
Serum Neutralization Profiles of Straw-Colored Fruit Bats (Eidolon helvum) in Makurdi (Nigeria), against Four Lineages of Lagos Bat Lyssavirus


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
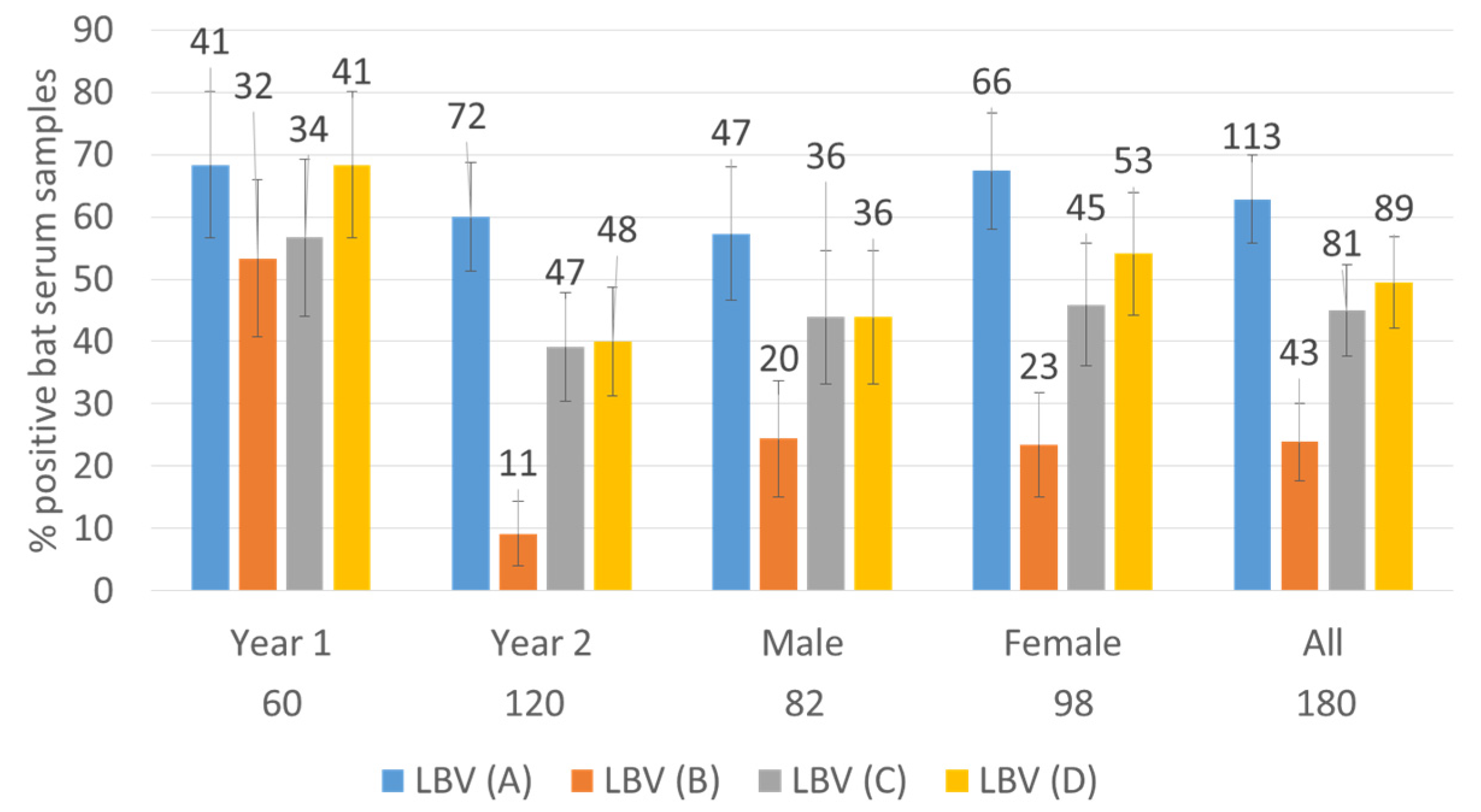
3. Results
3.1. Serum Samples Neutralized at Least One Lineage of LBV
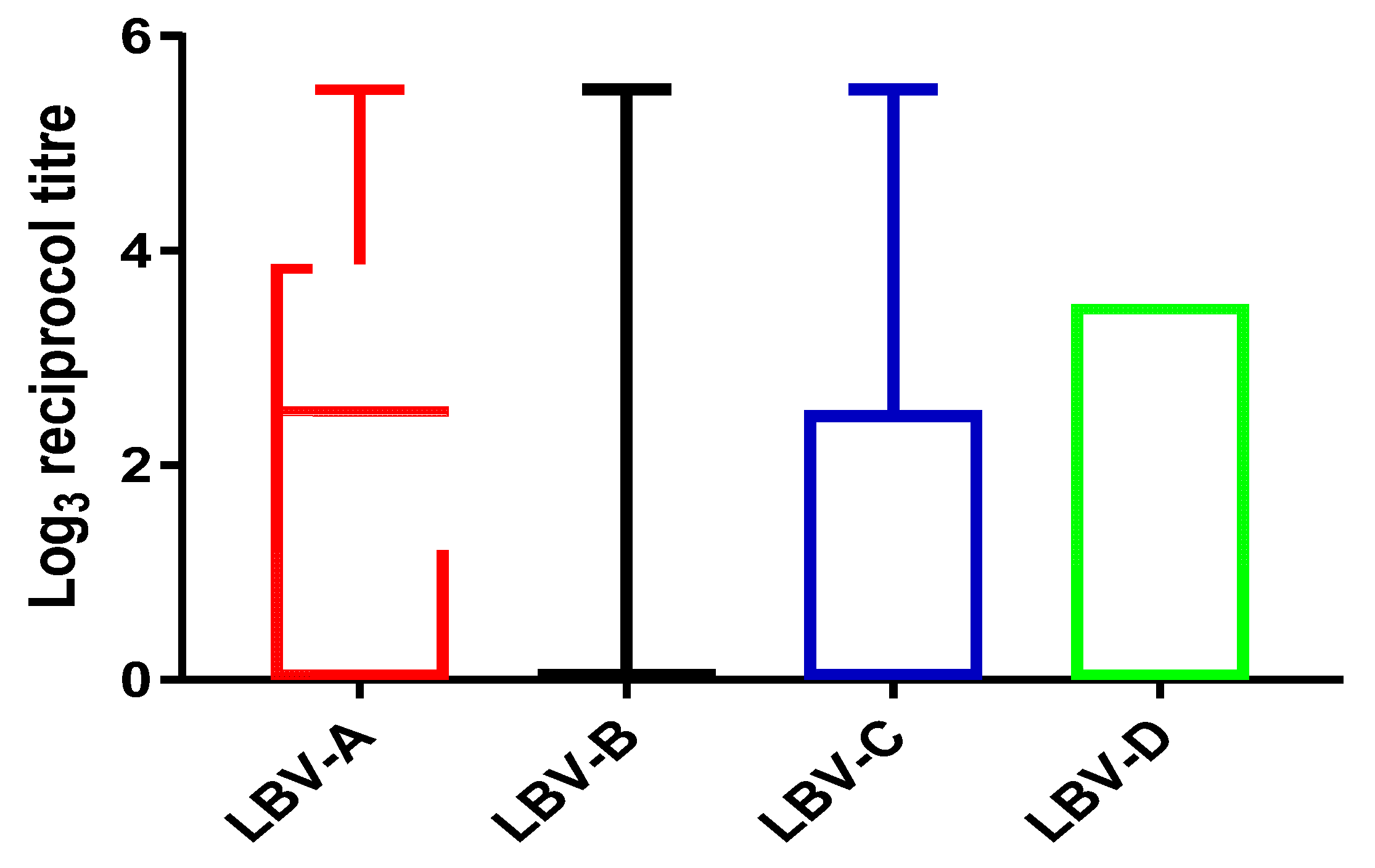
3.2. Titer Distribution against Different Lineages of LBV
3.3. Serum Samples Neutralized Only One LBV Lineage
3.4. Serum Samples Neutralized Multiple Lineages of LBV
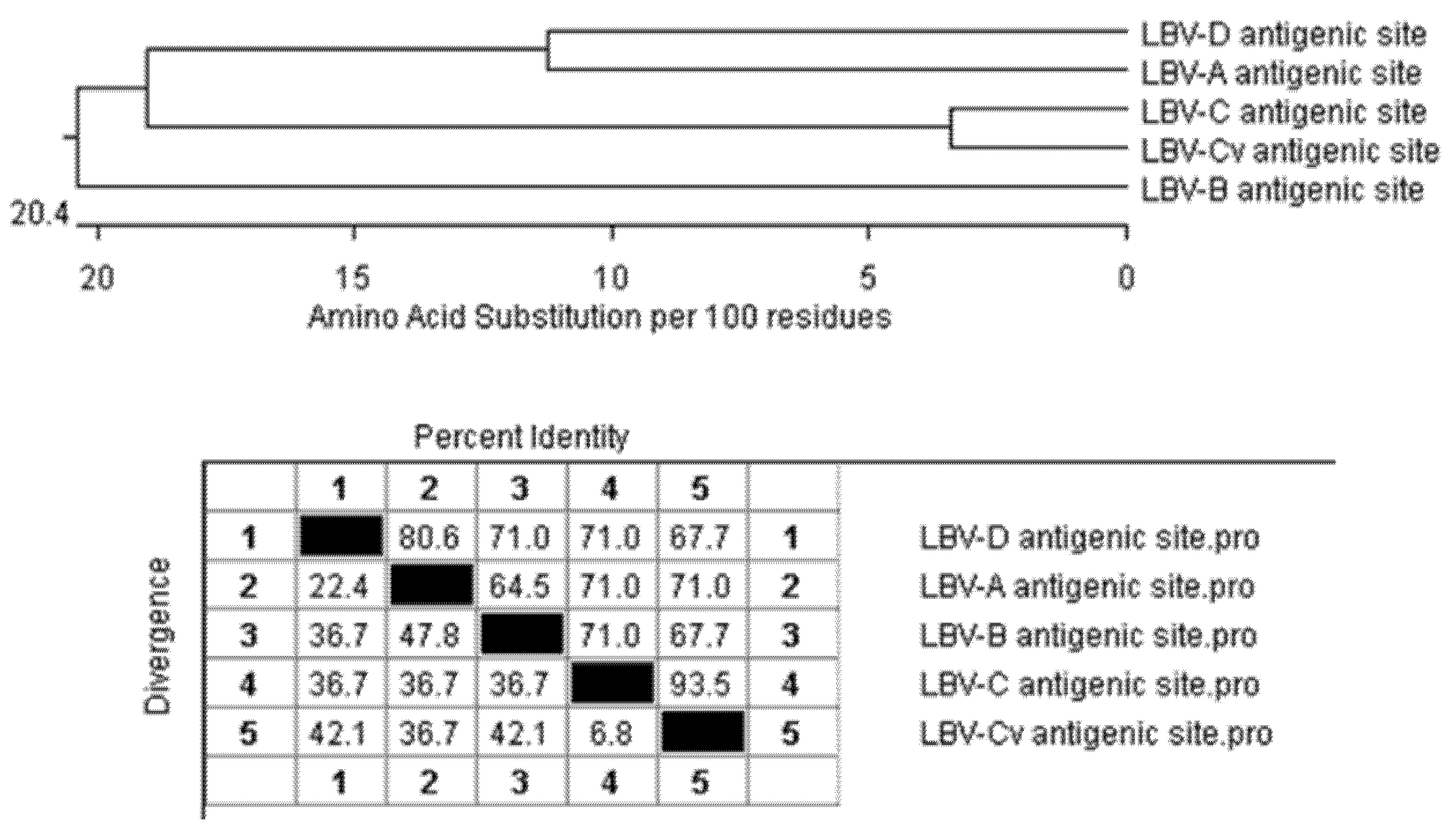
3.5. Antigenic Cartography
3.6. Sequence Similarities among LBV Lineages
4. Discussion
4.1. Evidence Supports the Presence of LBV-A in Nigeria
4.2. Serum Neutralization against LBV-B
4.3. Serum Neutralization against LBV-C and LBV-D
4.4. Cross-Neutralization among LBV Lineages
4.5. Other Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuhn, J.H.; Adkins, S.; Agwanda, B.R.; Al Kubrusli, R.; Alkhovsky, S.V.; Amarasinghe, G.K.; Avsic-Zupanc, T.; Ayllon, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. 2021 Taxonomic update of phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2021, 163, 1–54. [Google Scholar] [CrossRef]
- Boulger, L.R.; Porterfield, J.S. Isolation of a virus from Nigerian fruit bats. Trans. R. Soc. Trop. Med. Hyg. 1958, 52, 421–424. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Markotter, W.; Beagley, J.C.; Urazova, O.Y.; Breiman, R.F.; Rupprecht, C.E. Lagos bat virus in Kenya. J. Clin. Microbiol. 2008, 46, 1451–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markotter, W.; Kuzmin, I.; Rupprecht, C.E.; Nel, L.H. Phylogeny of Lagos bat virus: Challenges for lyssavirus taxonomy. Virus Res. 2008, 135, 10–21. [Google Scholar] [CrossRef]
