
Influenza A Virus Hemagglutinin and Other Pathogen Glycoprotein Interactions with NK Cell Natural Cytotoxicity Receptors NKp46, NKp44, and NKp30



Abstract

1. Introduction
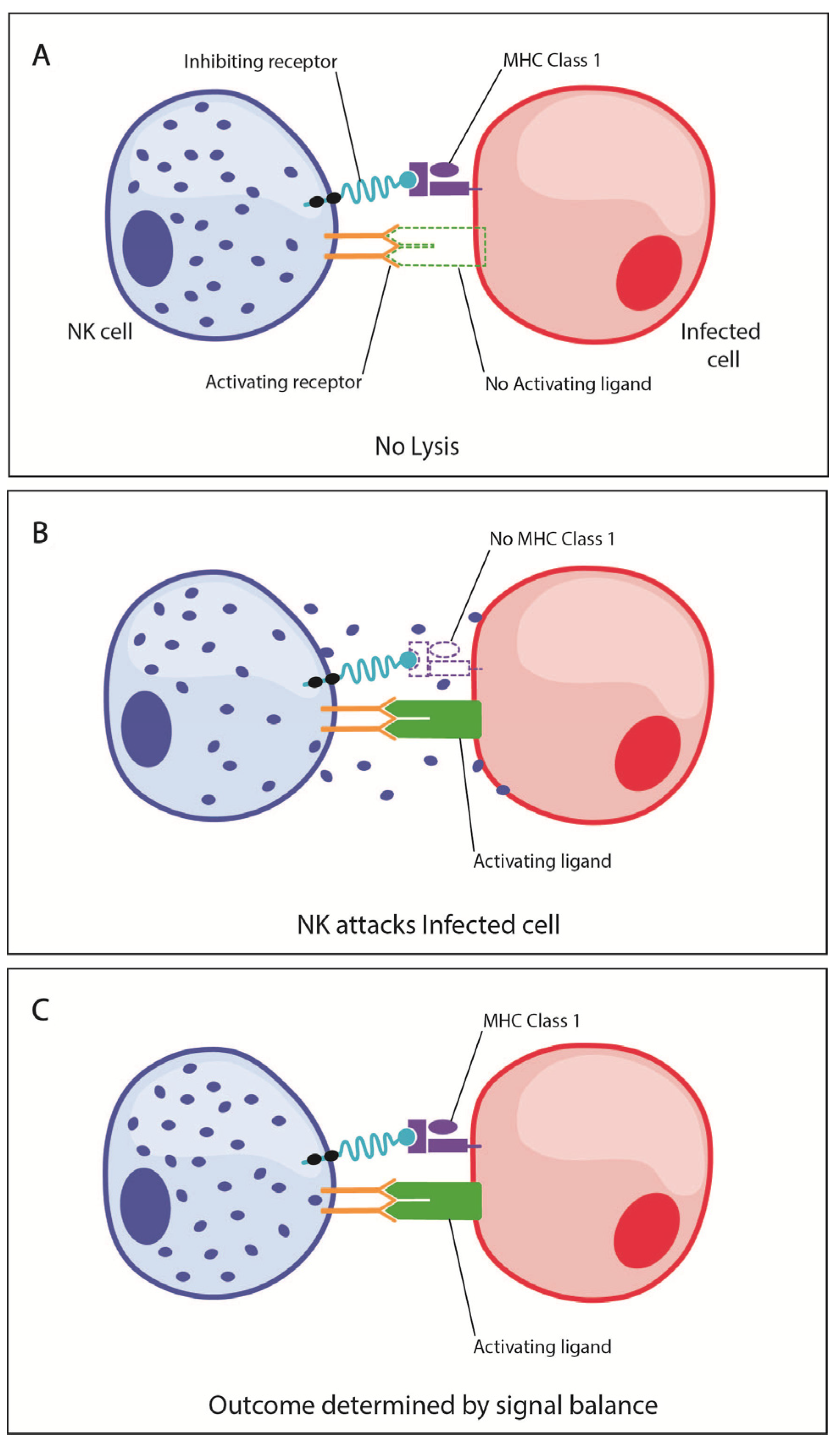
2. NK Cell Functional Biology
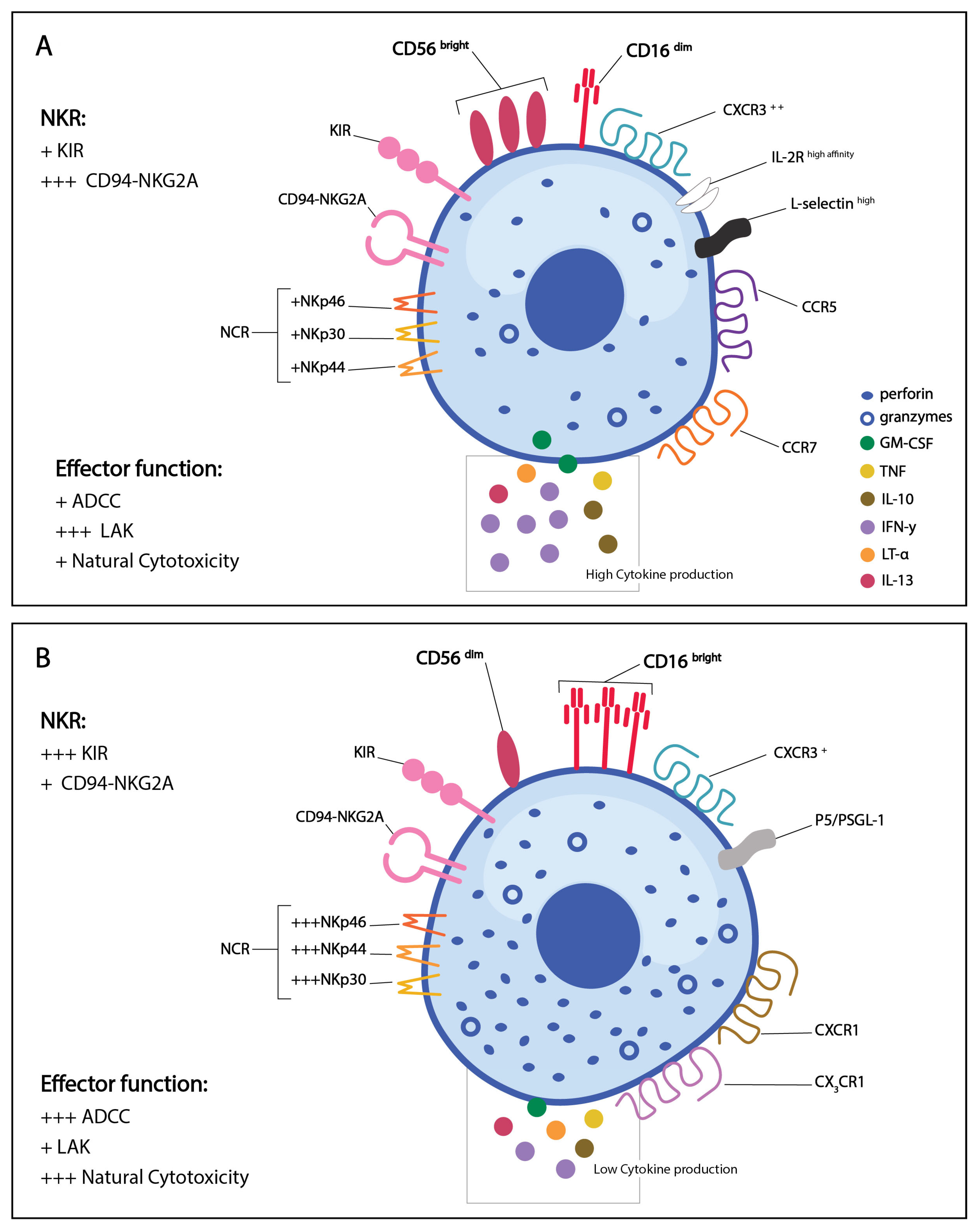
3. NK Cell Phenotypic Subsets
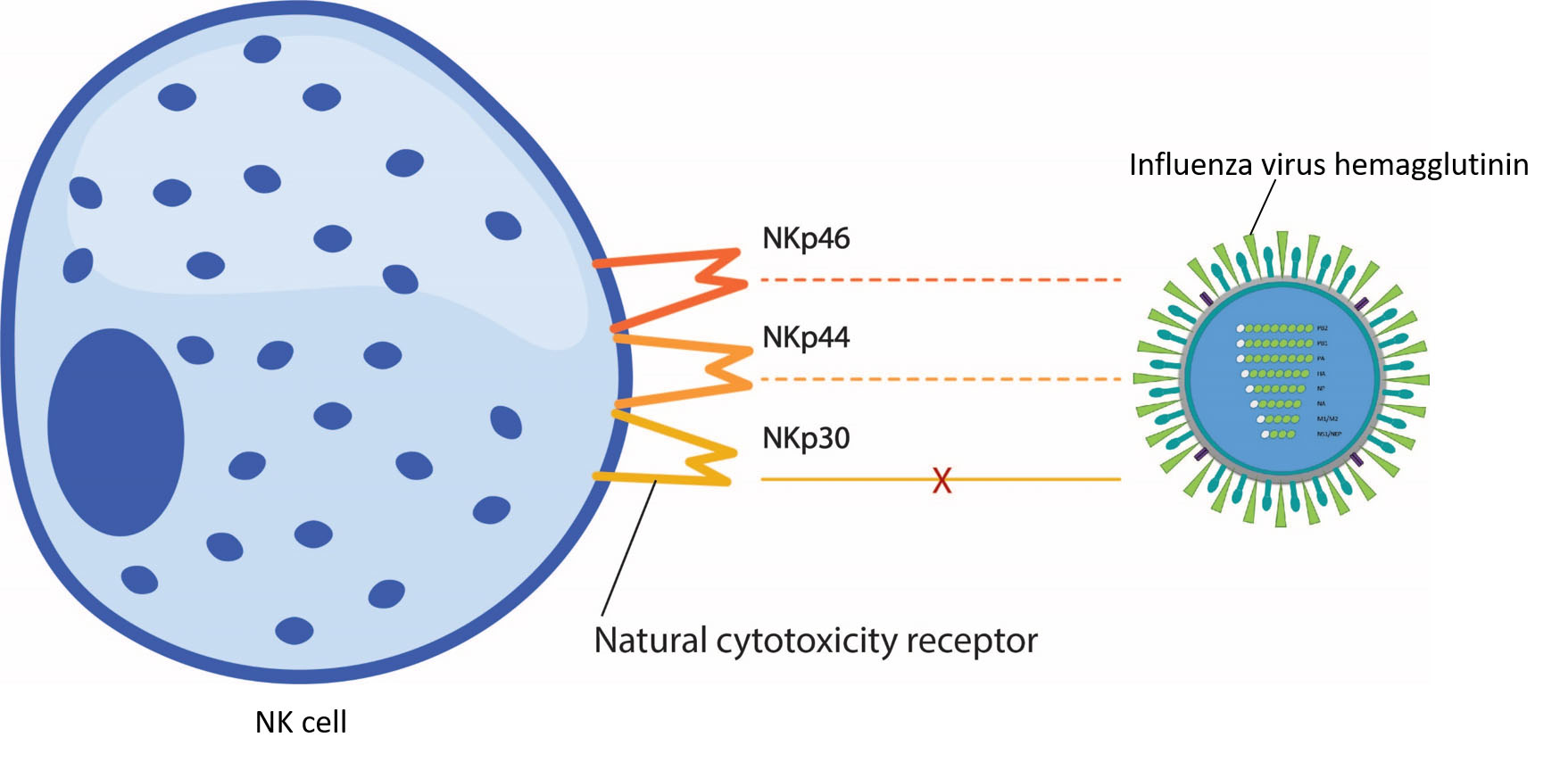
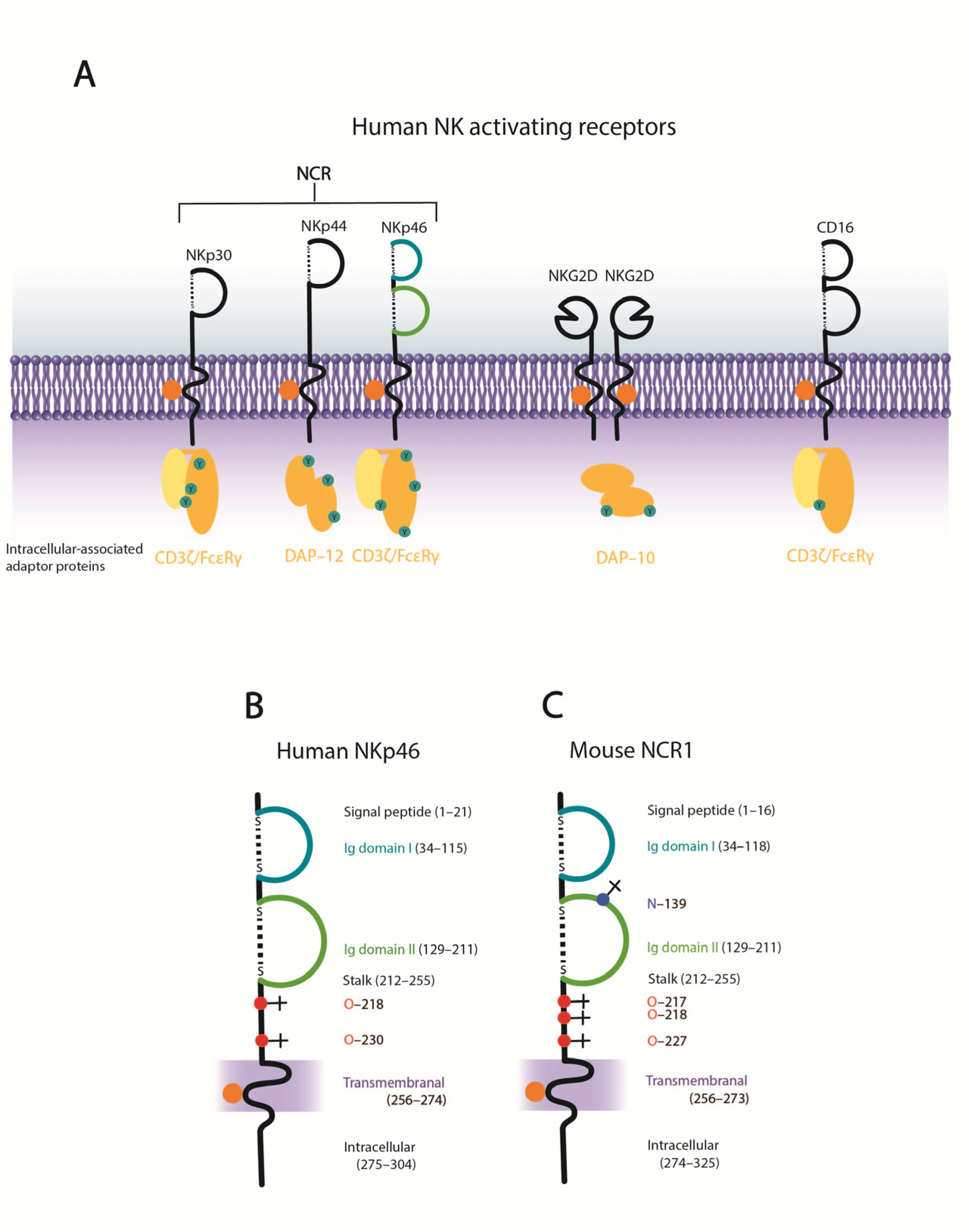
4. NK Cell Activating Receptors–the Natural Cytotoxicity Receptors
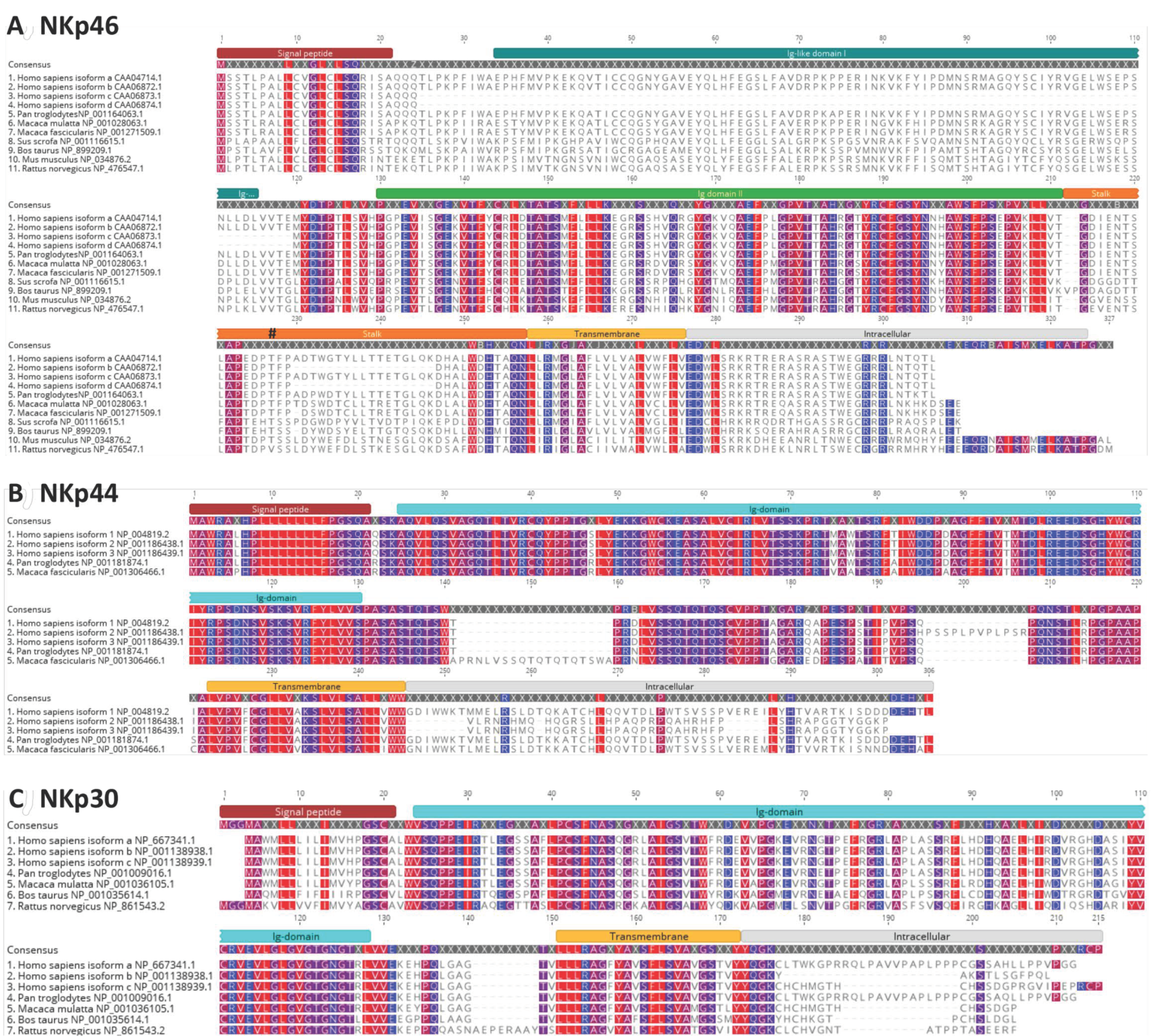
4.1. NKp46
4.2. NKp44
4.3. NKp30
5. Glycobiology of Human NK Cell NCRs
5.1. NKp46
5.2. NKp44
5.3. NKp30
6. Role of NK Cells in Influenza Infection
7. Functional Biology of Influenza a Virus Hemagglutinin
8. Influenza Haemagglutinin Interactions with NK Cell NCRS
8.1. Inlfuenza A Virus HA Interaction with NKp46
8.2. Inlfuenza A Virus HA Interaction with NKp44 and NKp30
8.3. Identification of NCR Domain that Engages with Influenza A Virus HA
8.4. Sialylated Glycans Are Required for Influenza A Virus HA to Interact with NCRs
8.5. Neuraminidase Treatment of NCRs Abrogates HA Binding
8.6. Sialic Acid Linkage and NCR Recognition
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Greenberg, A.H.; Hudson, L.; Shen, L.; Roitt, I.M. Antibody-dependent cell-mediated cytotoxicity due to a “null” lymphoid cell. Nat. New Biol. 1973, 242, 111–113. [Google Scholar] [CrossRef]
- Harding, B.; Pudifin, D.J.; Gotch, F.; Maclennan, I.C.M. Cytotoxic lymphocytes from rats depleted of thymus processed cells. Nat. New Biol. 1971, 232, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Van Boxel, J.A.; Stobo, J.D.; Paul, W.E.; Green, I. Antibody-dependent lymphoid cell-mediated cytotoxicity: No requirement for thymus-derived lymphocytes. Science 1972, 175, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Shifrin, N.; Raulet, D.H.; Ardolino, M. NK cell self tolerance, responsiveness and missing self recognition. Semin. Immunol. 2014, 26, 138–144. [Google Scholar] [CrossRef]
- Vivier, E. What is natural in natural killer cells? Immunol. Lett. 2006, 107, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cheent, K.; Khakoo, S.I. Natural killer cells: Integrating diversity with function. Immunology 2009, 126, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Maeda, K.; Nakanishi, N.; Rogers, B.L.; Haser, W.G.; Shitara, K.; Yoshida, H.; Takagaki, Y.; Augustin, A.A.; Tonegawa, S. Expression of the T-cell receptor gamma-chain gene products on the surface of peripheral T cells and T-cell blasts generated by allogeneic mixed lymphocyte reaction. Proc. Natl. Acad. Sci. USA 1987, 84, 6536–6540. [Google Scholar] [CrossRef] [PubMed]
- Sakano, H.; Kurosawa, Y.; Weigert, M.; Tonegawa, S. Identification and nucleotide sequence of a diversity DNA segment (D) of immunoglobulin heavy-chain genes. Nature 1981, 290, 562–565. [Google Scholar] [CrossRef]
- Hozumi, N.; Tonegawa, S. Evidence for somatic rearrangement of immunoglobulin genes coding for variable and constant regions. Proc. Natl. Acad. Sci. USA 1976, 73, 3628–3632. [Google Scholar] [CrossRef]
- Lanier, L.L. NK cell recognition. Annu. Rev. Immunol. 2005, 23, 225–274. [Google Scholar] [CrossRef] [PubMed]
- Biassoni, R.; Cantoni, C.; Pende, D.; Sivori, S.; Parolini, S.; Vitale, M.; Bottino, C.; Moretta, A. Human natural killer cell receptors and co-receptors. Immunol. Rev. 2001, 181, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Glas, R.; Franksson, L.; Une, C.; Eloranta, M.-L.; Öhlén, C.; Örn, A.; Kärre, K. Recruitment and activation of natural killer (NK) cells in vivo determined by the target cell phenotype: An adaptive component of NK cell–mediated responses. J. Exp. Med. 2000, 191, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Spits, H.; Blom, B.; Jaleco, A.-C.; Weijer, K.; Verschuren, M.C.M.; Dongen, J.J.M.; Heemskerk, M.H.M.; Res, P.C.M. Early stages in the development of human T, natural killer and thymic dendritic cells. Immunol. Rev. 1998, 165, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Herberman, R.B.; Nunn, M.E.; Lavrin, D.H. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. I. Distribution of reactivity and specificity. Int. J. Cancer 1975, 16, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Stegmann, K.; Riley, E. Activation of Natural Killer cells during microbial infections. Front. Immunol. 2012, 2. [Google Scholar] [CrossRef]
- Korbel, D.S.; Finney, O.C.; Riley, E.M. Natural killer cells and innate immunity to protozoan pathogens. Int. J. Parasitol. 2004, 34, 1517–1528. [Google Scholar] [CrossRef]
- Lodoen, M.B.; Lanier, L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006, 18, 391–398. [Google Scholar] [CrossRef]
- Schmidt, S.; Tramsen, L.; Lehrnbecher, T. Natural killer cells in antifungal immunity. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Biron, C.A.; Byron, K.S.; Sullivan, J.L. Severe herpesvirus infections in an adolescent without natural killer cells. N. Engl. J. Med. 1989, 320, 1731–1735. [Google Scholar] [CrossRef]
- Gazit, R.; Gruda, R.; Elboim, M.; Arnon, T.I.; Katz, G.; Achdout, H.; Hanna, J.; Qimron, U.; Landau, G.; Greenbaum, E.; et al. Lethal influenza infection in the absence of the natural killer cell receptor gene NCR1. Nat. Immunol. 2006, 7, 517. [Google Scholar] [CrossRef] [PubMed]
- Glasner, A.; Zurunic, A.; Meningher, T.; Lenac Rovis, T.; Tsukerman, P.; Bar-On, Y.; Yamin, R.; Meyers, A.F.A.; Mandeboim, M.; Jonjic, S.; et al. Elucidating the mechanisms of influenza virus recognition by NCR1. PLoS ONE 2012, 7, e36837. [Google Scholar] [CrossRef]
- Bar-On, Y.; Glasner, A.; Meningher, T.; Achdout, H.; Gur, C.; Lankry, D.; Vitenshtein, A.; Meyers, A.F.; Mandelboim, M.; Mandelboim, O. Neuraminidase-mediated, NKp46-dependent immune-evasion mechanism of influenza viruses. Cell Rep. 2013, 3, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Della Chiesa, M.; Marcenaro, E.; Sivori, S.; Carlomagno, S.; Pesce, S.; Moretta, A. Human NK cell response to pathogens. Semin. Immunol. 2014, 26, 152–160. [Google Scholar] [CrossRef]
- Frank, K.; Paust, S. Dynamic Natural Killer Cell and T Cell Responses to Influenza Infection. Front. Cell. Infect. Microbiol. 2020, 10, 425. [Google Scholar] [CrossRef] [PubMed]
