J.; Grassly, N.C. Causes of impaired oral vaccine efficacy in developing countries. Future Microbiol. 2018, 13, 97–118. [Google Scholar] [CrossRef]
- Lewis, Z.T.; Totten, S.M.; Smilowitz, J.T.; Popovic, M.; Parker, E.; Lemay, D.G.; Van Tassell, M.L.; Miller, M.J.; Jin, Y.-S.; German, J.B. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 2015, 3, 13. [Google Scholar] [CrossRef]
- Zivkovic, A.M.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Human milk glycobiome and its impact on the infant gastrointestinal microbiota. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4653–4658. [Google Scholar] [CrossRef]
- Colston, J.M.; Francois, R.; Pisanic, N.; Peñataro Yori, P.; McCormick, B.J.; Olortegui, M.P.; Gazi, M.A.; Svensen, E.; Ahmed, M.M.M.; Mduma, E. Effects of child and maternal histo-blood group antigen status on symptomatic and asymptomatic enteric infections in early childhood. J. Infect. Dis. 2019, 220, 151–162. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS Data 2019. Available online: https://www.unaids.org/sites/default/files/media_asset/2019-UNAIDS-data_en.pdf (accessed on 12 November 2019).
- Bunders, M.J.; Van Hamme, J.L.; Jansen, M.H.; Boer, K.; Kootstra, N.A.; Kuijpers, T.W. Fetal exposure to HIV-1 alters chemokine receptor expression by CD4+ T cells and increases susceptibility to HIV-1. Sci. Rep. 2014, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Epalza, C.; Goetghebuer, T.; Hainaut, M.; Prayez, F.; Barlow, P.; Dediste, A.; Marchant, A.; Levy, J. High incidence of invasive group B streptococcal infections in HIV-exposed uninfected infants. Pediatrics 2010, 126, e631–e638. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.; Jones, C.E.; Prendergast, A.J. HIV-exposed, uninfected infants: New global challenges in the era of paediatric HIV elimination. Lancet Inf. Dis. 2016, 16, e92–e107. [Google Scholar] [CrossRef]
- Feiterna-Sperling, C.; Weizsaecker, K.; Bührer, C.; Casteleyn, S.; Loui, A.; Schmitz, T.; Wahn, V.; Obladen, M. Hematologic effects of maternal antiretroviral therapy and transmission prophylaxis in HIV-1-exposed uninfected newborn infants. JAIDS 2007, 45, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Marinda, E.; Humphrey, J.H.; Iliff, P.J.; Mutasa, K.; Nathoo, K.J.; Piwoz, E.G.; Moulton, L.H.; Salama, P.; Ward, B.J.; Group, Z.S. Child mortality according to maternal and infant HIV status in Zimbabwe. Pediatr. Infect. Dis. J. 2007, 26, 519–526. [Google Scholar] [CrossRef]
- Slogrove, A.; Reikie, B.; Naidoo, S.; De Beer, C.; Ho, K.; Cotton, M.; Bettinger, J.; Speert, D.; Esser, M.; Kollmann, T. HIV-exposed uninfected infants are at increased risk for severe infections in the first year of life. J. Trop. Pediatr. 2012, 58, 505–508. [Google Scholar] [CrossRef]
- Rollins, N.C.; Ndirangu, J.; Bland, R.M.; Coutsoudis, A.; Coovadia, H.M.; Newell, M.-L. Exclusive breastfeeding, diarrhoeal morbidity and all-cause mortality in infants of HIV-infected and HIV uninfected mothers: An intervention cohort study in KwaZulu Natal, South Africa. PLoS ONE 2013, 8, e81307. [Google Scholar] [CrossRef]
- Brennan, A.T.; Bonawitz, R.; Gill, C.J.; Thea, D.M.; Kleinman, M.; Long, L.; McCallum, C.; Fox, M.P. A meta-analysis assessing diarrhea and pneumonia in HIV-exposed uninfected compared with HIV-unexposed uninfected infants and children. JAIDS 2019, 82, 1–8. [Google Scholar] [CrossRef]
