
Lasting Changes to Circulating Leukocytes in People with Mild SARS-CoV-2 Infections

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Recruitment and Blood Collection
2.2. Measurements of Anti-SARS-CoV-2 Antibodies
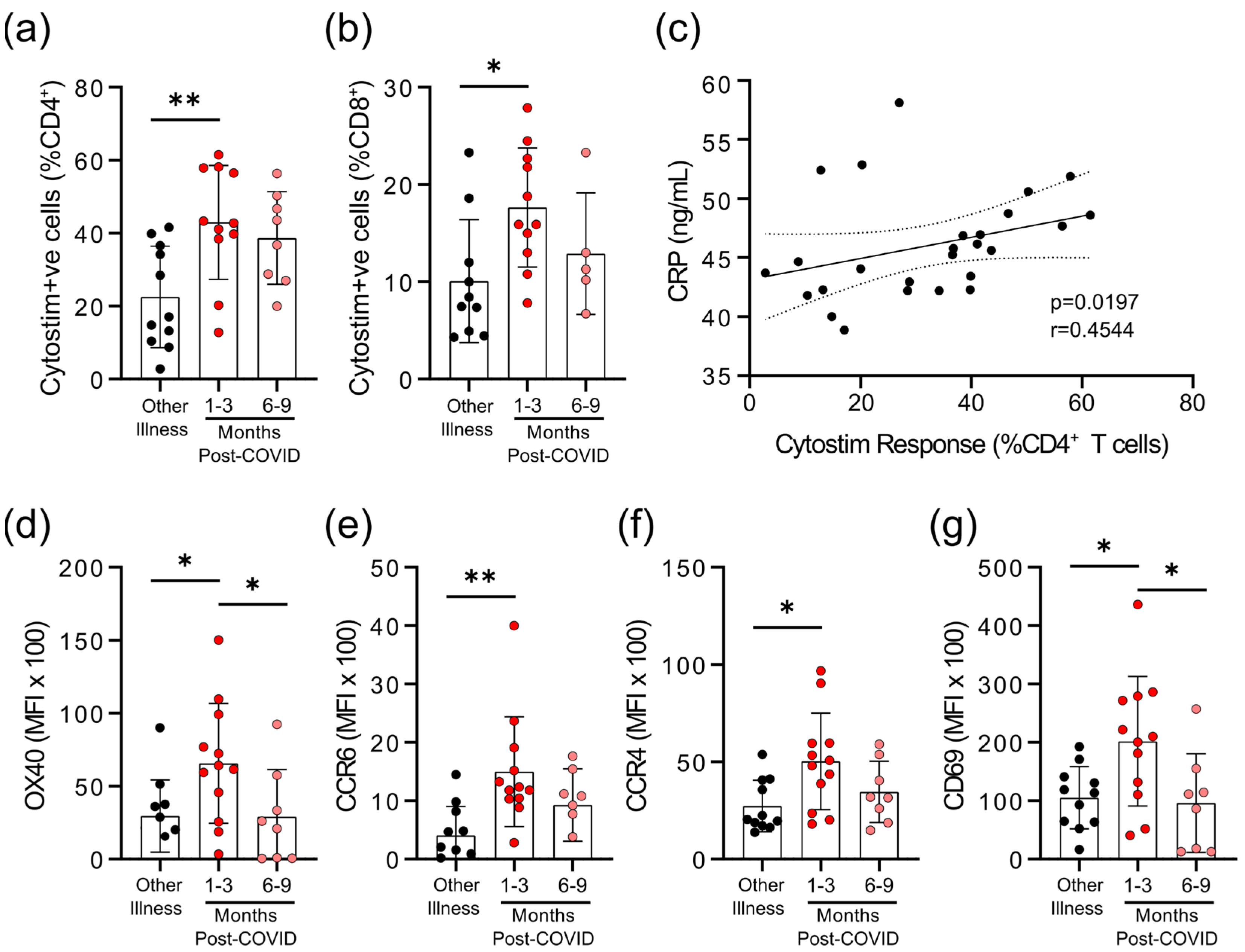