- Joyce, M.G.; Sun, P.D. The structural basis of ligand recognition by natural killer cell receptors. J. Biomed. Biotechnol. 2011, 2011, 203628. [Google Scholar] [CrossRef]
- Mandelboim, O.; Lieberman, N.; Lev, M.; Paul, L.; Arnon, T.I.; Bushkin, Y.; Davis, D.M.; Strominger, J.L.; Yewdell, J.W.; Porgador, A. Recognition of haemagglutinins on virus-infected cells by NKp46 activates lysis by human NK cells. Nature 2001, 409, 1055. [Google Scholar] [CrossRef]
- Arnon, T.I.; Achdout, H.; Lieberman, N.; Gazit, R.; Gonen-Gross, T.; Katz, G.; Bar-Ilan, A.; Bloushtain, N.; Lev, M.; Joseph, A.; et al. The mechanisms controlling the recognition of tumor- and virus-infected cells by NKp46. Blood 2004, 103, 664–672. [Google Scholar] [CrossRef]
- Ho, J.W.; Hershkovitz, O.; Peiris, M.; Zilka, A.; Bar-Ilan, A.; Nal, B.; Chu, K.; Kudelko, M.; Kam, Y.W.; Achdout, H.; et al. H5-type influenza virus hemagglutinin is functionally recognized by the natural killer-activating receptor NKp44. J. Virol. 2008, 82, 2028–2032. [Google Scholar] [CrossRef]
- Mendelson, M.; Tekoah, Y.; Zilka, A.; Gershoni-Yahalom, O.; Gazit, R.; Achdout, H.; Bovin, N.V.; Meningher, T.; Mandelboim, M.; Mandelboim, O.; et al. NKp46 O-Glycan Sequences That Are Involved in the Interaction with Hemagglutinin Type 1 of Influenza Virus. J. Virol. 2010, 84, 3789–3797. [Google Scholar] [CrossRef] [PubMed]
- Du, N.; Zhou, J.; Lin, X.; Zhang, Y.; Yang, X.; Wang, Y.; Shu, Y. Differential activation of NK cells by influenza A pseudotype H5N1 and 1918 and 2009 pandemic H1N1 viruses. J. Virol. 2010, 84, 7822–7831. [Google Scholar] [CrossRef] [PubMed]
- Chaushu, S.; Wilensky, A.; Gur, C.; Shapira, L.; Elboim, M.; Halftek, G.; Polak, D.; Achdout, H.; Bachrach, G.; Mandelboim, O. Direct recognition of Fusobacterium nucleatum by the NK cell natural cytotoxicity receptor NKp46 aggravates periodontal disease. PLoS Pathog. 2012, 8, e1002601. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.; Seidel, E.; Tsukerman, P.; Mandelboim, M.; Mandelboim, O. Influenza virus uses its neuraminidase protein to evade the recognition of two activating NK cell receptors. J. Infect. Dis. 2014, 210, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Glasner, A.; Roth, Z.; Varvak, A.; Miletic, A.; Isaacson, B.; Bar-On, Y.; Jonjic, S.; Khalaila, I.; Mandelboim, O. Identification of putative novel O-glycosylations in the NK killer receptor NCR1 essential for its activity. Cell Discov. 2015, 1, 15036. [Google Scholar] [CrossRef]
- Duev-Cohen, A.; Bar-On, Y.; Glasner, A.; Berhani, O.; Ophir, Y.; Levi-Schaffer, F.; Mandelboim, M.; Mandelboim, O. The human 2B4 and NTB-A receptors bind the influenza viral hemagglutinin and co-stimulate NK cell cytotoxicity. Oncotarget 2016, 7, 13093–13105. [Google Scholar] [CrossRef]
- McQuaid, S.; Loughran, S.; Power, P.; Maguire, P.; Walls, D.; Cusi, M.G.; Orvell, C.; Johnson, P. Haemagglutinin-neuraminidase from HPIV3 mediates human NK regulation of T cell proliferation via NKp44 and NKp46. J. Gen. Virol. 2018, 99, 763–767. [Google Scholar] [CrossRef]
- Achdout, H.; Arnon, T.I.; Markel, G.; Gonen-Gross, T.; Katz, G.; Lieberman, N.; Gazit, R.; Joseph, A.; Kedar, E.; Mandelboim, O. Enhanced recognition of human NK receptors after influenza virus infection. J. Immunol. 2003, 171, 915–923. [Google Scholar] [CrossRef]
- Arnon, T.I.; Lev, M.; Katz, G.; Chernobrov, Y.; Porgador, A.; Mandelboim, O. Recognition of viral hemagglutinins by NKp44 but not by NKp30. Eur. J. Immunol. 2001, 31, 2680–2689. [Google Scholar] [CrossRef]
- Jarahian, M.; Watzl, C.; Fournier, P.; Arnold, A.; Djandji, D.; Zahedi, S.; Cerwenka, A.; Paschen, A.; Schirrmacher, V.; Momburg, F. Activation of natural killer cells by Newcastle disease virus hemagglutinin-neuraminidase. J. Virol. 2009, 83, 8108–8121. [Google Scholar] [CrossRef]
- Diab, M.; Glasner, A.; Isaacson, B.; Bar-On, Y.; Drori, Y.; Yamin, R.; Duev-Cohen, A.; Danziger, O.; Zamostiano, R.; Mandelboim, M.; et al. NK-cell receptors NKp46 and NCR1 control human metapneumovirus infection. Eur. J. Immunol. 2017, 47, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Arnon, T.I.; Achdout, H.; Levi, O.; Markel, G.; Saleh, N.; Katz, G.; Gazit, R.; Gonen-Gross, T.; Hanna, J.; Nahari, E.; et al. Inhibition of the NKp30 activating receptor by pp65 of human cytomegalovirus. Nat. Immunol. 2005, 6, 515. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.E.; Howard, K.; Valés Gómez, M.; Reyburn, H.T. Expression of ICP0 is sufficient to trigger natural killer cell recognition of herpes simplex virus—infected cells by natural cytotoxicity receptors. J. Infect. Dis. 2007, 195, 1160–1168. [Google Scholar] [CrossRef] [PubMed]
- Madrid, A.S.; Ganem, D. Kaposi’s sarcoma-associated herpesvirus ORF54/dUTPase downregulates a ligand for the NK activating receptor NKp44. J. Virol. 2012, 86, 8693–8704. [Google Scholar] [CrossRef]
- Chisholm, S.E.; Reyburn, H.T. Recognition of vaccinia virus-infected cells by human natural killer cells depends on natural cytotoxicity receptors. J. Virol. 2006, 80, 2225–2233. [Google Scholar] [CrossRef]
- Jarahian, M.; Fiedler, M.; Cohnen, A.; Djandji, D.; Hämmerling, G.J.; Gati, C.; Cerwenka, A.; Turner, P.C.; Moyer, R.W.; Watzl, C.; et al. Modulation of NKp30- and NKp46-mediated natural killer cell responses by poxviral hemagglutinin. PLoS Pathog. 2011, 7, e1002195. [Google Scholar] [CrossRef]
- Hershkovitz, O.; Rosental, B.; Rosenberg, L.A.; Navarro-Sanchez, M.E.; Jivov, S.; Zilka, A.; Gershoni-Yahalom, O.; Brient-Litzler, E.; Bedouelle, H.; Ho, J.W.; et al. NKp44 receptor mediates interaction of the envelope glycoproteins from the West Nile and Dengue viruses with NK cells. J. Immunol. 2009, 183, 2610–2621. [Google Scholar] [CrossRef]
- Vankayalapati, R.; Wizel, B.; Weis, S.E.; Safi, H.; Lakey, D.L.; Mandelboim, O.; Samten, B.; Porgador, A.; Barnes, P.F. The NKp46 receptor contributes to NK cell lysis of mononuclear phagocytes infected with an intracellular bacterium. J. Immunol. 2002, 168, 3451–3457. [Google Scholar] [CrossRef]
- Garg, A.; Barnes, P.F.; Porgador, A.; Roy, S.; Wu, S.; Nanda, J.S.; Griffith, D.E.; Girard, W.M.; Rawal, N.; Shetty, S.; et al. Vimentin expressed on Mycobacterium tuberculosis-infected human monocytes is involved in binding to the NKp46 receptor. J. Immunol. 2006, 177, 6192–6198. [Google Scholar] [CrossRef]