- Brown, J.R.; Shah, D.; Breuer, J. Viral gastrointestinal infections and norovirus genotypes in a paediatric UK hospital, 2014–2015. J. Clin. Virol. 2016, 84, 1–6. [Google Scholar] [CrossRef]
- Freedman, S.B.; Eltorky, M.; Gorelick, M. Pediatric Emergency Research Canada Group Study, Evaluation of a gastroenteritis severity score for use in outpatient settings. Pediatrics 2010, 125, e1278–e1285. [Google Scholar] [CrossRef] [PubMed]
- Ruuska, T.; Vesikari, T. Rotavirus disease in Finnish children: Use of numerical scores for clinical severity of diarrhoeal episodes. Scand. J. Infect. Dis. 1990, 22, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Schnadower, D.; Tarr, P.I.; Gorelick, M.H.; O’Connell, K.; Roskind, C.G.; Powell, E.C.; Rao, J.; Bhatt, S.; Freedman, S.B. Validation of the modified Vesikari score in children with gastroenteritis in 5 US emergency departments. J. Pediatr. Gastroenterol. Nutr. 2013, 57, 514. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.K.; Le Saux, J.-C.; Parnaudeau, S.; Pommepuy, M.; Elimelech, M.; Le Guyader, F.S. Evaluation of removal of noroviruses during wastewater treatment, using real-time reverse transcription-PCR: Different behaviors of genogroups I and II. Appl. Environ. Microbiol. 2007, 73, 7891–7897. [Google Scholar] [CrossRef] [PubMed]
- Hoehne, M.; Schreier, E. Detection of Norovirus genogroup I and II by multiplex real-time RT-PCR using a 3’-minor groove binder-DNA probe. BMC Infect. Dis. 2006, 6, 69. [Google Scholar] [CrossRef]
- Kageyama, T.; Kojima, S.; Shinohara, M.; Uchida, K.; Fukushi, S.; Hoshino, F.B.; Takeda, N.; Katayama, K. Broadly reactive and highly sensitive assay for Norwalk-like viruses based on real-time quantitative reverse transcription-PCR. J. Clin. Microbiol. 2003, 41, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Loisy, F.; Atmar, R.; Guillon, P.; Le Cann, P.; Pommepuy, M.; Le Guyader, F. Real-time RT-PCR for norovirus screening in shellfish. J. Virol. Meth. 2005, 123, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Svraka, S.; Duizer, E.; Vennema, H.; de Bruin, E.; van der Veer, B.; Dorresteijn, B.; Koopmans, M. Etiological role of viruses in outbreaks of acute gastroenteritis in The Netherlands from 1994 through 2005. J. Clin. Microbiol. 2007, 45, 1389–1394. [Google Scholar] [CrossRef]
- Gentsch, J.; Gray, J.; Itturiza-Gomara, M.; Klena, J.; Kirkwood, C.; Armah, G.; Page, N. Manual of Rotavirus Detection and Characterization Methods; World Health Organization: Geneva, Switzerland, 2009; pp. 104–105. [Google Scholar]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98.
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kroneman, A.; Vennema, H.; Deforche, K.; Avoort, H.; Penaranda, S.; Oberste, M.; Vinjé, J.; Koopmans, M. An automated genotyping tool for enteroviruses and noroviruses. J. Clin. Virol. 2011, 51, 121–125. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.; Groome, M.J.; Mans, J.; Page, N. FUT2 Secretor Status Influences Susceptibility to VP4 Strain-Specific Rotavirus Infections in South African Children. Pathogens 2020, 9, 795. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of under-5 mortality in 2000–15: An updated systematic analysis with implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef]
- UNICEF. Estimates of Child Cause of Death, Diarrhoea 2018. Available online: https://data.unicef.org/topic/child-health/diarrhoeal-disease/ (accessed on 3 November 2020).
