
Omnipresence of Partitiviruses in Rice Aggregate Sheath Spot Symptom-Associated Fungal Isolates from Paddies in Thailand

 , , ,
, , , 
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Collection, Culture, and Maintenance
2.2. Taxonomic Analysis of Fungal Species
2.3. dsRNA Extraction
2.4. cDNA Library Construction, RT-PCR, and RLM-RACE
2.5. Next-Generation Sequencing (NGS) Approach
2.6. Bioinformatic Analysis
2.7. Virus Curing
2.8. Detection of Partitiviruses by RT-PCR
2.9. Pathogenicity Test
3. Results
3.1. Determination of Fungal Species
3.2. Screening of Fungal Isolates for Viral dsRNAs
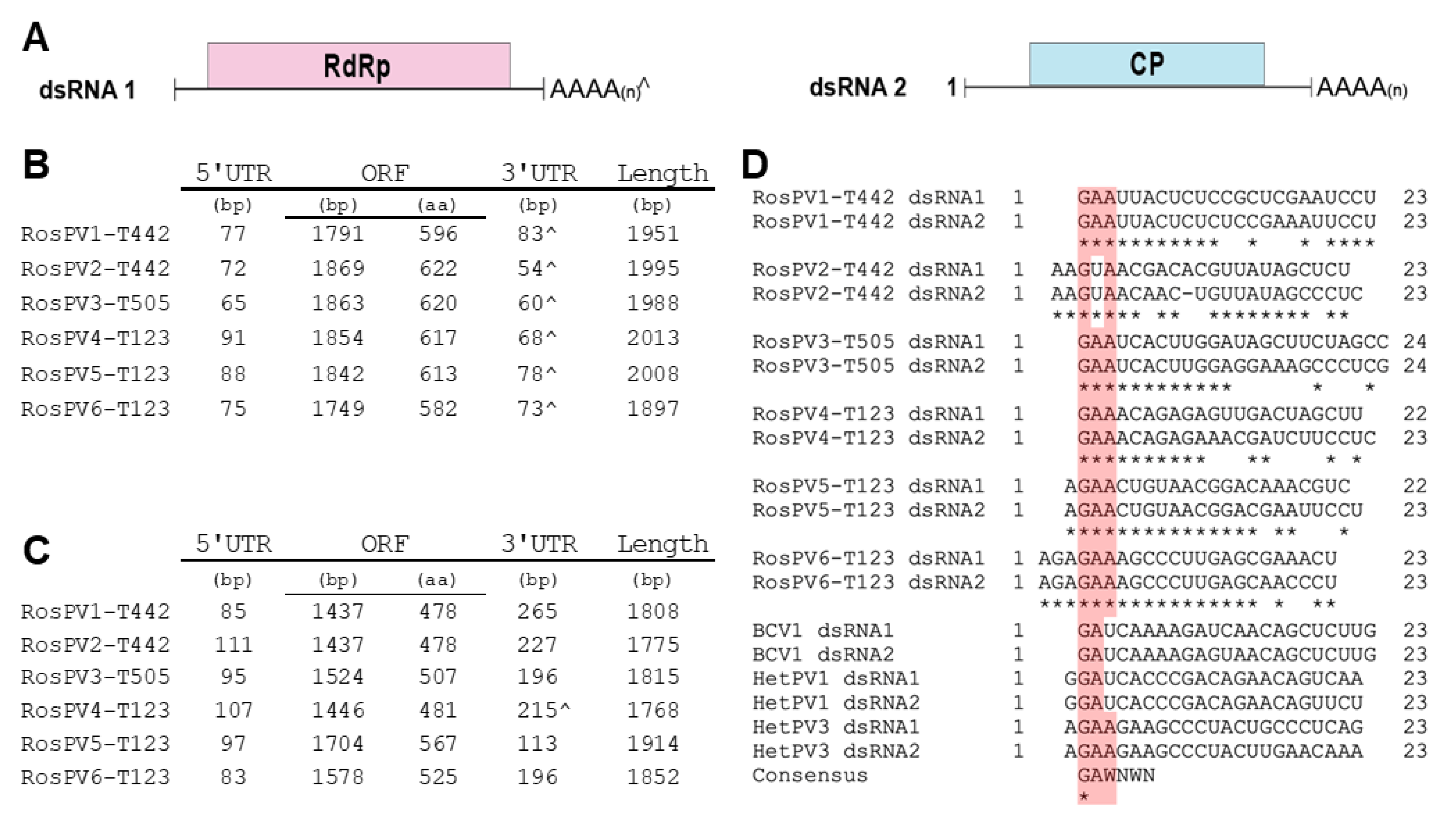
3.3. Genomic Organization of Analyzed dsRNA Elements
3.4. BLAST Search with Deduced Amino Acid Sequences of Viral Proteins
3.5. Phylogenetic Analysis of Partitiviruses
3.6. RT-PCR Based Detection of Partitiviruses from Fungal Isolates Used in This Study
3.7. Other Partitiviruses Coinfecting R. oryzae and R. oryzae-sativae
3.8. Isogenic Virus-Cured Isolates
4. Discussion
4.1. Multiple Infections of Partitiviruses in Rhizoctonia spp.
4.2. Widespread Nature of the Partitiviruses Found in This Study
4.3. Possible Viral Transmission through Hyphal Contact
4.4. Mycovirus-Curing in Rhizoctonia spp.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savary, S.; Willocquet, L.; Elazegui, F.A.; Castilla, N.P.; Teng, P.S. Rice pest constraints in tropical Asia: Quantification of yield losses due to rice pests in a range of production situations. Plant Dis. 2000, 84, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Chaijuckam, P.; Davis, R.M. Efficacy of natural plant products on the control of aggregate sheath spot of rice. Plant Dis. 2010, 94, 986–992. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gunnell, P.S.; Webster, R.K. Ceratobasidium oryzae-sativae sp. nov., the teleomorph of rhizoctonia oryzae-sativae and Ceratobasidium setariae comb. nov., the probable teleomorph of Rhizoctonia fumigata comb. nov. Mycologia 1987, 79, 731. [Google Scholar] [CrossRef]
- Lanoiselet, V.L.; Cother, E.J.; Ash, G.J.; Harper, J.D.I. Yield loss in rice caused by Rhizoctonia oryzae and R. oryzae-sativae in Australia. Australas. Plant Pathol. 2005, 34, 175–179. [Google Scholar] [CrossRef]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in biological control: From basic research to field implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal–plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef]
- Chen, B.; Nuss, D.L. Infectious cDNA clone of hypovirus CHV1-Euro7: A comparative virology approach to investigate virus-mediated hypovirulence of the chestnut blight fungus Cryphonectria parasitica. J. Virol. 1999, 73, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Biological control of chestnut blight: An example of virus-mediated attenuation of fungal pathogenesis. Microbiol. Rev. 1992, 56, 561–576. [Google Scholar] [CrossRef]