- Esin, S.; Batoni, G.; Counoupas, C.; Stringaro, A.; Brancatisano, F.L.; Colone, M.; Maisetta, G.; Florio, W.; Arancia, G.; Campa, M. Direct binding of human NK cell natural cytotoxicity receptor NKp44 to the surfaces of Mycobacteria and other bacteria. Infect. Immun. 2008, 76, 1719–1727. [Google Scholar] [CrossRef]
- Mavoungou, E.; Held, J.; Mewono, L.; Kremsner, P.G. A duffy binding—like domain is involved in the NKp30-mediated recognition of Plasmodium falciparum—Parasitized erythrocytes by natural killer cells. J. Infect. Dis. 2007, 195, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Rees, R.C.; Oxford, J. Modulation of human natural killer cytotoxicity by influenza virus and its subunit protein. Immunol. 1984, 52, 687–695. [Google Scholar]
- Arora, D.J.; Houde, M.; Justewicz, D.M.; Mandeville, R. In vitro enhancement of human natural cell-mediated cytotoxicity by purified influenza virus glycoproteins. J. Virol. 1984, 52, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Sivori, S.; Vitale, M.; Morelli, L.; Sanseverino, L.; Augugliaro, R.; Bottino, C.; Moretta, L.; Moretta, A. p46, a novel natural killer cell–specific surface molecule that mediates cell activation. J. Exp. Med. 1997, 186, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Bloushtain, N.; Qimron, U.; Bar-Ilan, A.; Hershkovitz, O.; Gazit, R.; Fima, E.; Korc, M.; Vlodavsky, I.; Bovin, N.V.; Porgador, A. Membrane-associated heparan sulfate proteoglycans are involved in the recognition of cellular targets by NKp30 and NKp46. J. Immunol. 2004, 173, 2392–2401. [Google Scholar] [CrossRef] [PubMed]
- Vieillard, V.; Strominger, J.L.; Debré, P. NK cytotoxicity against CD4+ T cells during HIV-1 infection: A gp41 peptide induces the expression of an NKp44 ligand. Proc. Natl. Acad. Sci. USA 2005, 102, 10981–10986. [Google Scholar] [CrossRef]
- Owen, R.E.; Yamada, E.; Thompson, C.I.; Phillipson, L.J.; Thompson, C.; Taylor, E.; Zambon, M.; Osborn, H.M.I.; Barclay, W.S.; Borrow, P. Alterations in receptor binding properties of recent human influenza H3N2 viruses are associated with reduced natural killer cell lysis of Infected cells. J. Virol. 2007, 81, 11170–11178. [Google Scholar] [CrossRef]
- Achdout, H.; Meningher, T.; Hirsh, S.; Glasner, A.; Bar-On, Y.; Gur, C.; Porgador, A.; Mendelson, M.; Mandelboim, M.; Mandelboim, O. Killing of Avian and Swine Influenza Virus by Natural Killer Cells. J. Virol. 2010, 84, 3993–4001. [Google Scholar] [CrossRef]
- Mao, H.; Tu, W.; Liu, Y.; Qin, G.; Zheng, J.; Chan, P.-L.; Lam, K.-T.; Peiris, J.S.M.; Lau, Y.-L. Inhibition of human natural killer cell activity by influenza virions and hemagglutinin. J. Virol. 2010, 84, 4148–4157. [Google Scholar] [CrossRef]
- Ito, K.; Higai, K.; Sakurai, M.; Shinoda, C.; Yanai, K.; Azuma, Y.; Matsumoto, K. Binding of natural cytotoxicity receptor NKp46 to sulfate- and α2,3-NeuAc-containing glycans and its mutagenesis. Biochem. Biophys. Res. Commun. 2011, 406, 377–382. [Google Scholar] [CrossRef]
- Ito, K.; Higai, K.; Shinoda, C.; Sakurai, M.; Yanai, K.; Azuma, Y.; Matsumoto, K. Unlike natural killer (NK) p30, natural cytotoxicity receptor NKp44 binds to multimeric α2,3-NeuNAc-containing N-glycans. Biol. Pharm. Bull. 2012, 35, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Vitenshtein, A.; Charpak-Amikam, Y.; Yamin, R.; Bauman, Y.; Isaacson, B.; Stein, N.; Berhani, O.; Dassa, L.; Gamliel, M.; Gur, C.; et al. NK cell recognition of Candida glabrata through binding of NKp46 and NCR1 to fungal ligands Epa1, Epa6, and Epa7. Cell Host Microbe 2016, 20, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Kumar, P.; Malarkannan, S. Evasion of natural killer cells by influenza virus. J. Leukoc. Biol. 2011, 89, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Ljunggren, H.-G.; Kärre, K. Experimental strategies and interpretations in the analysis of changes in MHC gene expression during tumour progression. Int. J. Immunogenet. 1986, 13, 141–151. [Google Scholar] [CrossRef]
- Bix, M.; Liao, N.-S.; Zijlstra, M.; Loring, J.; Jaenisch, R.; Raulet, D. Rejection of class I MHC-deficient haemopoietic cells by irradiated MHC-matched mice. Nature 1991, 349, 329. [Google Scholar] [CrossRef]
- Liao, N.; Bix, M.; Zijlstra, M.; Jaenisch, R.; Raulet, D. MHC class I deficiency: Susceptibility to natural killer (NK) cells and impaired NK activity. Science 1991, 253, 199–202. [Google Scholar] [CrossRef]
- Schuren, A.B.C.; Costa, A.I.; Wiertz, E.J.H.J. Recent advances in viral evasion of the MHC Class I processing pathway. Curr. Opin. Immunol. 2016, 40, 43–50. [Google Scholar] [CrossRef]
- Orr, M.T.; Lanier, L.L. Natural killer cell education and tolerance. Cell 2010, 142, 847–856. [Google Scholar] [CrossRef]
- Lanier, L.L.; Corliss, B.; Phillips, J.H. Arousal and inhibition of human NK cells. Immunol. Rev. 1997, 155, 145–154. [Google Scholar] [CrossRef]
- Di Vito, C.; Mikulak, J.; Mavilio, D. On the Way to Become a Natural Killer Cell. Front. Immunol. 2019, 10, 1812. [Google Scholar] [CrossRef]
- Thomas, L.M.; Peterson, M.E.; Long, E.O. Cutting edge: NK cell licensing modulates adhesion to target cells. J. Immunol. 2013, 191, 3981–3985. [Google Scholar] [CrossRef] [PubMed]
- Joncker, N.T.; Fernandez, N.C.; Treiner, E.; Vivier, E.; Raulet, D.H. NK cell responsiveness is tuned commensurate with the number of inhibitory receptors for self-MHC class I: The rheostat model. J. Immunol. 2009, 182, 4572–4580. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.; Salmon-Divon, M.; Johansson, M.H.; Pickman, Y.; Brodin, P.; Kärre, K.; Mehr, R.; Höglund, P. Probing natural killer cell education by Ly49 receptor expression analysis and computational modelling in single MHC class I mice. PLoS ONE 2009, 4, e6046. [Google Scholar] [CrossRef]
- Brodin, P.; Lakshmikanth, T.; Johansson, S.; Kärre, K.; Höglund, P. The strength of inhibitory input during education quantitatively tunes the functional responsiveness of individual natural killer cells. Blood 2009, 113, 2434–2441. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.C.; Lanier, L.L. Cutting edge: Viral infection breaks NK cell tolerance to “missing self”. J. Immunol. 2008, 181, 7453–7457. [Google Scholar] [CrossRef]