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Steele, A.D.; Duque, J.; Parashar, U.D. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Operario, D.J.; Platts-Mills, J.A.; Nadan, S.; Page, N.; Seheri, M.; Mphahlele, J.; Praharaj, I.; Kang, G.; Araujo, I.T.; Leite, J.P.G. Etiology of severe acute watery diarrhea in children in the global rotavirus surveillance network using quantitative polymerase chain reaction. J. Infect. Dis. 2017, 216, 220–227. [Google Scholar] [CrossRef]
- Anwari, P.; Safi, N.; Payne, D.C.; Jennings, M.C.; Rasikh, S.; Waciqi, A.S.; Parwiz, S.M. Rotavirus is the leading cause of hospitalizations for severe acute gastroenteritis among Afghan children <5 years old. Vaccine 2018, 36, 7765–7768. [Google Scholar]
- Liu, J.; Platts-Mills, J.A.; Juma, J.; Kabir, F.; Nkeze, J.; Okoi, C.; Operario, D.J.; Uddin, J.; Ahmed, S.; Alonso, P.L. Use of quantitative molecular diagnostic methods to identify causes of diarrhoea in children: A reanalysis of the GEMS case-control study. Lancet 2016, 388, 1291–1301. [Google Scholar] [CrossRef]
- Makgatho, E.; Patel, F.; Solomon, F.; Groome, M.J.; Lala, S.G.; Vallabh, P.; Dangor, Z. The Burden of Acute Diarrheal Disease in Young Hospitalized Urban South African Children Five Years after Rotavirus Vaccine Introduction: A Retrospective Descriptive Study. Pediatr. Infect. Dis. J. 2019, 38, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Nadan, S.; Taylor, M.B.; Groome, M.J.; Cohen, C.; Madhi, S.A.; Page, N.A. Epidemiology of human astroviruses among children younger than 5 years: Prospective hospital-based sentinel surveillance in South Africa, 2009–2014. J. Med. Virol. 2019, 91, 225–234. [Google Scholar] [CrossRef] [PubMed]
- The National Respiratory and Enteric Virus Surveillance System (NREVSS). Available online: https://www.cdc.gov/surveillance/nrevss/rotavirus/natl-trend.html (accessed on 20 January 2020).
- Lekana-Douki, S.E.; Kombila-Koumavor, C.; Nkoghe, D.; Drosten, C.; Drexler, J.F.; Leroy, E.M. Molecular epidemiology of enteric viruses and genotyping of rotavirus A, adenovirus and astrovirus among children under 5 years old in Gabon. Inter. J. Infect. Dis. 2015, 34, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Ouedraogo, N.; Kaplon, J.; Bonkoungou, I.J.O.; Traore, A.S.; Pothier, P.; Barro, N.; Ambert-Balay, K. Prevalence and genetic diversity of enteric viruses in children with diarrhea in Ouagadougou, Burkina Faso. PLoS ONE 2016, 11, e0153652. [Google Scholar] [CrossRef] [PubMed]
- Ronnelid, Y.; Bonkoungou, I.J.O.; Ouedraogo, N.; Barro, N.; Svensson, L.; Nordgren, J. Norovirus and rotavirus in children hospitalised with diarrhoea after rotavirus vaccine introduction in Burkina Faso. Epidemiol. Infect. 2020, 148, e245. [Google Scholar] [CrossRef] [PubMed]
- Kabue, J.P.; Meader, E.; Hunter, P.R.; Potgieter, N. Norovirus prevalence and estimated viral load in symptomatic and asymptomatic children from rural communities of Vhembe district, South Africa. J. Clin. Virol. 2016, 84, 12–18. [Google Scholar] [CrossRef]
- García, C.; DuPont, H.L.; Long, K.Z.; Santos, J.I.; Ko, G. Asymptomatic norovirus infection in Mexican children. J. Clin. Microbiol. 2006, 44, 2997–3000. [Google Scholar] [CrossRef]
- Mattioli, M.C.; Pickering, A.J.; Gilsdorf, R.J.; Davis, J.; Boehm, A.B. Hands and water as vectors of diarrheal pathogens in Bagamoyo, Tanzania. Environ. Sci. Technol. 2013, 47, 355–363. [Google Scholar] [CrossRef]
- UNICEF. Pneumonia and Diarrhoea-Tackling the Deadliest Diseases for the World’s Poorest Children. Available online: https://www.unicef.org/publications/index_65491.html (accessed on 3 November 2020).
