
Phage phiKZ—The First of Giants

 ,
,
Abstract
1. Introduction
2. Isolation and Growth Features of phiKZ
3. Structure of the phiKZ Phage Particle
4. Phage PhiKZ Transduction
5. Bacteriophage phiKZ Genome
6. Origin of the Group of Phylogenetically Related Species of P. aeruginosa Giant Phages and Their Species Diversity
7. Antibacterial Activity of P. aeruginosa Giant Phages and Features of Their Use in Phage Therapy
8. New Giant Phages of Pseudomonas aeruginosa and Other Bacterial Species
9. The Future of PhiKZ-like Phages—Basic Science and Practical Application
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krylov, V.N.; Zhazykov, I. Pseudomonas bacteriophage phiKZ-possible model for studying the genetic control of morphogenesis. Genetika 1978, 14, 678–685. [Google Scholar] [PubMed]
- Mesyanzhinov, V.V.; Robben, J.; Grymonprez, B.; Kostyuchenko, V.A.; Bourkaltseva, M.V.; Sykilinda, N.N.; Krylov, V.N.; Volckaert, G. The genome of bacteriophage φKZ of Pseudomonas aeruginosa. J. Mol. Biol. 2002, 317, 1–19. [Google Scholar] [CrossRef]
- Bradley, D.E. The Structure of Coliphages. J. Gen. Microbiol. 1963, 31, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Yap, M.L.; Rossmann, M.G. Structure and function of bacteriophage T4. Futur. Microbiol. 2014, 9, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.-W. Bacteriophage Electron Microscopy. Adv. Clin. Chem. 2012, 82, 1–32. [Google Scholar] [CrossRef]
- Forterre, P. Viruses in the 21st Century: From the Curiosity-Driven Discovery of Giant Viruses to New Concepts and Definition of Life. Clin. Infect. Dis. 2017, 65, S74–S79. [Google Scholar] [CrossRef] [PubMed]
- Brandes, N.; Linial, M. Giant Viruses—Big Surprises. Viruses 2019, 11, 404. [Google Scholar] [CrossRef]
- Hendrix, R.W. Jumbo Bacteriophages. Curr. Top. Microbiol. Immun. 2009, 328, 229–240. [Google Scholar] [CrossRef]
- Cornelissen, A.; Hardies, S.C.; Shaburova, O.V.; Krylov, V.N.; Mattheus, W.; Kropinski, A.M.; Lavigne, R. Complete Genome Sequence of the Giant Virus OBP and Comparative Genome Analysis of the Diverse KZ-Related Phages. J. Virol. 2011, 86, 1844–1852. [Google Scholar] [CrossRef]
- Brown, J.M.; LaBarre, B.A.; Hewson, I. Characterization of Trichodesmium-associated viral communities in the eastern Gulf of Mexico. FEMS Microbiol. Ecol 2013, 84, 603–613. [Google Scholar] [CrossRef][Green Version]
- Luo, Z.-H.; Yu, Y.-P.; Jost, G.; Xu, W.; Huang, X.-L. Complete genome sequence of a giant Vibrio bacteriophage VH7D. Mar. Genom. 2015, 24, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Jun, J.W.; Giri, S.S.; Yun, S.; Kim, H.J.; Kang, J.W.; Han, S.J.; Jeong, D.; Park, S.C. Isolation and characterisation of pVa-21, a giant bacteriophage with anti-biofilm potential against Vibrio alginolyticus. Sci. Rep. 2019, 9, 6284. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Giri, S.S.; Yun, S.; Kim, H.J.; Kang, J.W.; Han, S.J.; Kwon, J.; Jun, J.W.; Oh, W.T.; Park, S.C.; et al. Genomic characterization of bacteriophage pEt-SU, a novel phiKZ-related virus infecting Edwardsiella tarda. Arch. Virol. 2019, 165, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, S.T.; Redzuan, N.H.M.; Barton, M.K.; Amin, N.A.M.; Desmond, M.I.; Adams, L.E.; Ali, B.; Pardo, S.; Molleur, D.; Wu, W.; et al. Global Proteomic Profiling ofSalmonellaInfection by a Giant Phage. J. Virol. 2018, 93, e01833-18. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.; Ely, B. Analyses of four new Caulobacter Phicbkviruses indicate independent lineages. J. Gen. Virol. 2019, 100, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Lavysh, D.; Sokolova, M.L.; Minakhin, L.; Yakunina, M.; Artamonova, T.O.; Kozyavkin, S.; Makarova, K.S.; Koonin, E.V.; Severinov, K. The genome of AR9, a giant transducing Bacillus phage encoding two multisubunit RNA polymerases. Virology 2016, 495, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Kurochkina, L.P.; Semenyuk, P.I.; Sykilinda, N.N.; Miroshnikov, K.A. The unique two-component tail sheath of giant Pseudomonas phage PaBG. Virology 2018, 515, 46–51. [Google Scholar] [CrossRef]