- Chiba, S.; Salaipeth, L.; Lin, Y.-H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel bipartite double-stranded RNA mycovirus from the white root rot fungus Rosellinia necatrix: Molecular and biological characterization, taxonomic considerations, and potential for biological control. J. Virol. 2009, 83, 12801–12812. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S.; Ghabrial, S.A.; Xie, J. A novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G.; Peng, Y.; Xie, J.; Cheng, J.; Huang, J.; et al. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef] [PubMed]
- Aihara, M.; Urayama, S.; Le, M.T.; Katoh, Y.; Higashiura, T.; Fukuhara, T.; Arie, T.; Teraoka, T.; Komatsu, K.; Moriyama, H. Infection by Magnaporthe oryzae chrysovirus 1 strain A triggers reduced virulence and pathogenic race conversion of its host fungus, Magnaporthe oryzae. J. Gen. Plant Pathol. 2018, 84, 92–103. [Google Scholar] [CrossRef]
- Urayama, S.; Sakoda, H.; Takai, R.; Katoh, Y.; Minh Le, T.; Fukuhara, T.; Arie, T.; Teraoka, T.; Moriyama, H. A dsRNA mycovirus, Magnaporthe oryzae chrysovirus 1-B, suppresses vegetative growth and development of the rice blast fungus. Virology 2014, 448, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Shu, C.; Zhang, M.; Yang, M.; Zhou, E. Molecular characterization of a novel endornavirus conferring hypovirulence in rice sheath blight fungus Rhizoctonia solani AG-1 IA strain GD-2. Viruses 2019, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Abdoulaye, A.H.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. Complete genome sequence of a novel mitovirus from the phytopathogenic fungus Rhizoctonia oryzae-sativae. Arch. Virol. 2017, 162, 1409–1412. [Google Scholar] [CrossRef]
- Das, S.; Das, S. First report of a novel alphapartitivirus in the basidiomycete Rhizoctonia oryzae-sativae. Arch. Virol. 2019, 164, 889–892. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Arjona-López, J.M.; Telengech, P.; Suzuki, N.; López-Herrera, C.J. Coinfection of Rosellinia necatrix by a partitivirus and a virga-like virus is associated with hypovirulence. Eur. J. Plant Pathol. 2020, 158, 111–119. [Google Scholar] [CrossRef]
- Kamaruzzaman, M.; He, G.; Wu, M.; Zhang, J.; Yang, L.; Chen, W.; Li, G. A novel partitivirus in the hypovirulent isolate QT5-19 of the plant pathogenic fungus Botrytis cinerea. Viruses 2019, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, M.; Chen, Q.; Zhu, M.; Zhou, E. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456–457, 220–226. [Google Scholar] [CrossRef]
- Eusebio-Cope, A.; Suzuki, N. Mycoreovirus genome rearrangements associated with RNA silencing deficiency. Nucleic Acids Res. 2015, 43, 3802–3813. [Google Scholar] [CrossRef]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Telengech, P.; Hisano, S.; Mugambi, C.; Hyodo, K.; Arjona-López, J.M.; López-Herrera, C.J.; Kanematsu, S.; Kondo, H.; Suzuki, N. Diverse partitiviruses from the phytopathogenic fungus, Rosellinia necatrix. Front. Microbiol. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Lyu, R.; Zhang, Y.; Tang, Q.; Li, Y.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Xie, J. Two alphapartitiviruses co-infecting a single isolate of the plant pathogenic fungus Rhizoctonia solani. Arch. Virol. 2018, 163, 515–520. [Google Scholar] [CrossRef]