- Suzuki, N.; Suzuki, T.; Engleman, E.G. Evidence for the involvement of CD56 molecules in alloantigen-specific recognition by human natural-killer-cells. J. Exp. Med. 1991, 173, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Van Acker, H.H.; Capsomidis, A.; Smits, E.L.; Van Tendeloo, V.F. CD56 in the immune system: More than a marker for cytotoxicity? Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, G.; Thomas, D.; Lin, S.-L.; Goodman, K.; Morandi, B.; Muller, W.A.; Moretta, A.; Münz, C. The abundant NK cells in human secondary lymphoid tissues require activation to express killer cell Ig-like receptors and become cytolytic. J. Immunol. 2004, 172, 1455–1462. [Google Scholar] [CrossRef]
- Caligiuri, M.A.; Zmuidzinas, A.; Manley, T.J.; Levine, H.; Smith, K.A.; Ritz, J. Functional consequences of interleukin 2 receptor expression on resting human lymphocytes. Identification of a novel natural killer cell subset with high affinity receptors. J. Exp. Med. 1990, 171, 1509–1526. [Google Scholar] [CrossRef]
- Trinchieri, G.; Matsumoto-Kobayashi, M.; Clark, S.C.; Seehra, J.; London, L.; Perussia, B. Response of resting human peripheral blood natural killer cells to interleukin 2. J. Exp. Med. 1984, 160, 1147–1169. [Google Scholar] [CrossRef] [PubMed]
- Frey, M.; Packianathan, N.B.; Fehniger, T.A.; Ross, M.E.; Wang, W.-C.; Stewart, C.C.; Caligiuri, M.A.; Evans, S.S. Differential expression and function of L-selectin on CD56bright and CD56dim natural killer cell subsets. J. Immunol. 1998, 161, 400–408. [Google Scholar]
- André, P.; Spertini, O.; Guia, S.; Rihet, P.; Dignat-George, F.; Brailly, H.; Sampol, J.; Anderson, P.J.; Vivier, E. Modification of P-selectin glycoprotein ligand-1 with a natural killer cell-restricted sulfated lactosamine creates an alternate ligand for L-selectin. Proc. Natl. Acad. Sci. USA 2000, 97, 3400–3405. [Google Scholar] [CrossRef] [PubMed]
- Bryceson, Y.T.; March, M.E.; Barber, D.F.; Ljunggren, H.-G.; Long, E.O. Cytolytic granule polarization and degranulation controlled by different receptors in resting NK cells. J. Exp. Med. 2005, 202, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef]
- Voss, S.D.; Daley, J.; Ritz, J.; Robertson, M.J. Participation of the CD94 receptor complex in costimulation of human natural killer cells. J. Immunol. 1998, 160, 1618–1626. [Google Scholar]
- Campbell, J.J.; Qin, S.; Unutmaz, D.; Soler, D.; Murphy, K.E.; Hodge, M.R.; Wu, L.; Butcher, E.C. Unique subpopulations of CD56+ NK and NK-T peripheral blood lymphocytes identified by chemokine receptor expression repertoire. J. Immunol. 2001, 166, 6477–6482. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Fuchs, A.; Colonna, M.; Caligiuri, M.A. NK cell and DC interactions. Trends Immunol. 2004, 25, 47–52. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Huntington, N.D.; Nutt, S.L.; Smyth, M.J. Functional subsets of mouse natural killer cells. Immunol. Rev. 2006, 214, 47–55. [Google Scholar] [CrossRef]
- Lanier, L.L.; Corliss, B.; Wu, J.; Phillips, J.H. Association of DAP12 with Activating CD94/NKG2C NK Cell Receptors. Immunity 1998, 8, 693–701. [Google Scholar] [CrossRef]
- Garni-Wagner, B.A.; Purohit, A.; Mathew, P.A.; Bennett, M.; Kumar, V. A novel function-associated molecule related to non-MHC-restricted cytotoxicity mediated by activated natural killer cells and T cells. J. Immunol. 1993, 151, 60–70. [Google Scholar] [PubMed]
- Pende, D.; Parolini, S.; Pessino, A.; Sivori, S.; Augugliaro, R.; Morelli, L.; Marcenaro, E.; Accame, L.; Malaspina, A.; Biassoni, R.; et al. Identification and molecular characterization of NKp30, a novel triggering receptor involved in natural cytotoxicity mediated by human natural killer cells. J. Exp. Med. 1999, 190, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Bottino, C.; Vitale, M.; Pessino, A.; Augugliaro, R.; Malaspina, A.; Parolini, S.; Moretta, L.; Moretta, A.; Biassoni, R. NKp44, a triggering receptor involved in tumor cell lysis by activated human natural killer cells, is a novel member of the immunoglobulin superfamily. J. Exp. Med. 1999, 189, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Pessino, A.; Sivori, S.; Bottino, C.; Malaspina, A.; Morelli, L.; Moretta, L.; Biassoni, R.; Moretta, A. Molecular cloning of NKp46: A novel member of the immunoglobulin superfamily involved in triggering of natural cytotoxicity. J. Exp. Med. 1998, 188, 953–960. [Google Scholar] [CrossRef]
- Vitale, M.; Bottino, C.; Sivori, S.; Sanseverino, L.; Castriconi, R.; Marcenaro, E.; Augugliaro, R.; Moretta, L.; Moretta, A. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non–major histocompatibility complex–restricted tumor cell lysis. J. Exp. Med. 1998, 187, 2065–2072. [Google Scholar] [CrossRef]
- Hollyoake, M.; Campbell, R.D.; Aguado, B. NKp30 (NCR3) is a pseudogene in 12 inbred and wild mouse strains, but an expressed gene in Mus caroli. Mol. Biol. Evol. 2005, 22, 1661–1672. [Google Scholar] [CrossRef]
- Sivori, S.; Pende, D.; Bottino, C.; Marcenaro, E.; Pessino, A.; Biassoni, R.; Moretta, L.; Moretta, A. NKp46 is the major triggering receptor involved in the natural cytotoxicity of fresh or cultured human NK cells. Correlation between surface density of NKp46 and natural cytotoxicity against autologous, allogeneic or xenogeneic target cells. Eur. J. Immunol. 1999, 29, 1656–1666. [Google Scholar] [CrossRef]
- De Maria, A.; Fogli, M.; Costa, P.; Murdaca, G.; Puppo, F.; Mavilio, D.; Moretta, A.; Moretta, L. The impaired NK cell cytolytic function in viremic HIV-1 infection is associated with a reduced surface expression of natural cytotoxicity receptors (NKp46, NKp30 and NKp44). Eur. J. Immunol. 2003, 33, 2410–2418. [Google Scholar] [CrossRef]
- Costello, R.T.; Sivori, S.; Marcenaro, E.; Lafage-Pochitaloff, M.; Mozziconacci, M.-J.; Reviron, D.; Gastaut, J.-A.; Pende, D.; Olive, D.; Moretta, A. Defective expression and function of natural killer cell–triggering receptors in patients with acute myeloid leukemia. Blood 2002, 99, 3661–3667. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Gupta, R.; Jung, E.; Brunak, S. NetNGlyc 1.0 Server—Prediction of N-glycosylation sites in human proteins. Available online: http://www.cbs.dtu.dk/services/NetNGlyc/ (accessed on 14 August 2018).