- Khoa, H.V.; Midorikawa, Y.; Uchino, T.; Nakai, T.; Kato, G.; Kondo, H.; Hirono, I.; Labaiden, M.; Direkbusarakom, S.; Sano, M. Complete Genome Sequence of the Lytic Giant Bacteriophage pT24 Infecting Tenacibaculum spp., Isolated from a Shrimp Culture Pond. Genome Announc. 2017, 5, e00081-17. [Google Scholar] [CrossRef]
- Krylov, V.N.; Smirnova, T.A.; Rebentish, B.A.; Minenkova, I.B. Stucture of PhiKZ bacteriophage particles. Vopr. Virusol. 1978, 5, 568–571. [Google Scholar]
- Tiaglov, B.V.; Krylov, V.N.; Plotnikova, T.G.; Minaev, V.E.; Permogorov, V.I. Certain physico-chemical properties of bacteriophage phiKZ. Mol. Biol. 1980, 14, 1019–1022. [Google Scholar]
- Plotnikova, T.G.; Dzhusupova, A.B.; Khrenova, E.A.; Krylov, V.N. Genetic and pheno-genetic studies of the group of ts-mutants of Pseudomonas aeruginosa PAO1 phiKZ phage. Genetika 1982, 18, 1793–1798. [Google Scholar] [PubMed]
- Dzhusupova, A.B.; Plotnikova, T.G.; Krylov, V.N. Detection of transduction by virulent bacteriophage phi KZ of Pseudomo-nas aeruginosa chromosomal markers in the presence of plasmid RMS148. Genetika 1982, 18, 1799–1802. [Google Scholar] [PubMed]
- Smirnova, T.A.; Minenkova, I.B.; Khrenova, E.A.; Plotnikova, T.G.; Krylov, V.N. Electron microscope study of the intracel-lular development of the Pseudomonas aeruginosa bacteriophage phi KZ. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 1983, 5, 25–28. [Google Scholar]
- Krylov, V.N.; Smirnova, T.A.; Minenkova, I.B.; Plotnikova, T.G.; Zhazikov, I.Z.; Khrenova, E.A. Pseudomonas bacteriophage contains an inner body in its capsid. Can. J. Microbiol. 1984, 30, 758–762. [Google Scholar] [CrossRef]
- Khrenova, E.A.; Akhverdyan, V.Z.; Krylov, V.N. Relation of Pseudomonas aeruginosa bacteriophages phiKZ and 21 processing the unique nucleoprotein structure in heads. Genetika 1985, 11, 31–37. [Google Scholar]
- Sharibjanova, T.O.; Akhverdyan, V.Z.; Krylov, V.N. A comparative study of DNA homology and morphology of Pseudomonas aeruginosa bacteriophages to reveal phylogenetic relationships and for an express-classification. Genetika 1992, 28, 24–32. [Google Scholar]
- Krylov, V.N.; Tolmachova, T.O.; Akhverdian, V.Z. DNA homology in species of bacteriophages active on Pseudomonas aeruginosa. Arch. Virol. 1993, 131, 141–151. [Google Scholar] [CrossRef]
- Burkal’tseva, M.V.; Krylov, V.N.; Pleteneva, E.A.; Shaburova, O.V.; Krylov, S.V.; Volkart, G.; Sykilinda, N.N.; Kurochkina, L.P.; Mesianzhinov, V.V. Phenogenetic characterization of a group of giant Phi KZ-like bacteriophages of Pseudomonas aeruginosa. Russ. J. Genet. 2002, 38, 1470–1479. [Google Scholar] [CrossRef]
- Miller, E.S.; Heidelberg, J.F.; Eisen, J.A.; Nelson, W.C.; Durkin, A.S.; Ciecko, A.; Feldblyum, T.V.; White, O.; Paulsen, I.T.; Nierman, W.C.; et al. Complete Genome Sequence of the Broad-Host-Range Vibriophage KVP40: Comparative Genomics of a T4-Related Bacteriophage. J. Bacteriol. 2003, 185, 5220–5233. [Google Scholar] [CrossRef]
- Krylov, V.N.; Bourkaltseva, M.V.; Sykilinda, N.N.; Pleteneva, E.A.; Shaburova, O.V.; Kadykov, V.A.; Miller, S.; Biebl, M. Comparisons of the Genomes of New Giant Phages Isolated from Environmental Pseudomonas aeruginosa Strains of Different Regions. Russ. J. Genet. 2004, 40, 363–368. [Google Scholar] [CrossRef]
- Hertveldt, K.; Lavigne, R.; Pleteneva, E.; Sernova, N.; Kurochkina, L.; Korchevskii, R.; Robben, J.; Mesyanzhinov, V.; Krylov, V.N.; Volckaert, G. Genome Comparison of Pseudomonas aeruginosa Large Phages. J. Mol. Biol. 2005, 354, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-C.; Chen, C.-R.; Lin, J.-W.; Shen, G.-H.; Chang, K.-M.; Tseng, Y.-H.; Weng, S.-F. Isolation and Characterization of Novel Giant Stenotrophomonas maltophilia Phage φSMA5. Appl. Environ. Microbiol. 2005, 71, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Kiljunen, S.; Hakala, K.; Pinta, E.; Huttunen, S.; Pluta, P.; Gador, A.; Lönnberg, H.; Skurnik, M. Yersiniophage ϕR1-37 is a tailed bacteriophage having a 270 kb DNA genome with thymidine replaced by deoxyuridine. Microbiology 2005, 151, 4093–4102. [Google Scholar] [CrossRef] [PubMed]
- Shaburova, O.V.; Hertveldt, K.; de la Crus, D.M.; Krylov, S.V.; Pleteneva, E.A.; Burkaltseva, M.V.; Lavigne, R.; Volcaert, G.; Krylov, V.N. Comparison of new giant bacteriophages OBP and Lu11 of soil pseudomonads with bacteriophages of phiKZ-supergroup of Pseudomonas aeruginosa. Genetika 2006, 42, 1065–1074. [Google Scholar] [PubMed]