- Coertse, J.; Geldenhuys, M.; le Roux, K.; Markotter, W. Lagos Bat Virus, an Under-Reported Rabies-Related Lyssavirus. Viruses 2021, 13, 576. [Google Scholar] [CrossRef] [PubMed]
- Banyard, A.C.; Hayman, D.; Johnson, N.; McElhinney, L.; Fooks, A.R. Bats and lyssaviruses. Adv. Virus Res. 2011, 79, 239–289. [Google Scholar] [CrossRef]
- Begeman, L.; Suu-Ire, R.; Banyard, A.C.; Drosten, C.; Eggerbauer, E.; Freuling, C.M.; Gibson, L.; Goharriz, H.; Horton, D.L.; Jennings, D.; et al. Experimental Lagos bat virus infection in straw-colored fruit bats: A suitable model for bat rabies in a natural reservoir species. PLoS Negl. Trop. Dis. 2020, 14, e0008898. [Google Scholar] [CrossRef] [PubMed]
- Suu-Ire, R.; Begeman, L.; Banyard, A.C.; Breed, A.C.; Drosten, C.; Eggerbauer, E.; Freuling, C.M.; Gibson, L.; Goharriz, H.; Horton, D.L.; et al. Pathogenesis of bat rabies in a natural reservoir: Comparative susceptibility of the straw-colored fruit bat (Eidolon helvum) to three strains of Lagos bat virus. PLoS Negl. Trop. Dis. 2018, 12, e0006311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayman, D.T.; Fooks, A.R.; Rowcliffe, J.M.; McCrea, R.; Restif, O.; Baker, K.S.; Horton, D.L.; Suu-Ire, R.; Cunningham, A.A.; Wood, J.L. Endemic Lagos bat virus infection in Eidolon helvum. Epidemiol. Infect. 2012, 140, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- Suu-Ire, R.D.; Fooks, A.R.; Banyard, A.C.; Selden, D.; Amponsah-Mensah, K.; Riesle, S.; Ziekah, M.Y.; Ntiamoa-Baidu, Y.; Wood, J.L.N.; Cunningham, A.A. Lagos Bat Virus Infection Dynamics in Free-Ranging Straw-Colored Fruit Bats (Eidolon helvum). Trop. Med. Infect. Dis. 2017, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Freuling, C.M.; Binger, T.; Beer, M.; Adu-Sarkodie, Y.; Schatz, J.; Fischer, M.; Hanke, D.; Hoffmann, B.; Hoper, D.; Mettenleiter, T.C.; et al. Lagos bat virus transmission in an Eidolon helvum bat colony, Ghana. Virus Res. 2015, 210, 42–45. [Google Scholar] [CrossRef]