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.B.G.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Arnon, T.I.; Markel, G.; Mandelboim, O. Tumor and viral recognition by natural killer cells receptors. Semin. Cancer Biol. 2006, 16, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Moretta, A.; Biassoni, R.; Bottino, C.; Mingari, M.C.; Moretta, L. Natural cytotoxicity receptors that trigger human NK-cell-mediated cytolysis. Immunol. Today 2000, 21, 228–234. [Google Scholar] [CrossRef]
- Biassoni, R.; Pessino, A.; Bottino, C.; Pende, D.; Moretta, L.; Moretta, A. The murine homologue of the human NKp46, a triggering receptor involved in the induction of natural cytotoxicity. Eur. J. Immunol. 1999, 29, 1014–1020. [Google Scholar] [CrossRef]
- Watzl, C.; Long, E.O. Signal transduction during activation and inhibition of natural killer cells. Curr. Protoc. Immunol. 2010, 90, 11.19B.11–11.19B.17. [Google Scholar] [CrossRef]
- Campbell, K.S.; Yusa, S.-I.; Kikuchi-Maki, A.; Catina, T.L. NKp44 triggers NK cell activation through DAP12 association that is not influenced by a putative cytoplasmic inhibitory sequence. J. Immunol. 2004, 172, 899–906. [Google Scholar] [CrossRef]
- Byrd, A.; Hoffmann, S.C.; Jarahian, M.; Momburg, F.; Watzl, C. Expression analysis of the ligands for the natural killer cell receptors NKp30 and NKp44. PLoS ONE 2007, 2, e1339. [Google Scholar] [CrossRef]
- Augugliaro, R.; Parolini, S.; Castriconi, R.; Marcenaro, E.; Cantoni, C.; Nanni, M.; Moretta, L.; Moretta, A.; Bottino, C. Selective cross-talk among natural cytotoxicity receptors in human natural killer cells. Eur. J. Immunol. 2003, 33, 1235–1241. [Google Scholar] [CrossRef]
- Meresse, B.; Curran, S.A.; Ciszewski, C.; Orbelyan, G.; Setty, M.; Bhagat, G.; Lee, L.; Tretiakova, M.; Semrad, C.; Kistner, E.; et al. Reprogramming of CTLs into natural killer–like cells in celiac disease. J. Exp. Med. 2006, 203, 1343–1355. [Google Scholar] [CrossRef]
- Stewart, C.A.; Walzer, T.; Robbins, S.H.; Malissen, B.; Vivier, E.; Prinz, I. Germ-line and rearranged Tcrd transcription distinguish bona fide NK cells and NK-like γδ T cells. Eur. J. Immunol. 2007, 37, 1442–1452. [Google Scholar] [CrossRef]
- Bensussan, A.; Remtoula, N.; Sivori, S.; Bagot, M.; Moretta, A.; Marie-Cardine, A. Expression and function of the natural cytotoxicity receptor NKp46 on circulating malignant CD4+ T lymphocytes of Sézary Syndrome patients. J. Invest. Dermatol. 2011, 131, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Falco, M.; Cantoni, C.; Bottino, C.; Moretta, A.; Biassoni, R. Identification of the rat homologue of the human NKp46 triggering receptor. Immunol. Lett. 1999, 68, 411–414. [Google Scholar] [CrossRef]
- De Maria, A.; Ugolotti, E.; Rutjens, E.; Mazza, S.; Radic, L.; Faravelli, A.; Koopman, G.; Di Marco, E.; Costa, P.; Ensoli, B.; et al. NKp44 expression, phylogenesis and function in non-human primate NK cells. Int. Immunol. 2009, 21, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Mair, K.H.; Essler, S.E.; Patzl, M.; Storset, A.K.; Saalmüller, A.; Gerner, W. NKp46 expression discriminates porcine NK cells with different functional properties. Eur. J. Immunol. 2012, 42, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Storset, A.K.; Kulberg, S.; Berg, I.; Boysen, P.; Hope, J.C.; Dissen, E. NKp46 defines a subset of bovine leukocytes with natural killer cell characteristics. Eur. J. Immunol. 2004, 34, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Straub, C.; Neulen, M.-L.; Sperling, B.; Windau, K.; Zechmann, M.; Jansen, C.A.; Viertlboeck, B.C.; Göbel, T.W. Chicken NK cell receptors. Dev. Comp. Immunol. 2013, 41, 324–333. [Google Scholar] [CrossRef]
- Seidel, E.; Glasner, A.; Mandelboim, O. Virus-mediated inhibition of natural cytotoxicity receptor recognition. Cell. Mol. Life Sci. 2012, 69, 3911–3920. [Google Scholar] [CrossRef]
- Gur, C.; Enk, J.; Kassem, S.A.; Suissa, Y.; Magenheim, J.; Stolovich-Rain, M.; Nir, T.; Achdout, H.; Glaser, B.; Shapiro, J.; et al. Recognition and killing of human and murine pancreatic β cells by the NK receptor NKp46. J. Immunol. 2011, 187, 3096–3103. [Google Scholar] [CrossRef]
- Narni-Mancinelli, E.; Gauthier, L.; Baratin, M.; Guia, S.; Fenis, A.; Deghmane, A.-E.; Rossi, B.; Fourquet, P.; Escalière, B.; Kerdiles, Y.M.; et al. Complement factor P is a ligand for the natural killer cell–activating receptor NKp46. Sci. Immunol. 2017, 2, eaam9628. [Google Scholar] [CrossRef]
- Fuchs, A.; Cella, M.; Kondo, T.; Colonna, M. Paradoxic inhibition of human natural interferon-producing cells by the activating receptor NKp44. Blood 2005, 106, 2076–2082. [Google Scholar] [CrossRef]
- Von Lilienfeld-Toal, M.; Nattermann, J.; Feldmann, G.; Sievers, E.; Frank, S.; Strehl, J.; Schmidt-Wolf, I.G.H. Activated γδ T cells express the natural cytotoxicity receptor natural killer p44 and show cytotoxic activity against myeloma cells. Clin. Exp. Immunol. 2006, 144, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Baychelier, F.; Sennepin, A.; Ermonval, M.; Dorgham, K.; Debré, P.; Vieillard, V. Identification of a cellular ligand for the natural cytotoxicity receptor NKp44. Blood 2013, 122, 2935–2942. [Google Scholar] [CrossRef] [PubMed]
- Rosental, B.; Brusilovsky, M.; Hadad, U.; Oz, D.; Appel, M.Y.; Afergan, F.; Yossef, R.; Rosenberg, L.A.; Aharoni, A.; Cerwenka, A.; et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J. Immunol. 2011, 187, 5693–5702. [Google Scholar] [CrossRef] [PubMed]
- De Maria, A.; Biassoni, R.; Fogli, M.; Rizzi, M.; Cantoni, C.; Costa, P.; Conte, R.; Mavilio, D.; Ensoli, B.; Cafaro, A.; et al. Identification, molecular cloning and functional characterization of NKp46 and NKp30 natural cytotoxicity receptors in Macaca fascicularis NK cells. Eur. J. Immunol. 2001, 31, 3546–3556. [Google Scholar] [CrossRef]
- Tang, Q.; Grzywacz, B.; Wang, H.; Kataria, N.; Cao, Q.; Wagner, J.E.; Blazar, B.R.; Miller, J.S.; Verneris, M.R. Umbilical cord blood T cells express multiple natural cytotoxicity receptors after IL-15 stimulation, but only NKp30 Is functional. J. Immunol. 2008, 181, 4507–4515. [Google Scholar] [CrossRef]
- Correia, D.V.; Fogli, M.; Hudspeth, K.; da Silva, M.G.; Mavilio, D.; Silva-Santos, B. Differentiation of human peripheral blood Vδ1+ T cells expressing the natural cytotoxicity receptor NKp30 for recognition of lymphoid leukemia cells. Blood 2011, 118, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Hudspeth, K.; Fogli, M.; Correia, D.V.; Mikulak, J.; Roberto, A.; Della Bella, S.; Silva-Santos, B.; Mavilio, D. Engagement of NKp30 on Vδ1 T cells induces the production of CCL3, CCL4, and CCL5 and suppresses HIV-1 replication. Blood 2012, 119, 4013–4016. [Google Scholar] [CrossRef]
- Ferlazzo, G.; Tsang, M.L.; Moretta, L.; Melioli, G.; Steinman, R.M.; Münz, C. Human dendritic cells activate resting natural killer (NK) cells and are recognized via the NKp30 receptor by activated NK cells. J. Exp. Med. 2002, 195, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Chiesa, M.D.; Carlomagno, S.; Pende, D.; Aricò, M.; Moretta, L.; Moretta, A. NK-dependent DC maturation is mediated by TNFα and IFNγ released upon engagement of the NKp30 triggering receptor. Blood 2005, 106, 566–571. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Carosio, R.; Pende, D.; Marcenaro, S.; Rivera, P.; Zocchi, M.R.; Moretta, L.; Poggi, A. NK cell-mediated lysis of autologous antigen-presenting cells is triggered by the engagement of the phosphatidylinositol 3-kinase upon ligation of the natural cytotoxicity receptors NKp30 and NKp46. Eur. J. Immunol. 2001, 31, 1656–1665. [Google Scholar] [CrossRef]
- Brandt, C.S.; Baratin, M.; Yi, E.C.; Kennedy, J.; Gao, Z.; Fox, B.; Haldeman, B.; Ostrander, C.D.; Kaifu, T.; Chabannon, C.; et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J. Exp. Med. 2009, 206, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- von Strandmann, E.P.; Simhadri, V.R.; von Tresckow, B.; Sasse, S.; Reiners, K.S.; Hansen, H.P.; Rothe, A.; Böll, B.; Simhadri, V.L.; Borchmann, P.; et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity 2007, 27, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Ponassi, M.; Biassoni, R.; Conte, R.; Spallarossa, A.; Moretta, A.; Moretta, L.; Bolognesi, M.; Bordo, D. The three-dimensional structure of the human NK cell receptor NKp44, a triggering partner in natural cytotoxicity. Structure 2003, 11, 725–734. [Google Scholar] [CrossRef]
- World Health Organization. Influenza (Seasonal). Available online: http://www.who.int/mediacentre/factsheets/fs211/en/ (accessed on 3 July 2018).