- Thomas, J.A.; Rolando, M.R.; Carroll, C.A.; Shen, P.S.; Belnap, D.M.; Weintraub, S.T.; Serwer, P.; Hardies, S.C. Characterization of Pseudomonas chlororaphis myovirus 201ϕ2-1 via genomic sequencing, mass spectrometry, and electron microscopy. Virology 2008, 376, 330–338. [Google Scholar] [CrossRef]
- Serwer, P.; Hayes, S.J.; Thomas, J.A.; Hardies, S.C. Propagating the missing bacteriophages: A large bacteriophage in a new class. Virol. J. 2007, 4, 21. [Google Scholar] [CrossRef]
- Doermann, A.H. Lysis and Lysis Inhibition with Escherichia coli Bacteriophage. J. Bacteriol. 1948, 55, 257–276. [Google Scholar] [CrossRef]
- Pleteneva, E.A.; Krylov, S.V.; Shaburova, O.V.; Bourkal’Tseva, M.V.; Miroshnikov, K.A.; Krylov, V.N. Pseudolysogeny of Pseudomonas aeruginosa bacteria infected with φKZ-like bacteriophages. Russ. J. Genet. 2010, 46, 20–25. [Google Scholar] [CrossRef]
- Fokine, A.; Kostyuchenko, V.A.; Efimov, A.V.; Kurochkina, L.P.; Sykilinda, N.N.; Robben, J.; Volckaert, G.; Hoenger, A.; Chipman, P.R.; Battisti, A.J.; et al. A Three-dimensional Cryo-electron Microscopy Structure of the Bacteriophage ϕKZ Head. J. Mol. Biol. 2005, 352, 117–124. [Google Scholar] [CrossRef]
- Fokine, A.; Battisti, A.J.; Bowman, V.D.; Efimov, A.V.; Kurochkina, L.P.; Chipman, P.R.; Mesyanzhinov, V.V.; Rossmann, M.G. Cryo-EM Study of the Pseudomonas Bacteriophage φKZ. Structure 2007, 15, 1099–1104. [Google Scholar] [CrossRef]
- Wu, W.; Thomas, J.A.; Cheng, N.; Black, L.; Steven, A. Bubblegrams Reveal the Inner Body of Bacteriophage ϕKZ. Science 2012, 335, 182. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A.; Weintraub, S.T.; Wu, W.; Winkler, D.C.; Cheng, N.; Steven, A.C.; Black, L.W. Extensive proteolysis of head and inner body proteins by a morphogenetic protease in the giantPseudomonas aeruginosaphage φKZ. Mol. Microbiol. 2012, 84, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Monson, R.; Foulds, I.; Foweraker, J.; Welch, M.; Salmond, G.P.C. The Pseudomonas aeruginosa generalized transducing phage φPA3 is a new member of the φKZ-like group of ‘jumbo’ phages, and infects model laboratory strains and clinical isolates from cystic fibrosis patients. Microbiology 2011, 157, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Miroshnikov, K.A.; Faizullina, N.M.; Sykilinda, N.N.; Mesyanzhinov, V.V. Properties of the endolytic transglycosylase encoded by gene 144 of Pseudomonas aeruginosa bacteriophage phiKZ. Biochemistry (Moscow) 2006, 71, 300–305. [Google Scholar] [CrossRef]
- Yakunina, M.; Artamonova, T.; Borukhov, S.; Makarova, K.S.; Severinov, K.; Minakhin, L. A non-canonical multisubunit RNA polymerase encoded by a giant bacteriophage. Nucleic Acids Res. 2015, 43, 10411–10420. [Google Scholar] [CrossRef]
- Orekhova, M.; Koreshova, A.; Artamonova, T.O.; Khodorkovskii, M.; Yakunina, M.V. The study of the phiKZ phage non-canonical non-virion RNA polymerase. Biochem. Biophys. Res. Commun. 2019, 511, 759–764. [Google Scholar] [CrossRef]
- Chaikeeratisak, V.; Nguyen, K.; Egan, M.E.; Erb, M.L.; Vavilina, A.; Pogliano, J. The Phage Nucleus and Tubulin Spindle Are Conserved among Large Pseudomonas Phages. Cell Rep. 2017, 20, 1563–1571. [Google Scholar] [CrossRef]
- Danilova, Y.A.; Belousova, V.V.; Moiseenko, A.V.; Vishnyakov, I.E.; Yakunina, M.V.; Sokolova, O.S. Maturation of Pseudo-Nucleus Compartment in P. aeruginosa, Infected with Giant phiKZ Phage. Viruses 2020, 12, 1197. [Google Scholar] [CrossRef]
- Mendoza, S.D.; Nieweglowska, E.S.; Govindarajan, S.; Leon, L.M.; Berry, J.D.; Tiwari, A.; Chaikeeratisak, V.; Pogliano, J.; Agard, D.A.; Bondy-Denomy, J. A bacteriophage nucleus-like compartment shields DNA from CRISPR nucleases. Nat. Cell Biol. 2020, 577, 244–248. [Google Scholar] [CrossRef]
- Aylett, C.H.; Izoré, T.; Amos, L.A.; Löwe, J. Structure of the Tubulin/FtsZ-Like Protein TubZ from Pseudomonas Bacteriophage ΦKZ. J. Mol. Biol. 2013, 425, 2164–2173. [Google Scholar] [CrossRef]
- Kazlauskas, D.; Venclovas, Č. Computational analysis of DNA replicases in double-stranded DNA viruses: Relationship with the genome size. Nucleic Acids Res. 2011, 39, 8291–8305. [Google Scholar] [CrossRef] [PubMed]