- Mebatsion, T.; Cox, J.H.; Frost, J.W. Isolation and characterization of 115 street rabies virus isolates from Ethiopia by using monoclonal antibodies: Identification of 2 isolates as Mokola and Lagos bat viruses. J. Infect. Dis. 1992, 166, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Mayer, A.E.; Niezgoda, M.; Markotter, W.; Agwanda, B.; Breiman, R.F.; Rupprecht, C.E. Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Res. 2010, 149, 197–210. [Google Scholar] [CrossRef]
- Markotter, W.; Coertse, J.; De Vries, L.; Geldenhuys, M.; Mortlock, M. Bat-borne viruses in Africa: A critical review. J. Zool (1987) 2020, 311, 77–98. [Google Scholar] [CrossRef]
- Weyer, J.; Szmyd-Potapczuk, A.V.; Blumberg, L.H.; Leman, P.A.; Markotter, W.; Swanepoel, R.; Paweska, J.T.; Nel, L.H. Epidemiology of human rabies in South Africa, 1983-2007. Virus Res. 2011, 155, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, C.A.; Kuzmin, I.V.; Blanton, J.D.; Weldon, W.C.; Manangan, J.S.; Rupprecht, C.E. Efficacy of rabies biologics against new lyssaviruses from Eurasia. Virus Res. 2005, 111, 44–54. [Google Scholar] [CrossRef]
- Cliquet, F.; Aubert, M.; Sagne, L. Development of a fluorescent antibody virus neutralisation test (FAVN test) for the quantitation of rabies-neutralising antibody. J. Immunol. Methods 1998, 212, 79–87. [Google Scholar] [CrossRef]