- Carlin, L.E.; Hemann, E.A.; Zacharias, Z.R.; Heusel, J.W.; Legge, K.L. Natural killer cell recruitment to the lung during influenza a virus infection is dependent on CXCR3, CCR5, and virus exposure dose. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Pommerenke, C.; Wilk, E.; Srivastava, B.; Schulze, A.; Novoselova, N.; Geffers, R.; Schughart, K. Global transcriptome analysis in influenza-infected mouse lungs reveals the kinetics of innate and adaptive host immune responses. PLoS ONE 2012, 7, e41169. [Google Scholar] [CrossRef]
- Taub, D.D.; Sayers, T.J.; Carter, C.R.; Ortaldo, J.R. Alpha and beta chemokines induce NK cell migration and enhance NK-mediated cytolysis. J. Immunol. 1995, 155, 3877–3888. [Google Scholar]
- Cooper, G.E.; Ostridge, K.; Khakoo, S.I.; Wilkinson, T.M.A.; Staples, K.J. Human CD49a(+) Lung Natural Killer Cell Cytotoxicity in Response to Influenza A Virus. Front. Immunol. 2018, 9, 1671. [Google Scholar] [CrossRef]
- Scharenberg, M.; Vangeti, S.; Kekäläinen, E.; Bergman, P.; Al-Ameri, M.; Johansson, N.; Sondén, K.; Falck-Jones, S.; Färnert, A.; Ljunggren, H.G.; et al. Influenza A Virus Infection Induces Hyperresponsiveness in Human Lung Tissue-Resident and Peripheral Blood NK Cells. Front. Immunol. 2019, 10, 1116. [Google Scholar] [CrossRef]
- Prager, I.; Watzl, C. Mechanisms of natural killer cell-mediated cellular cytotoxicity. J. Leukoc. Biol. 2019, 105, 1319–1329. [Google Scholar] [CrossRef]
- Stein-Streilein, J.; Guffee, J. In vivo treatment of mice and hamsters with antibodies to asialo GM1 increases morbidity and mortality to pulmonary influenza infection. J. Immunol. 1986, 136, 1435–1441. [Google Scholar]
- Abdul-Careem, M.F.; Mian, M.F.; Yue, G.; Gillgrass, A.; Chenoweth, M.J.; Barra, N.G.; Chew, M.V.; Chan, T.; Al-Garawi, A.A.; Jordana, M.; et al. Critical role of natural killer cells in lung immunopathology during influenza infection in mice. J. Infect. Dis. 2012, 206, 167–177. [Google Scholar] [CrossRef]
- Zhou, G.; Juang, S.W.W.; Kane, K.P. NK cells exacerbate the pathology of influenza virus infection in mice. Eur. J. Immunol. 2013, 43, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.Q.; Ho, A.W.S.; Tang, Y.; Wong, K.H.S.; Chua, B.Y.L.; Gasser, S.; Kemeny, D.M. NK cells regulate CD8+ T cell priming and dendritic cell migration during influenza a infection by IFN-γ and perforin-dependent mechanisms. J. Immunol. 2012, 189, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, S.G.; Huprikar, J. The influence of defective-interfering particles of the PR-8 strain of influenza A virus on the pathogenesis of pulmonary infection in mice. J. Infect. Dis. 1979, 140, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Wang, J.; Li, A.; Zhao, W.; Wang, D.; Zhang, W.; Yan, J.; Gao, G.F.; Liu, W.; Fang, M. Swift and strong NK cell responses protect 129 mice against high-dose influenza virus infection. J. Immunol. 2016, 196, 1842–1854. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Tu, W.; Qin, G.; Law, H.K.W.; Sia, S.F.; Chan, P.-L.; Liu, Y.; Lam, K.-T.; Zheng, J.; Peiris, M.; et al. Influenza virus virectly infects human natural killer cells and induces cell apoptosis. J. Virol. 2009, 83, 9215–9222. [Google Scholar] [CrossRef]
- Guo, H.; Kumar, P.; Moran, T.M.; Garcia-Sastre, A.; Zhou, Y.; Malarkannan, S. The functional impairment of natural killer cells during influenza virus infection. Immunol. Cell Biol. 2009, 87, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Cross, K.J.; Burleigh, L.M.; Steinhauer, D.A. Mechanisms of cell entry by influenza virus. Expert Rev. Mol. Med. 2001, 3, 1–18. [Google Scholar] [CrossRef]
- Skehel, J.J.; Wiley, D.C. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef]
- Seto, J.T.; Chang, F.S. Functional significance of sialidase during influenza virus multiplication: An electron microscope study. J. Virol. 1969, 4, 58–66. [Google Scholar] [CrossRef]
- Seto, J.T.; Rott, R. Functional significance of sialidase during influenza virus multiplication. Virology 1966, 30, 731–737. [Google Scholar] [CrossRef]
- Wiley, D.C.; Skehel, J.J.; Waterfield, M. Evidence from studies with a cross-linking reagent that the haemagglutinin of influenza virus is a trimer. Virology 1977, 79, 446–448. [Google Scholar] [CrossRef]
- Wilson, I.A.; Skehel, J.J.; Wiley, D.C. Structure of the haemagglutinin membrane glycoprotein of influenza at 3 Å resolution. Nature 1981, 289, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lee, K.H.; Steinhauer, D.A.; Stevens, D.J.; Skehel, J.J.; Wiley, D.C. Structure of the hemagglutinin precursor cleavage site, a determinant of influenza pathogenicity and the origin of the labile conformation. Cell 1998, 95, 409–417. [Google Scholar] [CrossRef]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Bordoli, L.; Kiefer, F.; Arnold, K.; Benkert, P.; Battey, J.; Schwede, T. Protein structure homology modeling using SWISS-MODEL workspace. Nat. Protoc. 2008, 4, 1–13. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses. Family: Orthomyxoviridae. Available online: https://talk.ictvonline.org/ictv-reports/ictv_9th_report/negative-sense-rna-viruses-2011/w/negrna_viruses/209/orthomyxoviridae (accessed on 10 November 2020).
- Bullough, P.A.; Hughson, F.M.; Skehel, J.J.; Wiley, D.C. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature 1994, 371, 37–43. [Google Scholar] [CrossRef]
- Skehel, J.J.; Bayley, P.M.; Brown, E.B.; Martin, S.R.; Waterfield, M.D.; White, J.M.; Wilson, I.A.; Wiley, D.C. Changes in the conformation of influenza virus hemagglutinin at the pH optimum of virus-mediated membrane fusion. Proc. Natl. Acad. Sci. USA 1982, 79, 968–972. [Google Scholar] [CrossRef]
- Han, X.; Bushweller, J.H.; Cafiso, D.S.; Tamm, L.K. Membrane structure and fusion-triggering conformational change of the fusion domain from influenza hemagglutinin. Nat. Struct. Biol. 2001, 8, 715–720. [Google Scholar] [CrossRef]
- Han, X.; Tamm, L.K. pH-dependent self-association of influenza hemagglutinin fusion peptides in lipid bilayers. J. Mol. Biol. 2000, 304, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Gamblin, S.J.; Haire, L.F.; Russell, R.J.; Stevens, D.J.; Xiao, B.; Ha, Y.; Vasisht, N.; Steinhauer, D.A.; Daniels, R.S.; Elliot, A.; et al. The structure and receptor binding properties of the 1918 influenza hemagglutinin. Science 2004, 303, 1838–1842. [Google Scholar] [CrossRef] [PubMed]
- Gamblin, S.J.; Skehel, J.J. Influenza hemagglutinin and neuraminidase membrane glycoproteins. J. Biol. Chem. 2010, 285, 28403–28409. [Google Scholar] [CrossRef]
- Martín, J.; Wharton, S.A.; Lin, Y.P.; Takemoto, D.K.; Skehel, J.J.; Wiley, D.C.; Steinhauer, D.A. Studies of the binding properties of influenza hemagglutinin receptor-site mutants. Virology 1998, 241, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.; Kuiken, T. H5N1 virus attachment to lower respiratory tract. Science 2006, 312, 399. [Google Scholar] [CrossRef]
- Van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Human and avian influenza viruses target different cells in the lower respiratory tract of humans and other mammals. Am. J. Pathology 2007, 171, 1215–1223. [Google Scholar] [CrossRef]
- Rogers, G.N.; Paulson, J.C. Receptor determinants of human and animal influenza virus isolates: Differences in receptor specificity of the H3 hemagglutinin based on species of origin. Virology 1983, 127, 361–373. [Google Scholar] [CrossRef]
- Reid, A.H.; Janczewski, T.A.; Lourens, R.M.; Elliot, A.J.; Daniels, R.S.; Berry, C.L.; Oxford, J.S.; Taubenberger, J.K. 1918 influenza pandemic and highly conserved viruses with two receptor-binding variants. Emerg. Infect. Dis. 2003, 9, 1249. [Google Scholar] [CrossRef]
- Glaser, L.; Stevens, J.; Zamarin, D.; Wilson, I.A.; García-Sastre, A.; Tumpey, T.M.; Basler, C.F.; Taubenberger, J.K.; Palese, P. A single amino acid substitution in 1918 influenza virus hemagglutinin changes receptor binding specificity. J. Virol. 2005, 79, 11533–11536. [Google Scholar] [CrossRef]
- Xu, R.; McBride, R.; Nycholat, C.M.; Paulson, J.C.; Wilson, I.A. Structural characterization of the hemagglutinin receptor specificity from the 2009 H1N1 influenza pandemic. J. Virol. 2012, 86, 982–990. [Google Scholar] [CrossRef]
- Rogers, G.N.; D’Souza, B.L. Receptor binding properties of human and animal H1 influenza virus isolates. Virology 1989, 173, 317–322. [Google Scholar] [CrossRef]
- Dortmans, J.C.F.M.; Dekkers, J.; Wickramasinghe, I.N.A.; Verheije, M.H.; Rottier, P.J.M.; van Kuppeveld, F.J.M.; de Vries, E.; de Haan, C.A.M. Adaptation of novel H7N9 influenza A virus to human receptors. Sci. Rep. 2013, 3, 3058. [Google Scholar] [CrossRef] [PubMed]
- Mimura, Y.; Ashton, P.R.; Takahashi, N.; Harvey, D.J.; Jefferis, R. Contrasting glycosylation profiles between Fab and Fc of a human IgG protein studied by electrospray ionization mass spectrometry. J. Immunol. Methods 2007, 326, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.M.; Cosgrave, E.F.J.; Struwe, W.B.; Wormald, M.; Davey, G.P.; Jefferis, R.; Rudd, P.M. Glycosylation and Fc receptors. In Current Topics in Microbiology and Immunology; Daeron, M., Nimmerjahn, F., Eds.; Springer: Cham, Switzerland, 2014; Volume 382, pp. 165–199. [Google Scholar]
- Trombetta, C.M.; Ulivieri, C.; Cox, R.J.; Remarque, E.J.; Centi, C.; Perini, D.; Piccini, G.; Rossi, S.; Marchi, S.; Montomoli, E. Impact of erythrocyte species on assays for influenza serology. J. Prev. Med. Hyg. 2018, 59, E1–E7. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, A.; Brusilovsky, M.; Kundu, K.; Ottolenghi, A.; Campbell, K.S.; Porgador, A. Splice variants of human natural cytotoxicity receptors: Novel innate immune checkpoints. Cancer Immunol. Immunother. 2018, 67, 1871–1883. [Google Scholar] [CrossRef]
- Clare, M.A.; Collins, R.; Vaughan, R. Polymorphism in the natural cytotoxicity receptors(NCR) NKp30, NKp44 and NKp46 – a preliminary study. Hum. Immunol. 2005, 66, 18. [Google Scholar] [CrossRef]