- Kazlauskas, D.; Krupovic, M.; Venclovas, Č. The logic of DNA replication in double-stranded DNA viruses: Insights from global analysis of viral genomes. Nucleic Acids Res. 2016, 44, 4551–4564. [Google Scholar] [CrossRef]
- Kazlauskas, D.; Krupovic, M.; Guglielmini, J.; Forterre, P.; Venclovas, Č. Diversity and evolution of B-family DNA polymerases. Nucleic Acids Res. 2020, 48, 10142–10156. [Google Scholar] [CrossRef] [PubMed]
- Sau, K.; Sau, S.; Mandal, S.C.; Ghosh, T.C. Factors Influencing the Synonymous Codon and Amino Acid Usage Bias in AT-rich Pseudomonas aeruginosa Phage PhiKZ. Acta Biochim. Biophys. Sin. 2005, 37, 625–633. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lecoutere, E.; Ceyssens, P.-J.; Miroshnikov, K.A.; Mesyanzhinov, V.V.; Krylov, V.N.; Noben, J.-P.; Robben, J.; Hertveldt, K.; Volckaert, G.; Lavigne, R. Identification and comparative analysis of the structural proteomes of ϕKZ and EL, two giant Pseudomonas aeruginosa bacteriophages. Proteomics 2009, 9, 3215–3219. [Google Scholar] [CrossRef] [PubMed]
- De Smet, J.; Zimmermann, M.; Kogadeeva, M.; Ceyssens, P.-J.; Vermaelen, W.; Blasdel, B.; Jang, H.B.; Sauer, U.; Lavigne, R. High coverage metabolomics analysis reveals phage-specific alterations to Pseudomonas aeruginosa physiology during infection. ISME J. 2016, 10, 1823–1835. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, R.B.; Latta, R.L. Phage Typing of Pseudomonas aeruginosai Clinical and Epidemiologic Considerations. J. Infect. Dis. 1974, 130, S33–S42. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.; Sutton, L. Restriction-modification systems determined by Pseudomonas plasmids. Plasmid 1982, 8, 141–147. [Google Scholar] [CrossRef]
- Jacoby, G.A.; Sutton, L. Properties of plasmids responsible for production of extended-spectrum beta-lactamases. Antimicrob. Agents Chemother. 1991, 35, 164–169. [Google Scholar] [CrossRef]
- Ackermann, H.-W.; Dubow, M.S.; Jarvis, A.W.; Jones, L.A.; Krylov, V.N.; Maniloff, J.; Rocourt, J.; Safferman, R.S.; Schneider, J.; Seldin, L.; et al. The species concept and its application to tailed phages. Arch. Virol. 1992, 124, 69–82. [Google Scholar] [CrossRef]
- Krylov, V.N.; Cruz, D.M.D.; Hertveldt, K.; Ackermann, H.-W. “φKZ-like viruses”, a proposed new genus of myovirus bacteriophages. Arch. Virol. 2007, 152, 1955–1959. [Google Scholar] [CrossRef] [PubMed]
- Krylov, V.N.; Miroshnikov, K.A.; Krylov, S.V.; Veyko, V.P.; Pleteneva, E.A.; Shaburova, O.V.; Bourkal’Tseva, M.V. Interspecies migration and evolution of bacteriophages of the genus phiKZ: The purpose and criteria of the search for new phiKZ-like bacteriophages. Russ. J. Genet. 2010, 46, 138–145. [Google Scholar] [CrossRef]
- Tétart, F.; Desplats, C.; Kutateladze, M.; Monod, C.; Ackermann, H.-W.; Krisch, H.M. Phylogeny of the Major Head and Tail Genes of the Wide-Ranging T4-Type Bacteriophages. J. Bacteriol. 2001, 183, 358–366. [Google Scholar] [CrossRef]
- Krylov, V.N. Phage Therapy in Terms of Bacteriophage Genetics: Hopes, Prospects, Safety, Limitations. Russ. J. Genet. 2001, 37, 715–730. [Google Scholar] [CrossRef]
- Krylov, V.; Shaburova, O.; Pleteneva, E.; Krylov, S.; Kaplan, A.; Burkaltseva, M.; Polygach, O.; Chesnokova, E. Selection of phages and conditions for the safe phage therapy against Pseudomonas aeruginosa infections. Virol. Sin. 2015, 30, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Krylov, V.N. Bacteriophages of Pseudomonas aeruginosa. Adv. Clin. Chem. 2014, 88, 227–278. [Google Scholar] [CrossRef]
- Pleteneva, E.A.; Shaburova, O.V.; Burkaltseva, M.V.; Krylov, S.V.; Kaplan, A.M.; Chesnokova, E.N.; Polygach, O.A.; Voroshilova, N.N.; Mikhailova, N.A.; Zverev, V.V.; et al. Novel approach to composition of, bacteriophage mixtures for antibacterial therapy. J. Microbiol. Epidemiol. Immunobiol. 2016, 5, 3–11. [Google Scholar]
- Jacoby, G.A. AmpC beta-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef]
- Ceyssens, P.J.; Minakhin, L.; Van den Bossche, A.; Yakunina, M.; Klimuk, E.; Blasdel, B.; De Smet, J.; Noben, J.P.; Bläsi, U.; Severinov, K.; et al. Development of giant bacteriophage ϕKZ is independent of the host transcription apparatus. J. Virol. 2014, 88, 10501–10510. [Google Scholar] [CrossRef]