- Brookes, S.M.; Aegerter, J.N.; Smith, G.C.; Healy, D.M.; Jolliffe, T.A.; Swift, S.M.; Mackie, I.J.; Pritchard, J.S.; Racey, P.A.; Moore, N.P.; et al. European bat lyssavirus in Scottish bats. Emerg. Infect. Dis. 2005, 11, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Lapedes, A.S.; de Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Osterhaus, A.D.; Fouchier, R.A. Mapping the antigenic and genetic evolution of influenza virus. Science 2004, 305, 371–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shipley, R.; Wright, E.; Lean, F.Z.X.; Selden, D.; Horton, D.L.; Fooks, A.R.; Banyard, A.C. Assessing Rabies Vaccine Protection against a Novel Lyssavirus, Kotalahti Bat Lyssavirus. Viruses 2021, 13, 947. [Google Scholar] [CrossRef] [PubMed]
- Horton, D.L.; McElhinney, L.M.; Marston, D.A.; Wood, J.L.; Russell, C.A.; Lewis, N.; Kuzmin, I.V.; Fouchier, R.A.; Osterhaus, A.D.; Fooks, A.R.; et al. Quantifying antigenic relationships among the lyssaviruses. J. Virol. 2010, 84, 11841–11848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.S.; Horton, D.L.; Easton, A.J.; Fooks, A.R.; Banyard, A.C. Rabies virus vaccines: Is there a need for a pan-lyssavirus vaccine? Vaccine 2012, 30, 7447–7454. [Google Scholar] [CrossRef]
- Dzikwi, A.A.; Kuzmin, I.I.; Umoh, J.U.; Kwaga, J.K.; Ahmad, A.A.; Rupprecht, C.E. Evidence of Lagos bat virus circulation among Nigerian fruit bats. J. Wildl. Dis. 2010, 46, 267–271. [Google Scholar] [CrossRef] [Green Version]
- Hayman, D.T.; Fooks, A.R.; Horton, D.; Suu-Ire, R.; Breed, A.C.; Cunningham, A.A.; Wood, J.L. Antibodies against Lagos bat virus in megachiroptera from West Africa. Emerg. Infect. Dis. 2008, 14, 926–928. [Google Scholar] [CrossRef]