- Mans, J. Norovirus Infections and Disease in Lower-Middle and Low-Income Countries, 1997–2018. Viruses 2019, 11, 341. [Google Scholar] [CrossRef]
- Seheri, L.M.; Magagula, N.B.; Peenze, I.; Rakau, K.; Ndadza, A.; Mwenda, J.M.; Weldegebriel, G.; Steele, A.D.; Mphahlele, M.J. Rotavirus strain diversity in Eastern and Southern African countries before and after vaccine introduction. Vaccine 2018, 36, 7222–7230. [Google Scholar] [CrossRef]
- Page, N.A.; Seheri, L.M.; Groome, M.J.; Moyes, J.; Walaza, S.; Mphahlele, J.; Kahn, K.; Kapongo, C.N.; Zar, H.J.; Tempia, S.; et al. Temporal association of rotavirus vaccination and genotype circulation in South Africa: Observations from 2002 to 2014. Vaccine 2018, 36, 7231–7237. [Google Scholar] [CrossRef] [PubMed]
- João, E.D.; Munlela, B.; Chissaque, A.; Chilaule, J.; Langa, J.; Augusto, O.; Boene, S.S.; Anapakala, E.; Sambo, J.; Guimarães, E.; et al. Molecular epidemiology of rotavirus A strains pre- and post-vaccine (Rotarix(R)) introduction in Mozambique, 2012-2019: Emergence of Genotypes G3P[4] and G3P[8]. Pathogens 2020, 9, 671. [Google Scholar] [CrossRef]
- Magagula, N.B.; Esona, M.D.; Nyaga, M.M.; Stucker, K.M.; Halpin, R.A.; Stockwell, T.B.; Seheri, M.L.; Steele, A.D.; Wentworth, D.E.; Mphahlele, M.J. Whole genome analyses of G1P[8] rotavirus strains from vaccinated and non-vaccinated South African children presenting with diarrhea. J. Med. Virol. 2015, 87, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, E.A.; Iturriza-Gómara, M.; Maes, P.; Patton, J.T. Full genome-based classification of rotaviruses reveals a common origin between human Wa-Like and porcine rotavirus strains and human DS-1-like and bovine rotavirus strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, P.; Aswath, K.; Collins, N.; Ahmed, T.; Olórtegui, M.P.; Kosek, M.; Cebelinski, E.; Cooper, P.J.; Bucardo, F.; Lopez, M.R. Near-complete genome sequences of several new norovirus genogroup II genotypes. Genome Announc. 2018, 6, e00007-18. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.-C.; Zhang, X.-F.; Xia, M.; Tan, M.; Quigley, C.; Lei, W.; Fang, H.; Zhong, W.; Lee, B.; Pang, X. Antigenic relatedness of norovirus GII. 4 variants determined by human challenge sera. PLoS ONE 2015, 10, e0124945. [Google Scholar] [CrossRef] [PubMed]
- Kambhampati, A.; Payne, D.C.; Costantini, V.; Lopman, B.A. Host genetic susceptibility to enteric viruses: A systematic review and metaanalysis. Clin. Infect. Dis. 2015, 62, 11–18. [Google Scholar] [CrossRef]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.-W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef]
- Mabasa, V.; Meno, K.; Taylor, M.; Mans, J. Environmental surveillance for noroviruses in selected South African wastewaters 2015–2016: Emergence of the novel GII. 17. Food Environ. Virol. 2018, 10, 16–28. [Google Scholar] [CrossRef]
- Page, N.; Groome, M.J.; Murray, T.; Nadan, S.; Netshikweta, R.; Keddy, K.H.; Poonsamy, B.; Moyes, J.; Walaza, S.; Kahn, K. Sapovirus prevalence in children less than five years of age hospitalised for diarrhoeal disease in South Africa, 2009–2013. J. Clin. Virol. 2016, 78, 82–88. [Google Scholar] [CrossRef]