- Warren, R.J.; Bose, S.K. Bacteriophage-induced Inhibition of Host Functions 1. J. Virol. 1968, 2, 327–334. [Google Scholar] [CrossRef]
- Krylov, S.V.; Pleteneva, E.A.; Bourkaltseva, M.V.; Shaburova, O.V.; Miroshnikov, K.A.; Lavigne, R.; Cornelissen, A.; Krylov, V.N. Genome instability of Pseudomonas aeruginosa phages of the EL species: Examination of virulent mutants. Russ. J. Genet. 2011, 47, 162–167. [Google Scholar] [CrossRef]
- Krylov, V.N. The role of horizontal gene transfer by bacteriophages in the origin of pathogenic bacteria. Russ. J. Genet. 2003, 39, 483–504. [Google Scholar] [CrossRef]
- Yuan, Y.; Gao, M. Jumbo Bacteriophages: An Overview. Front. Microbiol. 2017, 8, 403. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, E.M.; Mattheus, W.; Cornelissen, A.; Shaburova, O.; Krylov, V.N.; Kropinski, A.M.; Lavigne, R. Complete genome sequence of the giant Pseudomonas phage Lu11. J. Virol. 2012, 86, 6369–6370. [Google Scholar] [CrossRef] [PubMed]
- Sykilinda, N.N.; Bondar, A.A.; Gorshkova, A.S.; Kurochkina, L.P.; Kulikov, E.E.; Shneider, M.M.; Kadykov, V.A.; Solovjeva, N.V.; Kabilov, M.R.; Mesyanzhinov, V.V.; et al. Complete Genome Sequence of the Novel Giant Pseudomonas Phage PaBG. Genome Announc. 2014, 2. [Google Scholar] [CrossRef]
- Buttimer, C.; Born, Y.; Lucid, A.; Loessner, M.J.; Fieseler, L.; Coffey, A. Erwinia amylovora phage vB_EamM_Y3 represents another lineage of hairy Myoviridae. Res. Microbiol. 2018, 169, 505–514. [Google Scholar] [CrossRef]
- Pfreundt, U.; Spungin, D.; Hou, S.; Voß, B.; Bermanfrank, I.; Hess, W.R. Genome of a giant bacteriophage from a decaying Trichodesmium bloom. Mar. Genom. 2017, 33, 21–25. [Google Scholar] [CrossRef]
- Simoliūnas, E.; Kaliniene, L.; Truncaitė, L.; Zajančkauskaitė, A.; Staniulis, J.; Kaupinis, A.; Ger, M.; Valius, M.; Meškys, R. Klebsiella phage vB_KleM-RaK2—A giant singleton virus of the family Myoviridae. PLoS ONE 2013, 8, e60717. [Google Scholar] [CrossRef]
- Sullivan, M.B.; Coleman, M.L.; Weigele, P.; Rohwer, F.; Chisholm, S.W. Three Prochlorococcus Cyanophage Genomes: Signature Features and Ecological Interpretations. PLoS Biol. 2005, 3, e144. [Google Scholar] [CrossRef]
- Lee, J.-H.; Bai, J.; Shin, H.; Kim, Y.; Park, B.; Heu, S.; Ryu, S. A Novel Bacteriophage Targeting Cronobacter sakazakii Is a Potential Biocontrol Agent in Foods. Appl. Environ. Microbiol. 2015, 82, 192–201. [Google Scholar] [CrossRef]
- Chen, L.; Yuan, S.; Liu, Q.; Mai, G.; Yang, J.; Deng, D.; Zhang, B.; Liu, C.; Ma, Y. In Vitro Design and Evaluation of Phage Cocktails Against Aeromonas salmonicida. Front. Microbiol. 2018, 9, 1476. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, O.S.; Shaburova, O.; Pechnikova, E.; Shaytan, A.; Krylov, S.; Kiselev, N.; Krylov, V.N. Genome packaging in EL and Lin68, two giant phiKZ-like bacteriophages of P. aeruginosa. Virology 2014, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Semenyuk, P.I.; Orlov, V.N.; Sokolova, O.S.; Kurochkina, L.P. New GroEL-like chaperonin of bacteriophage OBP Pseudomonas fluorescens suppresses thermal protein aggregation in an ATP-dependent manner. Biochem. J. 2016, 473, 2383–2393. [Google Scholar] [CrossRef] [PubMed]
- Stanishneva-Konovalova, T.B.; Semenyuk, P.I.; Kurochkina, L.P.; Pichkur, E.B.; Vasilyev, A.L.; Kovalchuk, M.V.; Kirpichnikov, M.P.; Sokolova, O.S. Cryo-EM reveals an asymmetry in a novel single-ring viral chaperonin. J. Struct. Biol. 2020, 209, 107439. [Google Scholar] [CrossRef]
- Wälti, M.A.; Steiner, J.; Meng, F.; Chung, H.S.; Louis, J.M.; Ghirlando, R.; Tugarinov, V.; Nath, A.; Clore, G.M. Probing the mechanism of inhibition of amyloid-β(1–42)–induced neurotoxicity by the chaperonin GroEL. Proc. Natl. Acad. Sci. USA 2018, 115, E11924–E11932. [Google Scholar] [CrossRef]
- Wälti, M.A.; Schmidt, T.; Murray, D.T.; Wang, H.; Hinshaw, J.E.; Clore, G.M. Chaperonin GroEL accelerates protofibril formation and decorates fibrils of the Het-s prion protein. Proc. Natl. Acad. Sci. USA 2017, 114, 9104–9109. [Google Scholar] [CrossRef]