- Lee, E.U.; Roth, J.; Paulson, J.C. Alteration of terminal glycosylation sequences on N-linked oligosaccharides of Chinese hamster ovary cells by expression of beta-galactoside alpha 2,6-sialyltransferase. J. Biol. Chem. 1989, 264, 13848–13855. [Google Scholar] [CrossRef]
- Storset, A.K.; Slettedal, I.Ö.; Williams, J.L.; Law, A.; Dissen, E. Natural killer cell receptors in cattle: A bovine killer cell immunoglobulin-like receptor multigene family contains members with divergent signaling motifs. Eur. J. Immunol. 2003, 33, 980–990. [Google Scholar] [CrossRef]
- Meng, B.; Marriott, A.C.; Dimmock, N.J. The receptor preference of influenza viruses. Influenza Resp. Viruses 2010, 4, 147–153. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. Avian Influenza (version adopted May 2015). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Mammals, Birds and Bees); World Organisation for Animal Health: Paris, France, 2015; Volume 1, pp. 1–23. [Google Scholar]
- Swayne, D.E.; Suarez, D.L.; Sims, L.D. Influenza. In Diseases of Poultry, 13th ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; John Wiley & Sons, Inc.: Ames, Iowa, 2013; pp. 181–218. [Google Scholar]
- Luczo, J.M.; Stambas, J.; Durr, P.A.; Michalski, W.P.; Bingham, J. Molecular pathogenesis of H5 highly pathogenic avian influenza: The role of the haemagglutinin cleavage site motif. Rev. Med. Virol. 2015, 25, 406–430. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen (Species) | Isolate | NCRs Investigated | Summary | Reference |
|---|---|---|---|---|
| Influenza A virus | A/Bangkok/1/1979 (H3N2) | - | Pre-treatment of effector cells with purified HA blocked NK cell cytolytic activity. Suggests HA binds and functionally activated NK cells. | Ali et al. 1984 Immunology [52] |
| A/Victoria/1/1975 H3N2) | ||||
| Influenza B virus | B/Lyon/1979 | |||
| Influenza A virus | A//Port Chalmers/1/1973 (H3N2) | - | Whole IAV or rHA functionally activates NK cells. Pre-treatment of IAV with anti-HA antisera or anti-HA F(ab’)2 blocked NK cell cytolytic activity. | Arora et al. 1984 J. Virol. [53] |
| - | - | hNKp46, hCD16 | Biochemical analysis of hNKp46. Suggests that hNKp46 does not harbor N-linked glycosites, though does possess O-linked glycosites. | Sivori et al. 1997 J. Exp. Med. [54] |
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | hNKp46, hCD16 | SeV-infected cells bind NKp46-Ig. Cells transfected with SeV rHN bind NKp46-Ig. IAV HA binds NKp46-Ig, but not CD16-Ig or CD99-Ig. NA-treatment of hNKp46-Ig reduced binding by IAV. Lysis of SeV or IAV infected cells is reduced following anti-HN or anti-HA mAb treatment, respectively. | Mandelboim et al. 2001 Nature [28] |
| Murine respirovirus | Sendai virus | |||
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | hNKp44, hNKp30, hNKp46, hCD16 | Recognition of transformed cell lines by NKp46-Ig, NKp44-Ig, NKp30-Ig, and CD16-Ig, but not CD99-Ig. NKp44-Ig bound SeV or IAV infected, or SeV HN transfected cells. | Arnon et al. 2001 Eur. J. Immunol. [39] |
| Murine respirovirus | Sendai virus | |||
| Mycobacterium tuberculosis | H37Ra strain (avirulent) H37Rv strain (virulent) | hNKp46 | NKp46 blockade with antisera reduced NKp46-mediated NK cell lysis of infected monocytes. | Vankayalapati et al. 2002 J. Immunol. [48] |
| Murine respirovirus | Sendai virus | hNKp46 | hNKp46-Ig, but not hCD99-Ig, binds to SeV infected cells. Blockade of HN and hNKp46 binding with anti-HN mAb reduced binding. | Achdout et al. 2003 J. Immunol. [38] |
| Influenza A virus | A/Beijing/262/1995-like (H1N1) | hNKp46, hNKp44, hNKp30 | SeV, IAV, and IBV bind NKp44-Ig and NKp46-Ig, but poorly to NKp30-Ig, and not at all to CD99-Ig. NA-treatment of NKp46-Ig reduced binding by SeV or IAV. Interaction with NKp46 specifically occurs with the proximal C2-type Ig-like domain. Suggests α-2,6 sialic acids on Thr225 critical for interaction. | Arnon et al. 2004 Blood [29] |
| A/Moscow/10/1999-like (H3N2) | ||||
| A/Sydney/5/1997-like (H3N2) | ||||
| A/X-31 (A/Aichi/2/1968*A/Puerto Rico/8/1934) (H3N2) | ||||
| A/X-127 (A/Beijing/262/1995* A/Puerto Rico/8/1934) (H1N1) | ||||
| A/New Caledonia/20/1999 (H1N1) | ||||
| Influenza B virus | B/Yamanashi/166/1998 (Yamagata lineage) | |||
| Murine respirovirus | Sendai virus | |||
| - | - | hNKp46, hNKp30 | Identified 6-O-sulfo-N-acetylglucosamine and hNK46 and hNKp30 interactions. Pretreatment of NKp30-Ig or NKp46-Ig with 6-O-sulfo-LacNAc reduced NCR binding to tumor cells. Pre-treatment with 3-O-sulfo-LacNAc or 4,6-di-O-sulfo-LacNAc did not affect binding. Additionally, heparin or heparan sulfate pre-treatment inhibited binding of NCRs to tumor cells. | Bloushtain et al. 2004 J. Immunol. [55] |
| Human betaherpesvirus 5 | Human cytomegalovirus (AD169 strain) | hNKp46, hNKp44, hNKp30, hCD16 | HCMV-infected cells bind NKp30-Ig, no binding to NKp46-Ig, NKp44-Ig, CD16-Ig or CD99-Ig was detected. Pulldown experiments identified HCMV pp65 protein bound to NKp30-Ig. Recombinant pp65 bound NKp30-Ig only (KD 10 nM). Preincubation of NK cells with rpp65 blocked anti-NKp30 mAb binding. Counterintuitively, pp65 engagement of NKp30-Ig resulted in reduced NK cell cytolytic activity, resulting from CD3ζ chain dissociation from NKp30 complex. | Arnon et al. 2005 Nat. Immunol. [42] |
| Human immunodeficiency virus 1 | Human immunodeficiency virus 1 (Sf2 strain) | hNKp44 | HIV-1 infected cells induce NKp44L expression, and are highly susceptible to NK cell-mediated lysis. Ex vivo, NKp46L expressed on CD4+, but not CD8+, HIV positive individuals. NKp46L induced by gp41 (and its precursor, gp160). Anti-gp41 treatment of infected cells reduced NKp44L expression and blocked NK cytolytic activity. | Vieillard et al. 2005 PNAS [56] |
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | msNKp46 | IAV HA binds msNKp46-Ig. Generated NCR1gfp/gfp knockout mice. Lack of NCR1 resulted in 100% mortality following IAV infection. | Gazit et al. 2006 Nat. Immunol. [21] |
| Vaccinia virus | Modified vaccinia virus Ankara | hNKp46, hNKp44, hNKp30 | Vaccinia virus-infected cells recognize NKp30-Ig, NKp44-Ig, and NKp46-Ig. NKp30-Ig ligand was the most potently induced on virally-infected cells. Anti-NCR mAb blockade abrogated virally-infected cell:NCR interactions. Virally-induced ligand not directly identified. | Chisholm et al. 2006 J. Virol. [45] |
| Vaccinia virus Western Reserve | ||||
| Vaccinia virus Copenhagen | ||||
| Vaccinia virus Wyeth | ||||
| Vaccinia virus Lister | ||||
| Vaccinia virus IHD-J | ||||
| Vaccinia virus IHD-W | ||||
| Vaccinia virus Tian-Tan | ||||
| Vaccinia virus Tashkent | ||||
| Vaccinia virus USSR | ||||
| Vaccinia virus Patwadangar | ||||
| Vaccinia virus King Institute | ||||
| Vaccinia virus Dairen | ||||
| Buffalopox virus | ||||
| Rabbitpox virus (strain unknown) | ||||
| Vaccinia virus Evans | ||||
| Cowpox virus | Cowpox virus Brighton Red | |||
| Elephantpox virus | ||||
| Camelpox virus | Camelpox virus (strain unknown) | |||
| Mycobacterium tuberculosis | H37Ra strain (avirulent) | hNKp46 | NKp46-Ig binds to TB-infected monocytes. Immunoprecipitation and mass spectrometry analysis of NKp46-Ig membrane bound ligands from TB-infected monocytes identified vimentin (type III intermediate filament) as a ligand of NKp46. Vimentin was shown to be surface exposed on TB-infected monocytes. Vimentin was also upregulated on Listeria-infected monocytes. Vimentin antisera reduced NK cell-mediated lysis. | Garg et al. 2006 J. Immunol. [49] |
| Listeria monocytogenese | Lm68 strain, Serovar 1/2b | |||
| Human alphaherpesvirus 1 | Herpes simplex virus 1 (strain 17) Herpes simplex virus 1 (strain F) | hNKp46, hNKp44, hNKp30 | HSV1 virally-infected cells and HSV1 infected cell protein 0 (ICP0)-transfected cells are more susceptible to NK cell lysis. Cells infected with ∆ICP0 HSV mutant were less susceptible to lysis. Antibody blockade of the NCRs reduced NK cell lysis of HSV1-infected cells. Blockade of all three NCRs abrogate NK cell lysis of target cells. ICP0 is not surface expressed, thus ligand is likely to be cellular. | Chisholm et al. 2007 J. Infect. Dis. [43] |
| Plasmodium falciparum | FCR-3, D10, FP8, and 3D7 strains | hNKp46, hNKp30, hCD99 | P. falciparum erythrocyte membrane protein-1 duffy binding-like 1α domain peptides bound NKp30-Ig, and minimally with NKp46-Ig. NKp46 and NKp30 bound to P. falciparum infected erythrocytes. NCRs bound to proximal Ig-like domain. Treatment with trypsin abrogated erythrocyte:NCR interaction. Blockade with anti-NKp46 or NKp30 reduced NK cell cytolytic activity. | Mavoungou et al. 2007 J. Infect. Dis. [51] |
| Influenza A virus | A/England/878/1969 (H3N2) | - | CD56+ CD3- NK cells express both α-2,3 and α-2,6 sialic acids. Historical H3N2 virally-infected cells are lysed more efficiently by NK cells than those infected with contemporary H3N2 viruses. Treatment of NK cells with NA reduced NK-mediated lysis of IAV-infected cells. | Owen et al. 2007 J. Virol. [57] |
| A/England/401/1985 (H3N2) | ||||
| A/England/327/1990 (H3N2) | ||||
| A/England/289/1993 (H3N2) | ||||
| A/England/41/1996 (H3N2) | ||||
| A/England/356/1996 (H3N2) | ||||
| A/England/26/1999 (H3N2) | ||||
| A/England/919/1999 (H3N2) | ||||
| A/England/24/2000 (H3N2) | ||||
| Mycobacterium avium | NBL112/87 strain | hNKp46, hNKp44, hNKp30 | M. bovis induced expression of NKp44 on CD56bright NK cells, but not NKp30 or NKp46. All mycobacterium tested bound to NKp46-Ig. Additionally, N. farcinica (Gram-positive) and P. aeruginosa (Gram-negative) interacted with NKp44-Ig, minimally with NKp46-Ig, and not at all with NKp30-Ig. Electron microscopy revealed NKp44-Ig bound to surface of M. bovis and E. faecium, but not NKp46-Ig or NKp30-Ig. Interestingly, anti-NCR mAbs did not reduce M. bovis-induced NK cell activation, however, NKp44-Ig mAb reduced binding of NKp44-Ig to M. bovis. | Esin et al. 2008 J. Virol. [50] |
| Mycobacterium smegmatis | mc2 155 strain | |||