2.3. Assessment of T Cell Activation Induced Markers (AIM) for SARS-CoV-2 Peptides
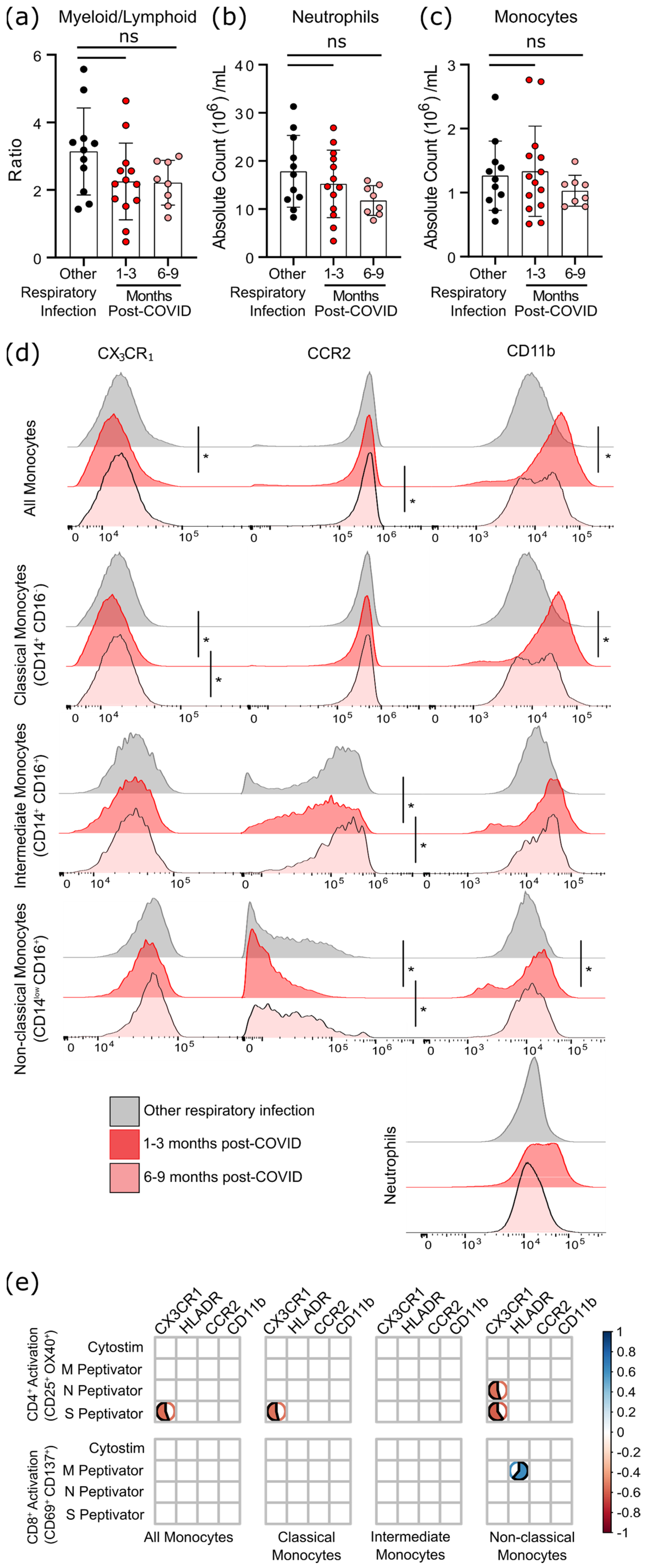
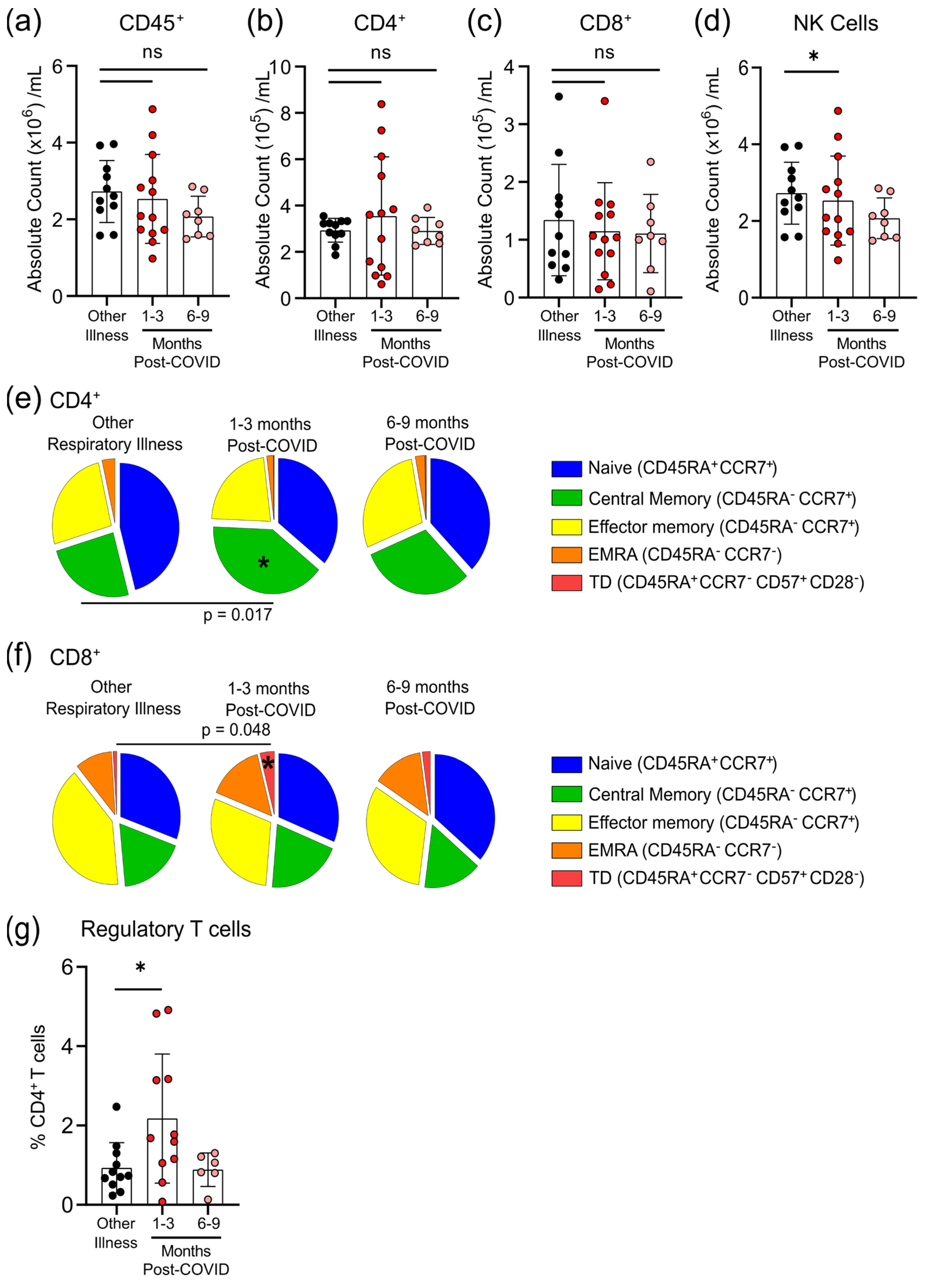