- Drulis-Kawa, Z.; Olszak, T.; Danis, K.; Majkowska-Skrobek, G.; Ackermann, H.-W. A giant Pseudomonas phage from Poland. Arch. Virol. 2013, 159, 567–572. [Google Scholar] [CrossRef]
- Abbasifar, R.; Griffiths, M.W.; Sabour, P.M.; Ackermann, H.-W.; Vandersteegen, K.; Lavigne, R.; Noben, J.-P.; Villa, A.A.; Abbasifar, A.; Nash, J.H.; et al. Supersize me: Cronobacter sakazakii phage GAP32. Virology 2014, 460, 138–146. [Google Scholar] [CrossRef]
- Caubilla-Barron, J.; Hurrell, E.; Townsend, S.; Cheetham, P.; Loc-Carrillo, C.; Fayet, O.; Prère, M.-F.; Forsythe, S. Genotypic and Phenotypic Analysis of Enterobacter sakazakii Strains from an Outbreak Resulting in Fatalities in a Neonatal Intensive Care Unit in France. J. Clin. Microbiol. 2007, 45, 3979–3985. [Google Scholar] [CrossRef]
- Kabanova, A.P.; Shneider, M.M.; Korzhenkov, A.A.; Bugaeva, E.N.; Miroshnikov, K.K.; Zdorovenko, E.L.; Kulikov, E.E.; Toschakov, S.V.; Ignatov, A.N.; Knirel, Y.A.; et al. Host Specificity of the Dickeya Bacteriophage PP35 Is Directed by a Tail Spike Interaction with Bacterial O-Antigen, Enabling the Infection of Alternative Non-pathogenic Bacterial Host. Front. Microbiol. 2019, 9, 3288. [Google Scholar] [CrossRef]
- Czajkowski, R. May the Phage be with You? Prophage-Like Elements in the Genomes of Soft Rot Pectobacteriaceae: Pectobacterium spp. and Dickeya spp. Front. Microbiol. 2019, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.K.; Van Der Wolf, J.M.; Saddler, G.S.; Lojkowska, E.; Helias, V.; Pirhonen, M.; Lahkin, L.T.; Elphinstone, J.G. Dickeya species: An emerging problem for potato production in Europe. Plant Pathol. 2011, 60, 385–399. [Google Scholar] [CrossRef]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, V.D.; Al-Abdely, H.M.; El-Kholy, A.A.; Alkhawaja, S.A.A.; Leblebicioglu, H.; Mehta, Y.; Rai, V.; Hung, N.V.; Kanj, S.S.; Salama, M.F.; et al. International Nosocomial Infection Control Consortium report, data summary of 50 countries for 2010-2015: Device-associated module. Am. J. Infect. Control. 2016, 44, 1495–1504. [Google Scholar] [CrossRef]
- Pires, D.P.; Costa, A.R.; Pinto, G.; Meneses, L.; Azeredo, J. Current challenges and future opportunities of phage therapy. FEMS Microbiol. Rev. 2020, 44, 684–700. [Google Scholar] [CrossRef]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162–173. [Google Scholar] [CrossRef]
- Rohde, C.; Resch, G.; Pirnay, J.P.; Blasdel, B.G.; Debarbieux, L.; Gelman, D.; Górski, A.; Hazan, R.; Huys, I.; Kakabadze, E. Expert Opinion on Three Phage Therapy Related Topics: Bacterial Phage Resistance, Phage Training and Prophages in Bacterial Production Strains. Viruses 2018, 10, 178. [Google Scholar] [CrossRef]
- São-José, C. Engineering of Phage-Derived Lytic Enzymes: Improving Their Potential as Antimicrobials. Antibiotics 2018, 7, 29. [Google Scholar] [CrossRef]
- Fenton, M.; McAuliffe, O.; O’Mahony, J.; Coffey, A. Recombinant bacteriophage lysins as antibacterials. Bioeng. Bugs 2010, 1, 9–16. [Google Scholar] [CrossRef]
- Breidenstein, E.B.; De La Fuente-Nunez, C.; Hancock, R.E. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa Biofilm, a Programmed Bacterial Life for Fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Latz, S.; Krüttgen, A.; Häfner, H.; Buhl, E.M.; Ritter, K.; Horz, H.-P. Differential Effect of Newly Isolated Phages Belonging to PB1-Like, phiKZ-Like and LUZ24-Like Viruses against Multi-Drug Resistant Pseudomonas aeruginosa under Varying Growth Conditions. Viruses 2017, 9, 315. [Google Scholar] [CrossRef] [PubMed]
- Can, K.; Aksu, U.; Yenen, O.Ş. Investigation of PhiKZ phage therapy against Pseudomonas aeruginosa in mouse pneumonia model. Turk. J. Med Sci. 2018, 48, 670–678. [Google Scholar] [CrossRef]
- Danis-Wlodarczyk, K.; Vandenheuvel, D.; Jang, H.B.; Briers, Y.; Olszak, T.; Arabski, M.; Wasik, S.; Drabik, M.; Higgins, G.; Tyrrell, J.; et al. A proposed integrated approach for the preclinical evaluation of phage therapy in Pseudomonas infections. Sci. Rep. 2016, 6, 28115. [Google Scholar] [CrossRef] [PubMed]
- Polygach, O.A.; Dabizheva, A.N.; Voroshilova, N.N. Effect of the Composition of Lytic Bacteriophages of P. aeruginosa Formation and Destruction of Bacterial Biofilms. Epidemiol. Vaccinal Prev. 2018, 17, 20–25. [Google Scholar] [CrossRef][Green Version]