- Murray, T.Y.; Nadan, S.; Page, N.A.; Taylor, M.B. Diverse sapovirus genotypes identified in children hospitalised with gastroenteritis in selected regions of South Africa. J. Clin. Virol. 2016, 76, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Mwenda, J.M.; Tate, J.E.; Parashar, U.D.; Mihigo, R.; Agocs, M.; Serhan, F.; Nshimirimana, D. African rotavirus surveillance network: A brief overview. Pediatr. Infect. Dis. J. 2014, 33 (Suppl. 1), S6–S8. [Google Scholar] [CrossRef] [PubMed]
- Nordgren, J.; Sharma, S.; Bucardo, F.; Nasir, W.; Günaydın, G.; Ouermi, D.; Nitiema, L.W.; Becker-Dreps, S.; Simpore, J.; Hammarström, L. Both Lewis and secretor status mediate susceptibility to rotavirus infections in a rotavirus genotype–dependent manner. Clin. Infect. Dis. 2014, 59, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Nordgren, J.; Svensson, L. Genetic susceptibility to human norovirus infection: An update. Viruses 2019, 11, 226. [Google Scholar] [CrossRef]
- Loureiro Tonini, M.A.; Pires Goncalves Barreira, D.M.; Bueno de Freitas Santolin, L.; Bondi Volpini, L.P.; Gagliardi Leite, J.P.; Le Moullac-Vaidye, B.; Le Pendu, J.; Cruz Spano, L. FUT2, Secretor Status and FUT3 Polymorphisms of Children with Acute Diarrhea Infected with Rotavirus and Norovirus in Brazil. Viruses 2020, 12, 1084. [Google Scholar] [CrossRef]
- Cannon, J.L.; Lopman, B.A.; Payne, D.C.; Vinjé, J. Birth cohort studies assessing norovirus infection and immunity in young children: A review. Clin. Infect. Dis. 2019, 69, 357–365. [Google Scholar] [CrossRef]
- Matussek, A.; Dienus, O.; Djeneba, O.; Simpore, J.; Nitiema, L.; Nordgren, J. Molecular characterization and genetic susceptibility of sapovirus in children with diarrhea in Burkina Faso. Infect. Genet. Evol. 2015, 32, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Wang, Q.; Katayama, K.; Saif, L.J. Comprehensive review of human sapoviruses. Clin. Microbiol. Rev. 2015, 28, 32–53. [Google Scholar] [CrossRef]
- Carlsson, B.; Kindberg, E.; Buesa, J.; Rydell, G.E.; Lidon, M.F.; Montava, R.; Mallouh, R.A.; Grahn, A.; Rodríguez-Díaz, J.; Bellido, J. The G428A nonsense mutation in FUT2 provides strong but not absolute protection against symptomatic GII. 4 Norovirus infection. PLoS ONE 2009, 4, e5593. [Google Scholar] [CrossRef]
- Frenck, R.; Bernstein, D.I.; Xia, M.; Huang, P.; Zhong, W.; Parker, S.; Dickey, M.; McNeal, M.; Jiang, X. Predicting susceptibility to norovirus GII. 4 by use of a challenge model involving humans. J. Infect. Dis. 2012, 206, 1386–1393. [Google Scholar] [CrossRef]
- Jin, M.; He, Y.; Li, H.; Huang, P.; Zhong, W.; Yang, H.; Zhang, H.; Tan, M.; Duan, Z.-J. Two gastroenteritis outbreaks caused by GII Noroviruses: Host susceptibility and HBGA phenotypes. PLoS ONE 2013, 8, e58605. [Google Scholar] [CrossRef]
- Liu, P.; Wang, X.; Lee, J.-C.; Teunis, P.; Hu, S.; Paradise, H.T.; Moe, C. Genetic susceptibility to norovirus GII. 3 and GII. 4 infections in Chinese pediatric diarrheal children. Pediatr. Infect. Dis. J. 2014, 33, e305. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Sano, D.; Suenaga, A.; Yoshimura, T.; Fuzawa, M.; Nakagomi, T.; Nakagomi, O.; Okabe, S. Histo-blood group antigen-like substances of human enteric bacteria as specific adsorbents for human noroviruses. J. Virol. 2013, 87, 9441–9451. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, P.; Tan, M.; Liu, Y.; Biesiada, J.; Meller, J.; Castello, A.A.; Jiang, B.; Jiang, X. Rotavirus VP8*: Phylogeny, host range, and interaction with histo-blood group antigens. J. Virol. 2012, 86, 9899–9910. [Google Scholar] [CrossRef] [PubMed]