- Vora, N.M.; Osinubi, M.O.V.; Davis, L.; Abdurrahman, M.; Adedire, E.B.; Akpan, H.; Aman-Oloniyo, A.F.; Audu, S.W.; Blau, D.; Dankoli, R.S.; et al. Bat and Lyssavirus Exposure among Humans in Area that Celebrates Bat Festival, Nigeria, 2010 and 2013. Emerg. Infect. Dis. 2020, 26, 1399–1408. [Google Scholar] [CrossRef]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Conrardy, C.; Ruone, S.; Kuzmin, I.V.; Guo, X.; Tao, Y.; Niezgoda, M.; Haynes, L.; Agwanda, B.; Breiman, R.F.; et al. Detection of novel SARS-like and other coronaviruses in bats from Kenya. Emerg. Infect. Dis. 2009, 15, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, P.L.; Firth, C.; Street, C.; Henriquez, J.A.; Petrosov, A.; Tashmukhamedova, A.; Hutchison, S.K.; Egholm, M.; Osinubi, M.O.; Niezgoda, M.; et al. Identification of a severe acute respiratory syndrome coronavirus-like virus in a leaf-nosed bat in Nigeria. mBio 2010, 1, e00208-10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, K.; Selleck, P.; Hooper, P.; Hyatt, A.; Gould, A.; Gleeson, L.; Westbury, H.; Hiley, L.; Selvey, L.; Rodwell, B.; et al. A morbillivirus that caused fatal disease in horses and humans. Science 1995, 268, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Bellini, W.J.; Rota, P.A.; Harcourt, B.H.; Tamin, A.; Lam, S.K.; Ksiazek, T.G.; Rollin, P.E.; Zaki, S.R.; Shieh, W.; et al. Nipah virus: A recently emergent deadly paramyxovirus. Science 2000, 288, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Muller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Cottontail, V.M.; Rasche, A.; Yordanov, S.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Delicat, A.; Paweska, J.T.; Gonzalez, J.P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esona, M.D.; Mijatovic-Rustempasic, S.; Conrardy, C.; Tong, S.; Kuzmin, I.V.; Agwanda, B.; Breiman, R.F.; Banyai, K.; Niezgoda, M.; Rupprecht, C.E.; et