- Paradis-Bleau, C.; Cloutier, I.; Lemieux, L.; Sanschagrin, F.; Laroche, J.; Auger, M.; Garnier, A.; Levesque, R.C. Peptidoglycan lytic activity of the Pseudomonas aeruginosa phage phiKZ gp144 lytic transglycosylase. FEMS Microbiol. Lett. 2007, 266, 201–209. [Google Scholar] [CrossRef]
- Briers, Y.; Walmagh, M.; Lavigne, R. Use of bacteriophage endolysin EL188 and outer membrane permeabilizers against Pseudomonas aeruginosa. J. Appl. Microbiol. 2011, 110, 778–785. [Google Scholar] [CrossRef]
- Walmagh, M.; Briers, Y.; Dos Santos, S.B.; Azeredo, J.; Lavigne, R. Characterization of Modular Bacteriophage Endolysins from Myoviridae Phages OBP, 201φ2-1 and PVP-SE1. PLoS ONE 2012, 7, e36991. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Scientific Research |
|---|---|
| 1978 | - First description of the structure and some biological properties of the new virulent Pseudomonas phage phiKZ with unique features distinguishing this phage from all those known before, including a large size of the head and DNA, and the presence in the head of an unusual complicated structural formation of a cylindrical shape named the “central or inner body” [1,19]. |
| 1980–1982 | - The results, obtained with tailless mutants of phage phiKZ, allowed for the circular location of the fibrous material around the “inner body” to be proven. It was shown that the genetic map of phiKZ is circular, while the dsDNA has a block structure, and the GC pair content is equal to 44% [20,21]. - It was shown that phiKZ is a general transducing phage [22]. |
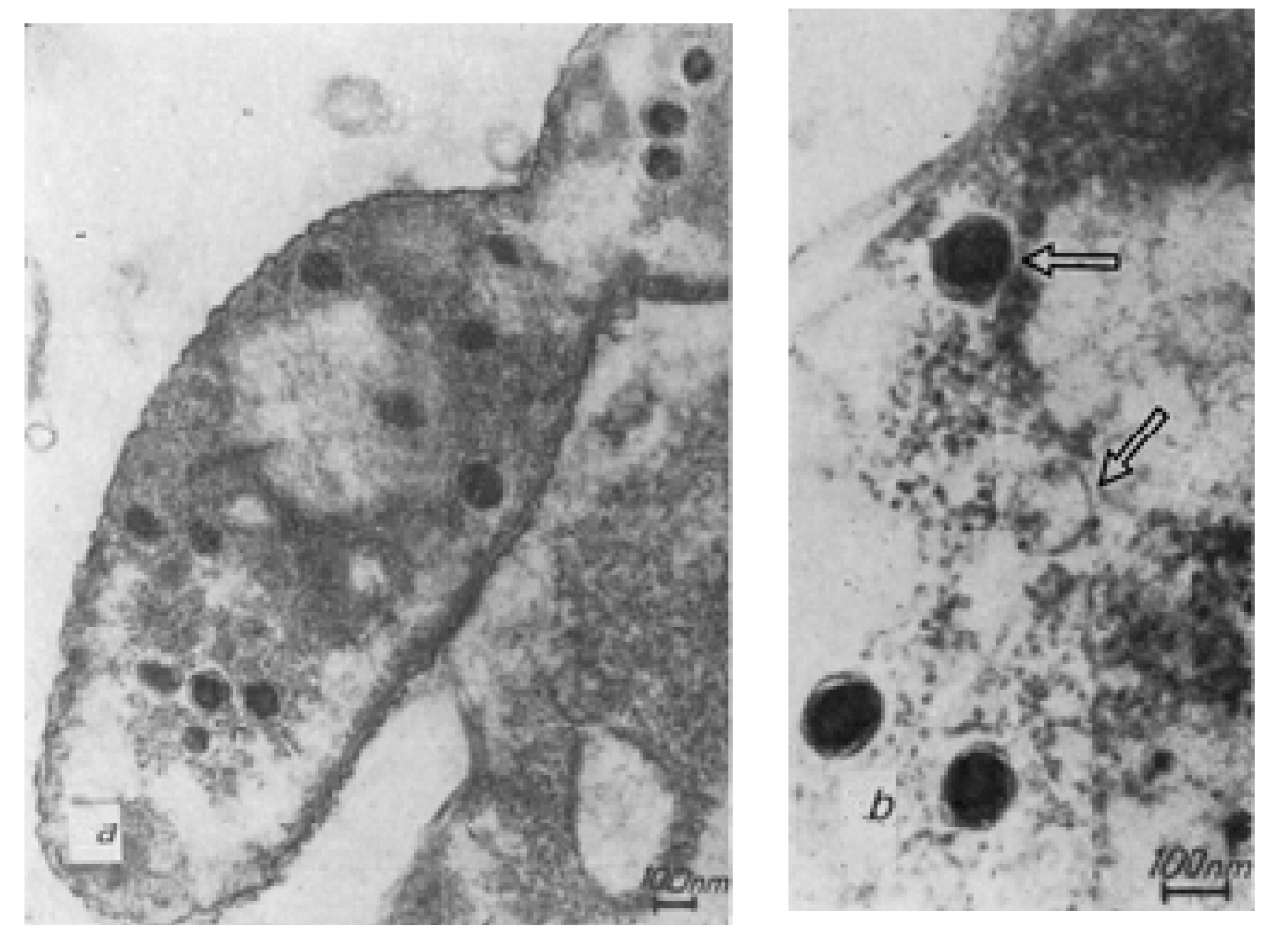
| 1983–1984 | - A detailed electron microscopical examination of phage phiKZ confirmed that in the center of the phage head is a cylinder of low electron density (“inner body”) coated by fibrinous material which is packaged around the inner body in a spool-like manner. These structures disappear after the adsorption of phage particles in bacteria. On studying the maturation of the phiKZ bacteriophage in infected Pseudomonas aeruginosa bacteria, successive stages of changes in the structure of phage particles after infection were observed. The mechanism for the packaging and ejection of phiKZ DNA was proposed [23,24]. |
| 1985 | - It was shown that the Lin21 (later known as phiLin21) bacteriophage (from the Lindbergh collection, with a large phage particle), contains in its capsid a central body similar in structure to that of phiKZ and is related to phage phiKZ. It was considered that the phages were representatives of a group of related phiKZ-like phages [25]. |
| 1992–1993 | - It was shown that bacteriophage Lin68 (later known as phiLin68) forms opalescent negative colonies (typical of phiKZ-related phages) similar to phage phiKZ in genome size and morphology of phage particle, but with a shorter tail. Phage Lin68 was assigned to another morphotype and another species [26,27]. |
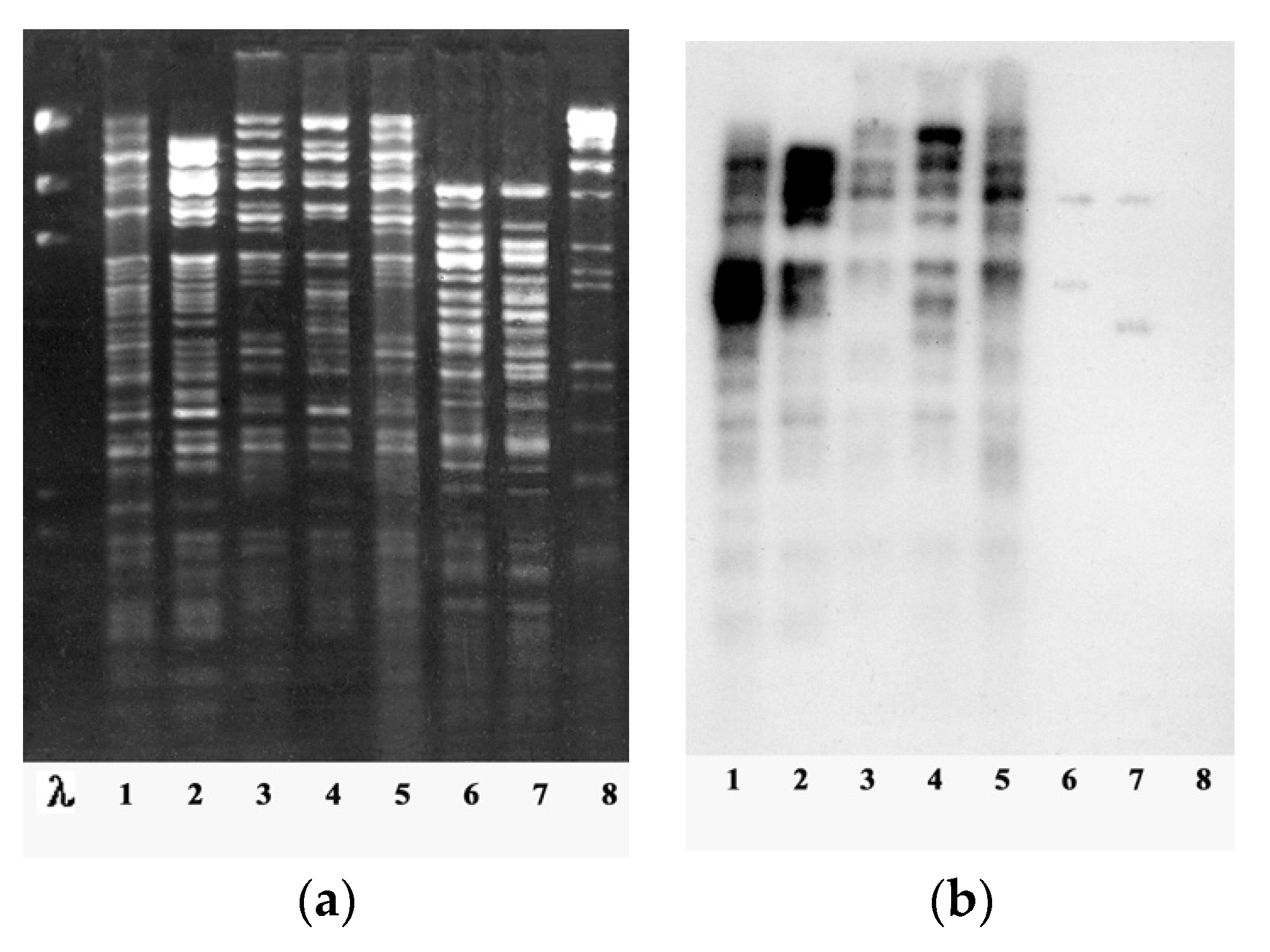
| 2002 | - The complete nucleotide sequence of phiKZ DNA and a preliminary analysis of its genome structure were presented. The genome contains 280,334 bp, and has 306 ORFs and 6 tRNAs. It was the largest bacteriophage genome to be fully sequenced at that time [2] - A comparative study was made of a group of P. aeruginosa virulent giant DNA bacteriophages similar to phage phiKZ with regard to several genetic and phenotypic properties. For all phages a broad spectrum of lytic activity was shown, as well as the ability to overcome the suppressing effect of plasmids and the capability for general transduction and pseudolysogeny. By DNA homology the phages were assigned to three species (represented by phages phiKZ, Lin68, and EL (later known as phiEL), respectively) and two new genera (phiKZ and EL) [28]. - For the first time, the term “giant phages” was used to denote a group of bacteriophages similar to phiKZ (with unusually large capsids and genome sizes). |
| 2003 | - The complete genome sequence of the T4-like broad-host-range vibriophage KVP40 was determined. The KVP40 genome is 244,835 bp in size and is related to the phage T4 genome [29]. |