2.4. Quantification of Peripheral Immunophenotype
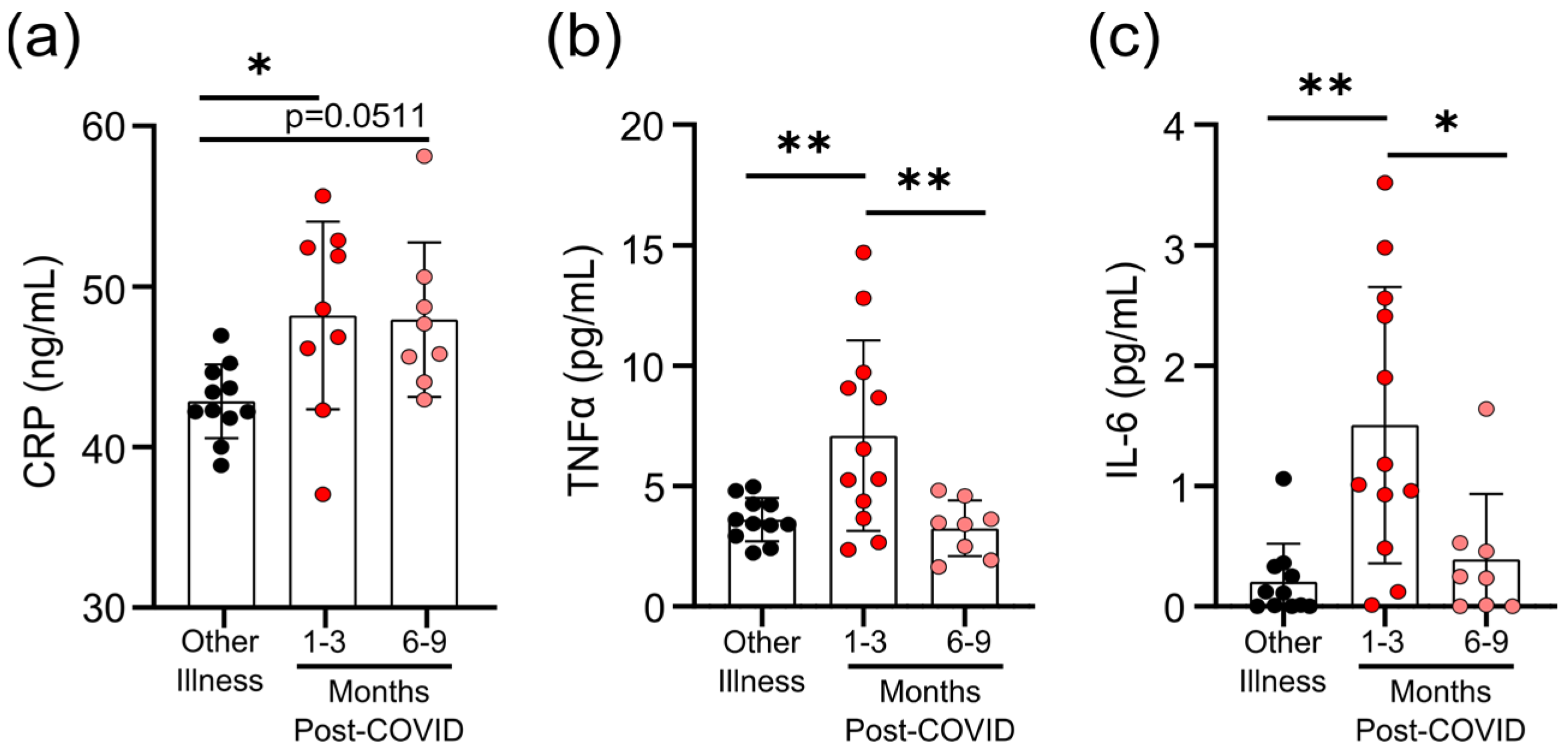
2.5. Measurements of Cytokines and C-Reactive Protein