- Pollock, L.; Bennett, A.; Jere, K.C.; Dube, Q.; Mandolo, J.; Bar-Zeev, N.; Heyderman, R.S.; Cunliffe, N.A.; Iturriza-Gomara, M. Nonsecretor histo–blood group antigen phenotype is associated with reduced risk of clinical rotavirus vaccine failure in Malawian infants. Clin. Infect. Dis. 2018, 69, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Seheri, M.; Nemarude, L.; Peenze, I.; Netshifhefhe, L.; Nyaga, M.M.; Ngobeni, H.G.; Maphalala, G.; Maake, L.L.; Steele, A.D.; Mwenda, J.M.; et al. Update of rotavirus strains circulating in Africa from 2007 through 2011. Pediatr. Infect. Dis. J. 2014, 33 (Suppl. 1), S76–S84. [Google Scholar] [CrossRef] [PubMed]
- Todd, S.; Page, N.A.; Steele, A.D.; Peenze, I.; Cunliffe, N.A. Rotavirus strain types circulating in Africa: Review of studies published during 1997–2006. J. Infect. Dis. 2010, 202 (Suppl. 1), S34–S42. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Cohort (n = 205) | Norovirus (+) Cohort (n = 32) | Rotavirus (+) Cohort (n = 46) | Virus (−) Cohort (n = 109) | Features Tested for Association | p-Value |
|---|---|---|---|---|---|---|
| Demographic Characteristics | ||||||
| Median age (in months) at entry (min-max) | 10 (0.5−64) | 10 (0.5−46) | 8.5 (0.5−48) | 14 (1−64) | a Age/Virus infection | 0.0002 chi2 = 13.845 |
| 0–<12, n (%) | 115 (56) | 21 (66) | 32 (70) | 48 (44) | ||
| 12–<24, n (%) | 51 (25) | 9 (28) | 10 (22) | 29 (27) | ||
| 24–<48, n (%) | 28 (14) | 2 (6) | 3 (7) | 22 (20) | ||
| 48–60, n (%) | 12 (16) | 0 | 1 (1) | 10 (9) | ||
| Gender, n (%) | ||||||
| Male | 122 (60) | 17 (53) | 26 (57) | 64 (59) | ||
| Female | 83 (40) | 15 (47) | 20 (43) | 45 (41) | ||
| Race, n (%) | ||||||
| White | 1 (0.5) | 1 (3) | 1 (2) | 0 | ||
| Black | 201 (98) | 31 (97) | 45 (98) | 106 (97) | ||
| Coloured | 2 (1) | 0 | 0 | 2 (2) | ||
| Asian | 1 (0.5) | 0 | 0 | 1 (1) | ||
| Environmental Features | ||||||
| Water source | a Water/ | |||||
| Indoor tap, n (%) | 79 (39) | 15 (47) | 24 (52) | 36 (33) | Virus | 0.1993 |
| Other, n (%) | 126 (61) | 17 (53) | 22 (48) | 71 (65) | infection | chi2 = 1.647 |
| Sanitation type | a Sanitation/ | |||||
| Flush toilet, n (%) | 126 (61) | 16 (50) | 34 (74) | 65 (60) | Virus | 0.09873 |
| Other, n (%) | 79 (39) | 16 (50) | 12 (26) | 44 (40) | infection | chi2 = 0.026 |
| Clinical characteristics | ||||||
| No. of days with diarrhoea, median (IQR) | 3 (3) | 3 (2) | 3 (3) | 3 (3) | ||
| 1–4 days, n (%) | 145 (71) | 25 (78) | 35 (75) | 36 (33) | ||
| 5 days, n (%) | 21 (10) | 2 (6) | 3 (7) | 25 (23) | ||
| ≥6 days, n (%) | 39 (19) | 5 (16) | 8 (17) | 48 (44) | ||
| Maximum no. of loose/watery stools in 24 h period, median (IQR) | 5 (3) | 5 (3.75) | 5 (3) | 5 (2.5) | ||
| 1–4, n (%) | 70 (35) | 14 (43) | 14 (30) | 36 (33) | ||
| 5, n (%) | 46 (22) | 5 (16) | 12 (26) | 25 (23) | ||
| ≥6, n (%) | 89 (43) | 13 (41) | 20 (44) | 48 (44) | ||
| Vomiting, n (%) | 118 (58) | 22 (69) | 28 (61) | 57 (28) | * Vomiting/adenovirus | 0.788 |