al. Reassortant group A rotavirus from straw-colored fruit bat (Eidolon helvum). Emerg. Infect. Dis. 2010, 16, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.H.; Cheng, P.K.; Lai, M.Y.; Leung, P.C.; Wong, K.K.; Lee, W.Y.; Lim, W.W. Virulence potential of fusogenic orthoreoviruses. Emerg. Infect. Dis. 2012, 18, 944–948. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| °% Positives (95% Confidence Intervals) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cut-Off | Both Years | Year 1 | Year 2 | Male | Female | |||||
| 9 | 74 | (67–80) | 82 | (72–91) | 70 | (62–78) | 71 | (61–81) | 78 | (69–86) |
| 27 | 52 | (44–59) | 62 | (49–74) | 47 | (38–56) | 45 | (34–56) | 57 | (47–67) |
| Total No. | 180 | 120 | 60 | 82 | 98 | |||||
| Cut Off | Lineages | A | B | C | D | Total |
|---|---|---|---|---|---|---|
| 9 | No. of sample | 23 | 0 | 2 | 8 | 33 |
| Titer range | 9–140.3 | n/a | 9 | 9–46.77 | ||
| 27 | No. of sample | 24 | 1 | 3 | 7 | 35 |
| Titer range | 27–140.3 | 27 | 27–81 | 27–243 |
| Cut-Off | Lineages | AB | AC | AD | BC | BD | CD | Total |
|---|---|---|---|---|---|---|---|---|
| 9 | No. of sample | 4 | 14 | 15 | 0 | 1 | 4 | 38 |
| Titer range | 9–140.3 | 9–420.89 | 15.95–140.3 | n/a | 15.99 | 9–140.3 | ||
| 27 | No. of sample | 0 | 3 | 16 | 0 | 0 | 2 | 21 |
| Titer range | n/a | 46.7–34,091.96 | 27–140.3 | n/a | n/a | 27–81 |
| Cut-Off | Lineages | BCD (Not A) | ACD (Not B) | ABD (Not C) | ABC (Not D) | ABCD |
|---|---|---|---|---|---|---|
| 9 | No. of sample | 5 | 24 | 1 | 1 | 31 |
| Highest titer | 243 | 11,363.89 | 46.77 | 420.89 | 420.89 | |
| 27 | No. of sample | 3 | 16 | 3 | 2 | 13 |
| Highest titer | 46.7 | 11,363.99 | 140.3 | 420.89 | 729 |