| Mycobacterium tuberculosis | H37Rv strain (virulent) | |||
| Mycobacterium bovis | ||||
| Salmonella enterica | Bacillus Calmette-Guérin (Pasteur) strain | |||
| Escherichia coli | ||||
| Streptococcus pyogenes | Serovar Enteritidis | |||
| Enterococcus faecium | ||||
| Pseudomonas aeruginosa | ||||
| Actinomyces meyeri | ||||
| Cellulomonas denverensis | DSM 15764 | |||
| Nocardia farcinica | DSM 43665 | |||
| Influenza A virus | A/VNH5N1-PR8/CDC-rg (A/Vietnam/1203/2004*A/Puerto Rico/8/1934) (H5N1–LPAIV) A/Cambodia/408008/2005 (H5N1–HPAIV) A/Puerto Rico/8/1934 (H1N1) | hNKp44, hNKp30 | Infection of cells with IAV enhances NK cell mediated lysis. Cells transfected with IAV HA (H5N1–HPAIV) bind NKp44-Ig, poorly to NKp30-Ig. rHA binds hNKp-44-Ig (ELISA). IAV pseudoparticles bind to hNKp44 transduced cells. | Ho et al. 2008 J. Virol. [30] |
| West Nile virus Dengue virus | West Nile virus ISR98-Goo1 Dengue virus 1 FGA/89 Dengue virus 4 Burma 1976 | hNKp44, hNKp46, hNKp30 | Recombinant DENV and WNV E glycoproteins bind hNKp44-Ig. WNV rE glycoprotein bound membrane-associated hNKp44-Ig. Binding was also demonstrated for WNV VLPs (E and prM glycoproteins) and virally-infected cells. VLP and hNKP44-Ig interaction was augmented by low pH treatment. Interactions mapped to domain III of WNV E glycoprotein. Anti-NKp44 serum reduced WNV E binding to hNKp44-Ig and reduce NK cell cytolytic activity. | Hershkovitz et al. 2009 J. Immunol [47] |
| Avian orthoavulavirus 1 | Newcastle disease virus (Ulster 2C strain) Newcastle disease virus (MTH-68/H strain) | hNK46, hNKp44, hNKp30 | NDV-infected cells bind hNKp46-Ig and hNKp44-Ig, but not hNKp30-Ig. Treatment with anti-HN mAb reduced HN binding to hNKp46-Ig and hNKp44-Ig. HN or F transfected cells revealed interaction directly mediated by HN. Desialylation of hNKp46-Ig and hNKp44-Ig abrogated interaction with HN. NK cells lysis blocked when NDV cells treated with NA-inhibitor or anti-HN mAb. | Jarahian et al. 2009 J. Virol. [40] |
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | hNKp46 | Suggests Thr225 does not harbor a unique glycoform (compared to Thr125). Characterized α2,3- and α2,6- O-linked glycans present on HEK293T, CHO and COS-7 expressed hNKp46. | Mendelson et al. 2010 J. Virol. [31] |
| A/Brisbane/59/2007 (H1N1) | ||||
| A/New Caledonia/20/1999 (H1N1) | ||||
| Influenza A virus | A/VNH5N1-PR8/CDC-rg (A/Vietnam/1203/2004*A/Puerto Rico/8/1934) (H5N1–LPAIV) A/Puerto Rico/8/1934 (H1N1) A/Texas/1/1977 (H3N2) A/Swine/Israel/2009 (pdmH1N1) | hNKp46, hNKp30 | Avian and human influenza viruses interact with hNKp46, although H5N1 HA:hNkp46 interaction did not induce NK cell-mediated lysis of H5 infected cells, whereas H1N1 did. Authors conclude no increase in lysis of H5 cells as H5 avian viruses have not evolved/adapted to humans. | Achdout et al. 2010 J. Virol. [58] |
| Influenza A virus | A/Brevig Mission/1/1918 (pdmH1N1) A/California/07/2009 (pdmH1N1) A/Anhui/1/2005 (H5N1–HPAIV) A/Brisbane/10/2007 (H3N2) | hNKp46 | In contrast to Achdout et al. 2010, Du et al. H5N1 and 1918 pdmH1N1 HA (pseudoviruses) induced greater NK cell activation and lysis, than 2009 pdmH1N1 IAV. hNKp46 was found to be downregulated upon stimulation. Treatment with anti-NKp46 mAb lead to reduced CD69 expression. | Du et al. 2010 J. Virol. [32] |
| Influenza A virus | A/Hong Kong/54/1998 (H1N1) | - | Pre-treatment of NK cells with whole IAV virions or rHA inhibits NK cell cytolytic activity by way of downregulation of the CD3 ζ chain and cytolytic granule exocytosis (NKp46 and NKp30 surface expression levels remained unchanged). NA-treatment of human NK cells reduces rHA internalization. | Mao et al. 2010 J. Virol. [59] |
| A/New Caledonia/20/1999 (H1N1) | ||||
| - | - | hNKp46 | Glycan-binding analysis of E. coli expressed NKp46-His and sulfate- or neuraminic acid containing multimeric glycans. Recombinant hNKp46 bound heparin-BSA and heparan-sulfate-BSA in the low µM range; 2-O-linked, 6-O-linked, and N-linked sulfates important for interaction. Binding to Sialyl Lewis X-expressing transferrin also demonstrated. | Ito et al. 2011 Biochem. Biophys. Res. Commun. [60] |
| Vaccinia virus Ectromelia virus | Vaccinia virus (Western Reserve strain) Ectromelia virus (MP-Nü strain) | hNKp46, hNKp30, hNKp44 | VV and ECTV infected cells upregulated ligands for hNKp46-Ig and hNKp30-Ig, but not hNKp44-Ig. ∆HA virus did not induce NCR ligands. Anti-HA mAb blocked HA hNKp46-Ig and hNKp30-Ig interactions. Desialyation/deglycosylation of hNKp46-Ig reduced binding with HA, though increased binding with hNKp30-Ig. Demonstrated that VV HA reduces B7-H6 binding to hNKp30. VV-infected cells less susceptible to NK cell-mediated lysis. | Jarahian et al. 2011 PLoS Path. [46] |
| Fusobacterium nucleatum Influenza A virus | PK1594 strain | hNKp46, msNKp46, hNKp44, hNKp30, hCD16 | F. nucleatum bacterium bind NKp46-Ig and NCR1-Ig, minimally with NKp44-Ig, and not at all with NKp30-Ig, CD16-Ig. Interaction was not sialic acid-dependent; and was heat, proteinase K, and pronase sensitive. | Chaushu et al. 2012 PLoS Path. [33] |
| A/Puerto Rico/8/1934 (H1N1) | ||||
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | msNKp46 | Binding of IAV HA to NCR1 is mediated by N-linked glycosylation. Deglycosylation of msNKp46 with PNGase F abolished binding. Deglycosylation of msNKp46 with O-linked glycanase cocktail had little effect on binding. Surprisingly, mutation of Asn139, Asn216 or Asn238 N-linked glycosites did not affect binding. | Glasner et al. 2012 PLoS ONE [22] |
| - | - | hNKp44, hNKp30 | Glycan-binding analysis of E. coli expressed NKp44-His and NKp30-His to sulfate- or neuraminic acid containing multimeric glycans. Recombinant hNKp46 and hNKp30 bound heparin-BSA in the low to mid nM range. NKp44, but not NKp30, bound Sialyl Lewis X-expressing transferrin. NA-treatment of transferrin abrogated binding. | Ito et al. 2012 Biol. Pharm. Bull. [61] |
| Human gammaherpesvirus 8 | Kaposi’s sarcoma-associated herpesvirus | hNKp44 | NKp44-Ig recognizes KSHV-infected cells. KSHV ORF54 downregulates NKp44L surface expression on KSHV-infected cells. | Madrid & Ganem 2012 J. Virol. [44] |
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | hNKp46, msNKp46, | Mortality and lung virus load of IAV-infected NCR1gfp/gfp knockout mice is dose-dependent. NCR1 is not protective at high IAV challenge doses. IAV NA impairs sialic acid-dependent HA recognition of NKp46-Ig. Inhibition of NA activity augments HA binding of NKp46-Ig. Blockade of NA augments NK cell cytolytic activity. NA treatment of IAV-infected NCR1gfp/gfp mice reduces mortality. | Bar-On et al. 2013 Cell Rep. [23] |
| A/Texas/1/1977 (H3N2) | ||||
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | hNKp46, msNKp46, hNKp44, hNKp30, KIR2DS4, KIR2DL1 | NA hinders NCR interactions with IAV HA. Inhibition of IAV NA activity increases NK cytolytic function. | Bar-On et al. 2014 J. Infect.Dis. [34] |
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | msNKp46 | Analysis of NCR1 glycosylation status. Identified occupied O-linked glycosites at Thr222 and Thr225. T225A and T225A mutation reduced lectin binding. Reported that O-linked glycosites are crucial for interactions with IAV HA and subsequent NK cytolytic activity. | Glasner et al. 2015 Cell Disc. [35] |
| Candida glabrata | BG2 | hNKp46 NCR1 | Candida glabrata adhesins Epa1, Epa6, and Epa7 (all of which are lectins) engage with hNKp46 and NCR1. Fungal clearance was impaired in NCR knockout mice. | Vitenshtein et al. 2016 Cell Host & Microbe [62] |
| Influenza A virus | A/Puerto Rico/8/1934 (H1N1) | hNKp46 | mAb blockade of hNKp46 inhibits NKp46 mediated NK cytolytic activity. Inhibition of NA activity increased HA binding to NKp46-Ig. HA binding to NA-treated NKp46-Ig was reduced. Demonstrate that HA also binds the 2B4 and NTB-A SLAM-family receptors in a sialic-acid dependent manner. | Duev-Cohen et al. 2016 Oncotarget [36] |
| A/Brisbane/59/2007 (H1N1) | ||||
| Human metapneumovirus | Human metapneumovirus (strain not reported) | hNKp46, msNKp46 | HMPV-infection induces expression of NKp46-Ig/NCR1-Ig ligand(s) in vitro. Blockade of HMPV and NKp46-Ig/anti-NCR1 interaction with anti-NKp46/anti-NCR1 mAb reduced NKp46-mediated NK cytolytic activity. Despite this, HMPV proteins do not interact directly with NKp46-Ig/NCR1-Ig. NCR1 controls HMPV virus load in vivo. | Diab et al. 2017 Eur. J. Immunol. [41] |
| Human respirovirus 3 Influenza A virus | Human pararainfluenza virus 3 (strain C-243) A/Puerto Rico/8/1934 (H1N1) | hNKp46, hNKp44 | HPIV3 HN induces NKp44 surface expression on CD14+ monocytes more potently than IAV HA. The opposite holds true for NKp46 expression. This effect can be abrogated by anti-HN or anti-HA antibody treatment. | McQuaid et al. 2018 [37] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luczo, J.M.; Ronzulli, S.L.; Tompkins, S.M. Influenza A Virus Hemagglutinin and Other Pathogen Glycoprotein Interactions with NK Cell Natural Cytotoxicity Receptors NKp46, NKp44, and NKp30. Viruses 2021, 13, 156. https://doi.org/10.3390/v13020156
Luczo JM, Ronzulli SL, Tompkins SM. Influenza A Virus Hemagglutinin and Other Pathogen Glycoprotein Interactions with NK Cell Natural Cytotoxicity Receptors NKp46, NKp44, and NKp30. Viruses. 2021; 13(2):156. https://doi.org/10.3390/v13020156
Chicago/Turabian StyleLuczo, Jasmina M., Sydney L. Ronzulli, and Stephen M. Tompkins. 2021. "Influenza A Virus Hemagglutinin and Other Pathogen Glycoprotein Interactions with NK Cell Natural Cytotoxicity Receptors NKp46, NKp44, and NKp30" Viruses 13, no. 2: 156. https://doi.org/10.3390/v13020156
APA StyleLuczo, J. M., Ronzulli, S. L., & Tompkins, S. M. (2021). Influenza A Virus Hemagglutinin and Other Pathogen Glycoprotein Interactions with NK Cell Natural Cytotoxicity Receptors NKp46, NKp44, and NKp30. Viruses, 13(2), 156. https://doi.org/10.3390/v13020156