- Picarelli, M.A.S.C.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M.; Colariccio, A. Extreme diversity of mycoviruses present in isolates of Rhizoctonia solani AG2-2 LP from Zoysia japonica from brazil. Front. Cell. Infect. Microbiol. 2019, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zeng, M.; Zhang, M.; Shu, C.; Zhou, E. Complete nucleotide sequence of a partitivirus from Rhizoctonia solani AG-1 IA strain C24. Viruses 2018, 10, 703. [Google Scholar] [CrossRef] [PubMed]
- Abdoulaye, A.H.; Hai, D.; Tang, Q.; Jiang, D.; Fu, Y.; Cheng, J.; Lin, Y.; Li, B.; Kotta-Loizou, I.; Xie, J. Two distant helicases in one mycovirus: Evidence of horizontal gene transfer between mycoviruses, coronaviruses and other nidoviruses. Virus Evol. 2021, 7, 1–11. [Google Scholar] [CrossRef]
- Zhang, M.; Zheng, L.; Liu, C.; Shu, C.; Zhou, E. Characterization of a novel dsRNA mycovirus isolated from strain A105 of Rhizoctonia solani AG-1 IA. Arch. Virol. 2018, 163, 427–430. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, H.; Zhang, M.; Cao, X.; Zhou, E. The complete genomic sequence of a novel mycovirus from Rhizoctonia solani AG-1 IA strain B275. Arch. Virol. 2013, 158, 1609–1612. [Google Scholar] [CrossRef]
- Zhong, J.; Chen, C.-Y.; Gao, B.-D. Genome sequence of a novel mycovirus of Rhizoctonia solani, a plant pathogenic fungus. Virus Genes 2015, 51, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tong Gai, X.; Xing Chen, R.; Li, C.X.; Zhao, G.K.; Yuan Xia, Z.; Zou, C.M.; Zhong, J. Characterization of three novel betapartitiviruses co-infecting the phytopathogenic fungus Rhizoctonia solani. Virus Res. 2019, 270, 197649. [Google Scholar] [CrossRef]
- Ihrmark, K.; Stenström, E.; Stenlid, J. Double-stranded RNA transmission through basidiospores of Heterobasidion annosum. Mycol. Res. 2004, 108, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Kanematsu, S.; Sasaki, A.; Onoue, M.; Oikawa, Y.; Ito, T. Extending the fungal host range of a partitivirus and a mycoreovirus from Rosellinia necatrix by inoculation of protoplasts with virus particles. Phytopathology 2010, 100, 922–930. [Google Scholar] [CrossRef]
- Cubeta, M.A.; Vilgalys, R. Population biology of the Rhizoctonia solani complex. Phytopathology 1997, 87, 446–447. [Google Scholar] [CrossRef] [PubMed]
- Ogoshi, A. Ecology and pathogenicity of anastomosis and interspecific groups of Rhizoctonia solani Kühn. Annu. Rev. Phytopathol. 1987, 25, 124–143. [Google Scholar] [CrossRef]
- Taheri, P.; Gnanamanickam, S.; Höfte, M. Characterization, genetic structure, and pathogenicity of Rhizoctonia spp. associated with rice sheath diseases in India. Phytopathology 2007, 97, 373–383. [Google Scholar] [CrossRef]
- Chiba, S.; Lin, Y.-H.; Kondo, H.; Kanematsu, S.; Suzuki, N. A novel victorivirus from a phytopathogenic fungus, Rosellinia necatrix, is infectious as particles and targeted by RNA silencing. J. Virol. 2013, 87, 6727–6738. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Lin, Y.-H.; Kondo, H.; Kanematsu, S.; Suzuki, N. Effects of defective interfering RNA on symptom induction by, and replication of, a novel partitivirus from a phytopathogenic fungus, Rosellinia necatrix. J. Virol. 2013, 87, 2330–2341. [Google Scholar] [CrossRef]