| 2004 | - A study of nine phiKZ-like giant phages isolated from natural sources was carried out. It was concluded that phages of the phiKZ species were common in natural populations of various regions, while EL- and Lin68-related phages were extremely rare. Phages isolated in different geographical regions showed no substantial differences [30]. |
| 2005 | - The complete nucleotide sequence of second phiKZ-like giant phage EL was presented. The EL genome comprises 211,215 bp and has 201 predicted ORFs. The EL genome does not share its DNA sequence homology with any other viruses and micro-organisms sequenced to date. However, one-third of the predicted EL gene products share a similarity with phiKZ proteins [31]. |
| 2005–2008 | - New data on the isolation and study of new giant bacteriophages active on different species and genera of bacteria appeared with regard to phiSMA5 (Stenotrophomonas maltophilia) [32], piR1-37 (Yersinia enterocolitica) [33], Lu11 (P. putida var. Manila) and OBP (P. fluorescens) [34], 201varphi2-1 (P. chlororaphis) [35], and 0305phi8-36 (Bacillus thuringiensis) [36]. |
| 2009 | - For tailed bacteriophages with genomes of more than 200 kbp of DNA the term “jumbo phages” was proposed [8]. |
| Since 2010 | - Mass sequencing of bacteriophage genomes began, with detection of phages with a large genome size active on different bacteria species. Studies on the possibilities of using giant bacteriophages for antibacterial therapy have been carried out. |
| Name | Source And Geographical Location Of The Phage Isolation | Capsid Size (D, nm) | Tail size (L/W, nm) | Genome Size, bp; (NCBI Reference Sequence, if Available) | Growth on Plasmid-Containing P. aeruginosa | ||
|---|---|---|---|---|---|---|---|
| PAO303 (Rms148) * | PAO38 (PMG53) * | PAO1 Mutants with Plasmids * Rms165, Rpl11, Rip64, Pmg73, Rms139, Pmg1, Pmg35, Rms149, Rms163 | |||||
| phages of the phiKZ species | |||||||
| phiKZ | sputum of a patient with pulmonary infection (Kazakhstan) | 120 | 180/20 | 280,334; (NC_004629.1) | - | - | + |
| phiLin21 | Lindberg Pseudomonas aeruginosa typing phage collection [57] were kindly transferred us by Prof. H-W. Ackermann, (Canada) | 120 | 180/20 | ~290,000 | - | - | + |
| phiLBG20 phiLBG21 phiLBG23 phiLBG26 | water sources of Moscow and Moscow region (Russia) | ||||||
| phiPBD1 phiPBD2 phiPBD3 phiPBD4 | Danube and its tributary (Germany) | ||||||
| PTB80 | commercial phage mix produced in Tbilisi (Georgia) | ||||||
| phiNN | commercial phage mix produced in Nizhnii Novgorod (Russia) | ||||||
| phiChe2/2 ** | infected post-burn wound of patient in Burn Department of Chelyabinsk hospital (Russia) | ||||||
| phages of the Lin68 species | |||||||
| phiLin68 | Lindberg Pseudomonas aeruginosa typing phage collection [26] were kindly transferred us by Prof. H-W. Ackermann, (Canada) | 120 | 160/20 | ~290,000 | + | - | + |
| phiLBG22 | water sources of Moscow region (Russia) | ||||||
| phages of the EL species | |||||||
| phiEL | water sources of Moscow region (Russia) | 120 | 180/20 | 211,215 (NC_007623.1) | - | - | + |
| phiRU | water sources of Moscow region (Russia) | 120 | 180/20 | ~220,000 | - | - | + |
| phiChe21/5 (later named as phiCHE) ** | infected post-burn wound of patient in Burn Department of Chelyabinsk hospital (Russia) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krylov, V.; Bourkaltseva, M.; Pleteneva, E.; Shaburova, O.; Krylov, S.; Karaulov, A.; Zhavoronok, S.; Svitich, O.; Zverev, V. Phage phiKZ—The First of Giants. Viruses 2021, 13, 149. https://doi.org/10.3390/v13020149
Krylov V, Bourkaltseva M, Pleteneva E, Shaburova O, Krylov S, Karaulov A, Zhavoronok S, Svitich O, Zverev V. Phage phiKZ—The First of Giants. Viruses. 2021; 13(2):149. https://doi.org/10.3390/v13020149
Chicago/Turabian StyleKrylov, Victor, Maria Bourkaltseva, Elena Pleteneva, Olga Shaburova, Sergey Krylov, Alexander Karaulov, Sergey Zhavoronok, Oxana Svitich, and Vitaly Zverev. 2021. "Phage phiKZ—The First of Giants" Viruses 13, no. 2: 149. https://doi.org/10.3390/v13020149
APA StyleKrylov, V., Bourkaltseva, M., Pleteneva, E., Shaburova, O., Krylov, S., Karaulov, A., Zhavoronok, S., Svitich, O., & Zverev, V. (2021). Phage phiKZ—The First of Giants. Viruses, 13(2), 149. https://doi.org/10.3390/v13020149