2.6. Statistical Analysis
3. Results
3.1. Demographics and Symptoms of Individuals Who Had SARS-CoV-2 and Other Respiratory Infections
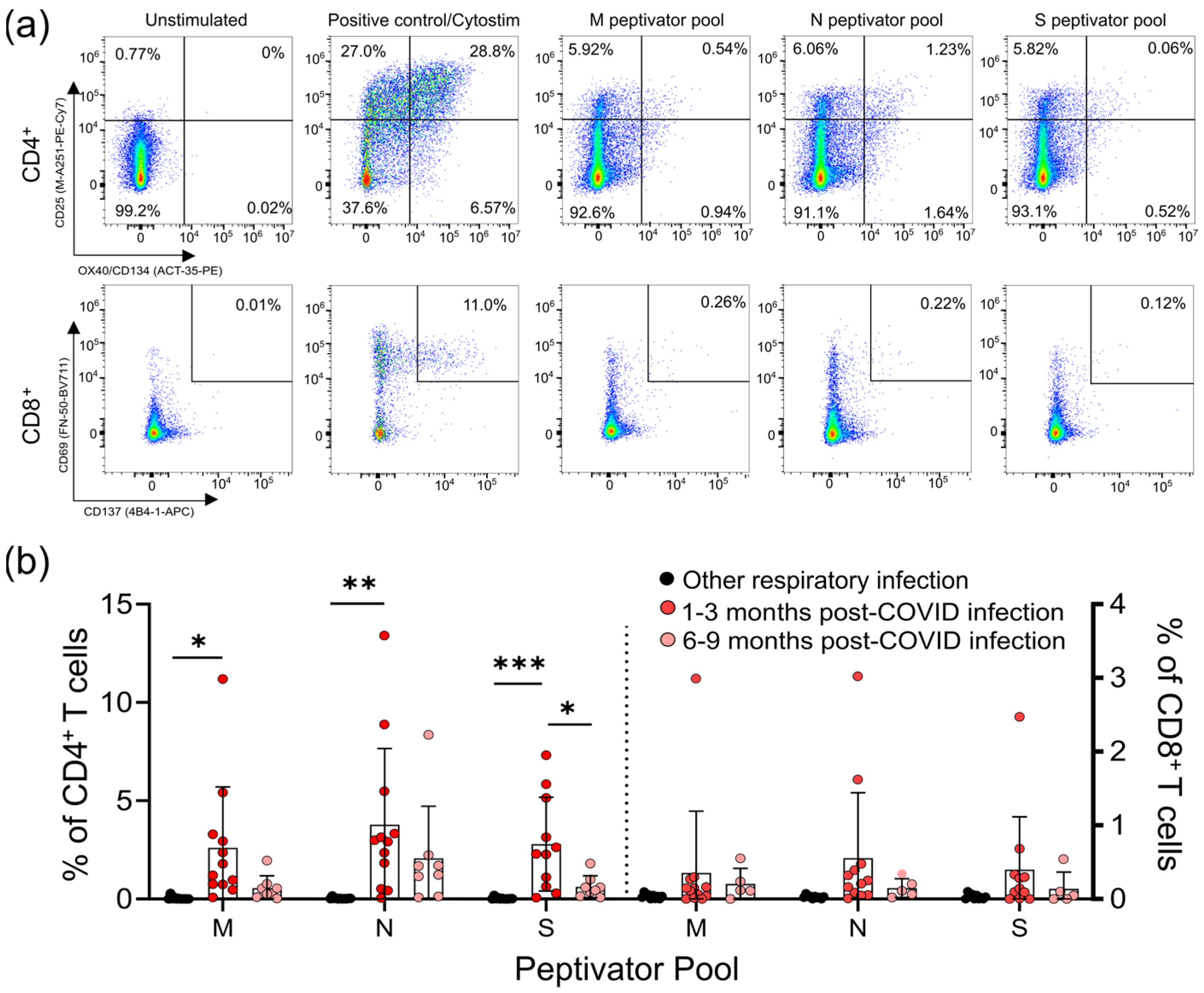
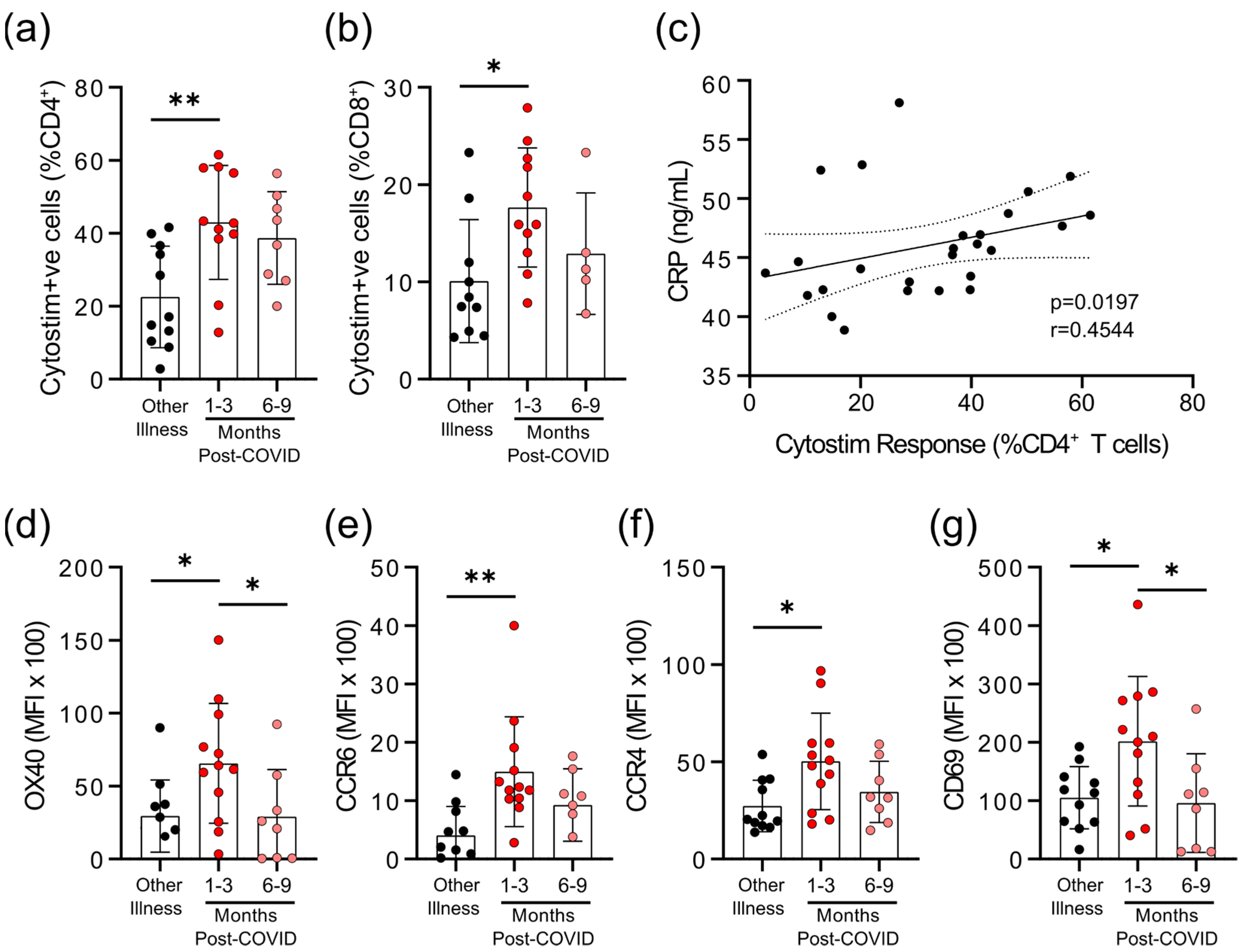
3.2. Survivors of Mild SARS-CoV-2 Infections