| Site IIb (34–42) | Site IIa (198–200) | Site I (226–231) | Site IV (263–264) | Site III (330–338) | Site ‘a’ (342–343) | |
|---|---|---|---|---|---|---|
| LBV-A | GCSETSSFT | RKA | TLCGKP | NR | KRVDNWVDI | KG |
| LBV-B | GCGTSSVFS | KKS | TLCGKP | NR | LKVDNWSEI | KG |
| LBV-C | GCSDTATFS | KKS | TLCGKP | NR | LRVDSWNDI | KG |
| LBV-Cv | GCSNTATFN | KKS | TLCGKP | NR | LRVDSWNDI | KG |
| LBV-D | GCSTSTSFS | RKA | TLCGKP | NR | RRVDNWTDI | KG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ameh, V.O.; Wu, G.; Goharriz, H.; Shipley, R.; Fooks, A.R.; Sabeta, C.T.; McElhinney, L.M. Serum Neutralization Profiles of Straw-Colored Fruit Bats (Eidolon helvum) in Makurdi (Nigeria), against Four Lineages of Lagos Bat Lyssavirus. Viruses 2021, 13, 2378. https://doi.org/10.3390/v13122378
Ameh VO, Wu G, Goharriz H, Shipley R, Fooks AR, Sabeta CT, McElhinney LM. Serum Neutralization Profiles of Straw-Colored Fruit Bats (Eidolon helvum) in Makurdi (Nigeria), against Four Lineages of Lagos Bat Lyssavirus. Viruses. 2021; 13(12):2378. https://doi.org/10.3390/v13122378
Chicago/Turabian StyleAmeh, Veronica Odinya, Guanghui Wu, Hooman Goharriz, Rebecca Shipley, Anthony R. Fooks, Claude T. Sabeta, and Lorraine M. McElhinney. 2021. "Serum Neutralization Profiles of Straw-Colored Fruit Bats (Eidolon helvum) in Makurdi (Nigeria), against Four Lineages of Lagos Bat Lyssavirus" Viruses 13, no. 12: 2378. https://doi.org/10.3390/v13122378
APA StyleAmeh, V. O., Wu, G., Goharriz, H., Shipley, R., Fooks, A. R., Sabeta, C. T., & McElhinney, L. M. (2021). Serum Neutralization Profiles of Straw-Colored Fruit Bats (Eidolon helvum) in Makurdi (Nigeria), against Four Lineages of Lagos Bat Lyssavirus. Viruses, 13(12), 2378. https://doi.org/10.3390/v13122378