- Chiba, S.; Lin, Y.; Kondo, H.; Kanematsu, S.; Suzuki, N. A novel betapartitivirus RnPV6 from Rosellinia necatrix tolerates host RNA silencing but is interfered by its defective RNAs. Virus Res. 2016, 219, 62–72. [Google Scholar] [CrossRef]
- Vainio, E.J.; Hakanpää, J.; Dai, Y.-C.; Hansen, E.; Korhonen, K.; Hantula, J. Species of Heterobasidion host a diverse pool of partitiviruses with global distribution and interspecies transmission. Fungal Biol. 2011, 115, 1234–1243. [Google Scholar] [CrossRef]
- Cubeta, M.A. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395. [Google Scholar] [CrossRef]
- Zhong, J.; Chen, D.; Zhu, H.J.; Gao, B.D.; Zhou, Q. Hypovirulence of Sclerotium rolfsii caused by associated RNA mycovirus. Front. Microbiol. 2016, 7, 1–18. [Google Scholar] [CrossRef]
- Kondo, H.; Kanematsu, S.; Suzuki, N. Viruses of the White Root Rot Fungus, Rosellinia necatrix. Adv. Virus Res. 2013, 86, 177–214. [Google Scholar] [CrossRef] [PubMed]
- Hillman, B.I.; Annisa, A.; Suzuki, N. Viruses of Plant-Interacting Fungi. Adv. Virus Res. 2018, 100, 99–116. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, T.; Luo, C.; Jiang, D.; Li, G.; Li, Q.; Hsiang, T.; Huang, J. Prevalence and diversity of mycoviruses infecting the plant pathogen Ustilaginoidea virens. Virus Res. 2015, 195, 47–56. [Google Scholar] [CrossRef]
- Parker, W.B. Metabolism and antiviral activity of ribavirin. Virus Res. 2005, 107, 165–171. [Google Scholar] [CrossRef]
- Schneider-Poetsch, T.; Ju, J.; Eyler, D.E.; Dang, Y.; Bhat, S.; Merrick, W.C.; Green, R.; Shen, B.; Liu, J.O. Inhibition of eukaryotic translation elongation by cycloheximide and lactimidomycin. Nat. Chem. Biol. 2010, 6, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Yang, H.-E.; Kim, D.-H. Identification of a novel partitivirus of Trichoderma harzianum NFCF319 and evidence for the related antifungal activity. Front. Plant Sci. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Sasaki, A.; Nakamura, H.; Suzuki, N.; Kanematsu, S. Characterization of a new megabirnavirus that confers hypovirulence with the aid of a co-infecting partitivirus to the host fungus, Rosellinia necatrix. Virus Res. 2016, 219, 73–82. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungus Species | Number of Isolates | dsRNA (%) * | Fungus Species | Number of Isolates | dsRNA (%) * |
|---|---|---|---|---|---|
| Achroiostachys saccharicxola | 3 | 0 (0%) | Nigrospora sphaerica | 2 | 1 (50%) |
| Bipolaris oryzae | 1 | 0 (0%) | Sarocladium oryzae | 1 | 0 (0%) |
| Bipolaris sivanesaniana | 7 | 0 (0%) | Sclerotium hydrophilum | 2 | 0 (0%) |
| Curvularia beasleyi | 1 | 0 (0%) | Setosphaeria rostrata | 2 | 0 (0%) |
| Fusarium equiseti | 1 | 0 (0%) | Simplicillium lamellicola | 1 | 0 (0%) |
| Fusarium incarnatum | 1 | 0 (0%) | Stachybotrys cf. elegans | 2 | 0 (0%) |
| Fusarium proliferatum | 4 | 0 (0%) | Rhizoctonia oryzae | 3 | 1 (33%) |
| Fusarium sacchari | 1 | 0 (0%) | Rhizoctonia oryzae-sativae | 37 | 12 (36%) |
| Gaeumannomyces oryzinus | 5 | 0 (0%) | Rhizoctonia solani | 1 | 1 (100%) |
| Nigrospora oryzae | 4 | 1 (25%) | Rhizoctonia zeae | 1 | 1 (100%) |
| Rhizoctonia oryzae-sativae Isolates | The aa Sequence Identities to Reported Partitiviruses (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RosPV1-T442 + | RosPV2-T442 + | RosPV3-T505 + | RosPV4-T123 + | RosPV5-T123 + | RosPV6-T123 + | RsRV2 * | RsRV3 * | RsRV4 * | RsRV5 * | RnPV6 * | RnPV16 * | ||