Have Reduced T Cell Responses to SARS-CoV-2 Peptides 6–9 Months Post Symptom Onset
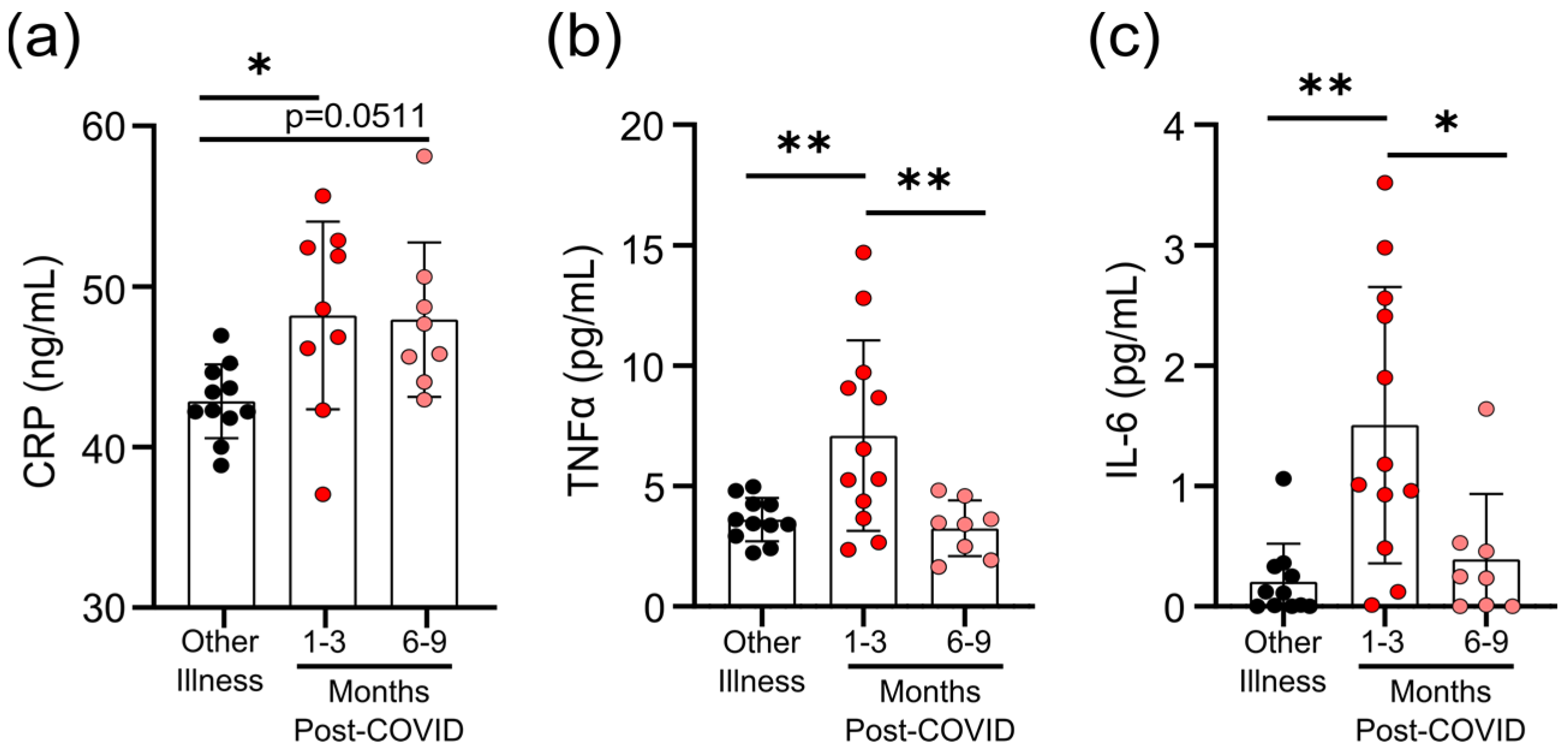