| No. of days with vomiting | norovirus | 0.242 | ||||
| 1 day, n (%) | 64 (54) | 7 (32) | 16 (57) | 36 (63) | rotavirus | 0.609 |
| 2 days, n (%) | 20 (17) | 4 (18) | 8 (29) | 6 (11) | sapovirus | 1 |
| ≥3 days, n (%) | 34 (29) | 11 (50) | 4 (14) | 15 (26) | ||
| Maximum no. vomiting episodes in 24 h | ||||||
| Median (IQR) | 1 (2) | 2 (2) | 1 (1) | 1 (2) | ||
| 1 episode, n (%) | 61 (52) | 7 (32) | 16 (57) | 30 (53) | ||
| 2−4 episodes, n (%) | 45 (38) | 12 (54) | 9 (32) | 21 (37) | ||
| ≥5 episodes, n (%) | 12 (10) | 3 (14) | 3 (11) | 6 (10) | ||
| Fever in previous 48 h, n (%) ‡ | 76 (37) | 5 (16) | 3 (7) | 17 (16) | * Fever/adenovirus | 1 |
| ≤37.0 °C | 171 (83) | 26 (81) | 43 (93) | 88 (81) | norovirus | 0.591 |
| 37.1–38.4 °C | 16 (8) | 3 (9.5) | 2 (4) | 8 (7) | rotavirus | 0.099 |
| 38.5–38.9 °C | 14 (7) | 3 (9.5) | 1 (2) | 9 (8) | sapovirus | 0.139 |
| ≥39.0 °C | 4 (2) | 0 | 0 | 4 (4) | ||
| Dehydration score | ||||||
| No dehydration (0), n (%) | 20 (10) | 3 (9) | 5 (11) | 10 (9) | ||
| Mild (1–5), n (%) | 15 (7) | 1 (3) | 3 (7) | 9 (8) | ||
| Moderate–severe (≥6), n (%) | 170 (83) | 28 (88) | 38 (83) | 90 (83) | ||
| Diarrhoea type | ||||||
| Watery, n (%) | 175 (85) | 26 (81) | 43 (93) | 89 (82) | ||
| Dysentery, n (%) | 30 (15) | 6 (19) | 3 (7) | 20 (18) | ||
| HIV status | a HIV | |||||
| Uninfected, unexposed, n (%) | 134 (65) | 22 (69) | 33 (72) | 70 (64) | exposure/ | |
| Uninfected, exposed, n (%) | 61 (30) | 8 (25) | 12 (26) | 33 (30) | adenovirus | 0.6227 chi2 = 0.947 |
| Infected, n (%) | 10 (5) | 2 (6) | 1 (2) | 6 (6) | norovirus | 0.9062 |
| chi2 = 0.197 | ||||||
| rotavirus | 0.6727 | |||||
| chi2 = 0.793 | ||||||
| sapovirus | 0.9229 | |||||
| chi2 = 0.160 | ||||||
| Baseline Vesikari score, mean (sd) | 10.4 (2.7) | 11.1 (2.5) | 10.1 (2.2) | 10.3 (2.9) | a HIV | |
| Mild (˂7), n (%) | 16 (8) | 1 (3) | 2 (4) | 12 (11) | exposure/ | |
| Moderate (7–10), n (%) | 88 (43) | 11 (34) | 24 (52) | 45 (41) | Severity of | 0.072 |
| Severe (11–20), n (%) | 101 (49) | 20 (63) | 20 (44) | 52 (48) | symptoms | chi2 = 8.5961 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossouw, E.; Brauer, M.; Meyer, P.; du Plessis, N.M.; Avenant, T.; Mans, J. Virus Etiology, Diversity and Clinical Characteristics in South African Children Hospitalised with Gastroenteritis. Viruses 2021, 13, 215. https://doi.org/10.3390/v13020215
Rossouw E, Brauer M, Meyer P, du Plessis NM, Avenant T, Mans J. Virus Etiology, Diversity and Clinical Characteristics in South African Children Hospitalised with Gastroenteritis. Viruses. 2021; 13(2):215. https://doi.org/10.3390/v13020215
Chicago/Turabian StyleRossouw, Esmari, Marieke Brauer, Pieter Meyer, Nicolette M. du Plessis, Theunis Avenant, and Janet Mans. 2021. "Virus Etiology, Diversity and Clinical Characteristics in South African Children Hospitalised with Gastroenteritis" Viruses 13, no. 2: 215. https://doi.org/10.3390/v13020215
APA StyleRossouw, E., Brauer, M., Meyer, P., du Plessis, N. M., Avenant, T., & Mans, J. (2021). Virus Etiology, Diversity and Clinical Characteristics in South African Children Hospitalised with Gastroenteritis. Viruses, 13(2), 215. https://doi.org/10.3390/v13020215