| TSD190103 | RdRp | 99% | 99% | ||||||||||
| CP | |||||||||||||
| TSD190108 | RdRp | 99% | 91% | 86% | |||||||||
| CP | 84% | ||||||||||||
| TSD190123 | RdRp | 100% | 100% | 100% | |||||||||
| CP | 100% | 100% | 100% | ||||||||||
| TSS190401 | RdRp | 91% | 99% | 92% | |||||||||
| CP | 83% | ||||||||||||
| TSS190442 | RdRp | 100% | 100% | 92% | |||||||||
| CP | 100% | 100% | 88% | 86% | |||||||||
| TSS190505 | RdRp | 94% | 100% | ||||||||||
| CP | 100% | 15% | |||||||||||
| Mycovirus | Rhizoctonia spp. | Virus-Curing Attempt | Method | Result of Virus-Curing | Source | |
|---|---|---|---|---|---|---|
| Species | Strain | |||||
| RsRV3 | R. solani | A105 | − | N/A | N/A | [28] |
| RsPV5 | R. solani | C24 | − | N/A | N/A | [26] |
| RsRV1 | R. solani | B275 | − | N/A | N/A | [29] |
| RsRV-HN008 | R. solani | HN008 | − | N/A | N/A | [30] |
| RosPV1 | R. oryzae-sativae | RS005 | − | N/A | N/A | [16] |
| RoMV1 | R. oryzae-sativae | 89-1 | − | N/A | N/A | [15] |
| RsEV1 | R. solani | GD-2 | + | Hyphal tipping | Unsuccessful | [14] |
| RsPV2 | R. solani | GD-11 | + | Hyphal tipping | Unsuccessful | [20] |
| RsPV6, 7, 8 | R. solani | YNBB-111 | + | Hyphal tipping and ribavirin | Unsuccessful | [31] |
| Protoplasting | Unsuccessful | |||||
| RsHV2 | R. solani | XN84 | + | Protoplasting | Successful | [27] |
| RosPV1 & 2-T442 | R. oryzae-sativae | TSD190442 | + | Protoplasting | Unsuccessful | |
| Heat treatment | Unsuccessful | |||||
| Hyphal tipping | Unsuccessful | |||||
| Hyphal tipping and ribavirin | Unsuccessful | This study | ||||
| Hyphal tipping and ribavirin and cycloheximide | Successful | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neang, S.; Bincader, S.; Rangsuwan, S.; Keawmanee, P.; Rin, S.; Salaipeth, L.; Das, S.; Kondo, H.; Suzuki, N.; Sato, I.; et al. Omnipresence of Partitiviruses in Rice Aggregate Sheath Spot Symptom-Associated Fungal Isolates from Paddies in Thailand. Viruses 2021, 13, 2269. https://doi.org/10.3390/v13112269
Neang S, Bincader S, Rangsuwan S, Keawmanee P, Rin S, Salaipeth L, Das S, Kondo H, Suzuki N, Sato I, et al. Omnipresence of Partitiviruses in Rice Aggregate Sheath Spot Symptom-Associated Fungal Isolates from Paddies in Thailand. Viruses. 2021; 13(11):2269. https://doi.org/10.3390/v13112269
Chicago/Turabian StyleNeang, Sokty, Santiti Bincader, Sansern Rangsuwan, Pisut Keawmanee, Soriya Rin, Lakha Salaipeth, Subha Das, Hideki Kondo, Nobuhiro Suzuki, Ikuo Sato, and et al. 2021. "Omnipresence of Partitiviruses in Rice Aggregate Sheath Spot Symptom-Associated Fungal Isolates from Paddies in Thailand" Viruses 13, no. 11: 2269. https://doi.org/10.3390/v13112269
APA StyleNeang, S., Bincader, S., Rangsuwan, S., Keawmanee, P., Rin, S., Salaipeth, L., Das, S., Kondo, H., Suzuki, N., Sato, I., Takemoto, D., Rattanakreetakul, C., Pongpisutta, R., Arakawa, M., & Chiba, S. (2021). Omnipresence of Partitiviruses in Rice Aggregate Sheath Spot Symptom-Associated Fungal Isolates from Paddies in Thailand. Viruses, 13(11), 2269. https://doi.org/10.3390/v13112269