3.3. Survivors of Mild SARS-CoV-2 Infections Have Evidence of Sustained Inflammation 1–3 Months Post Symptom Onset
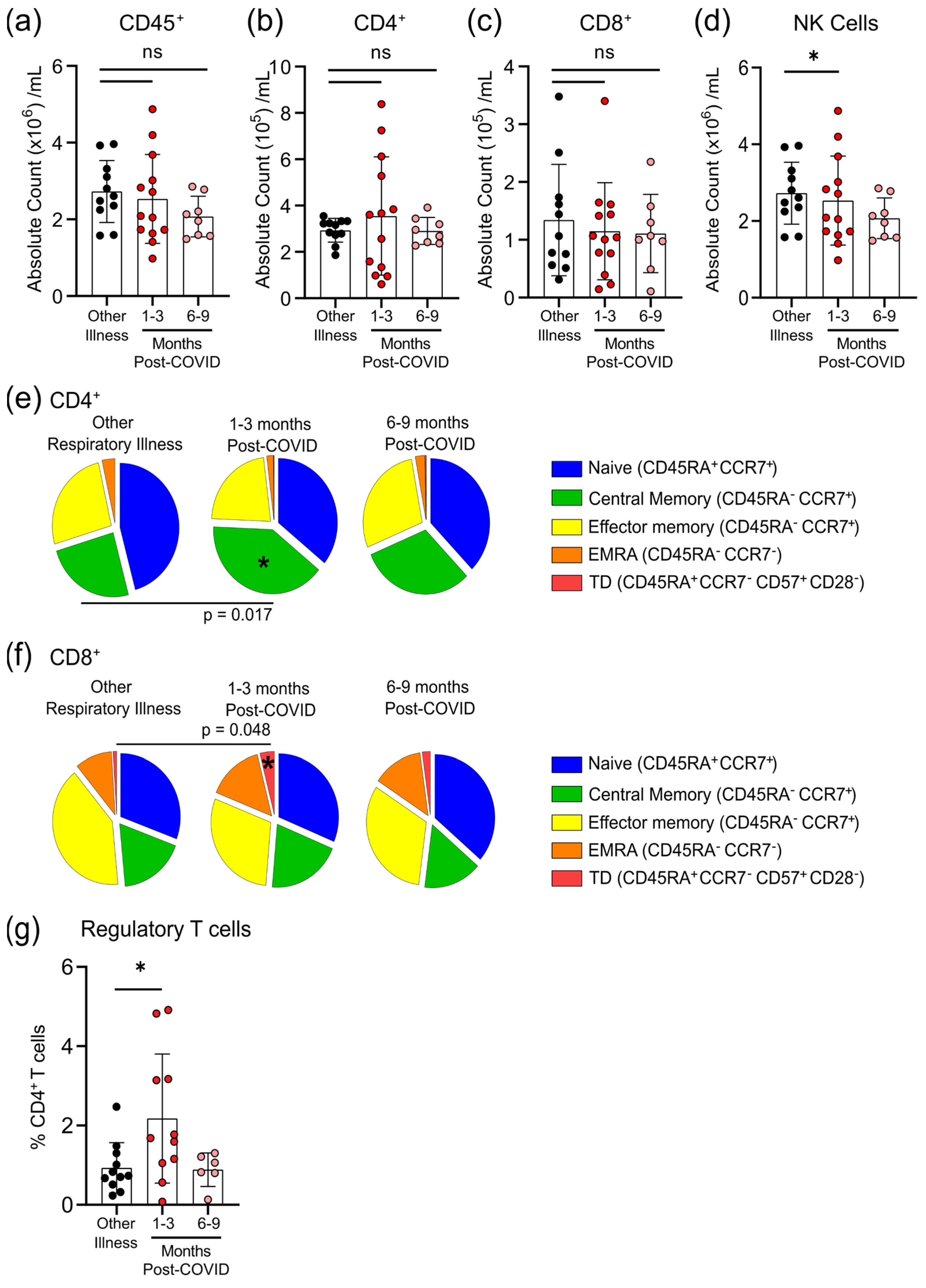
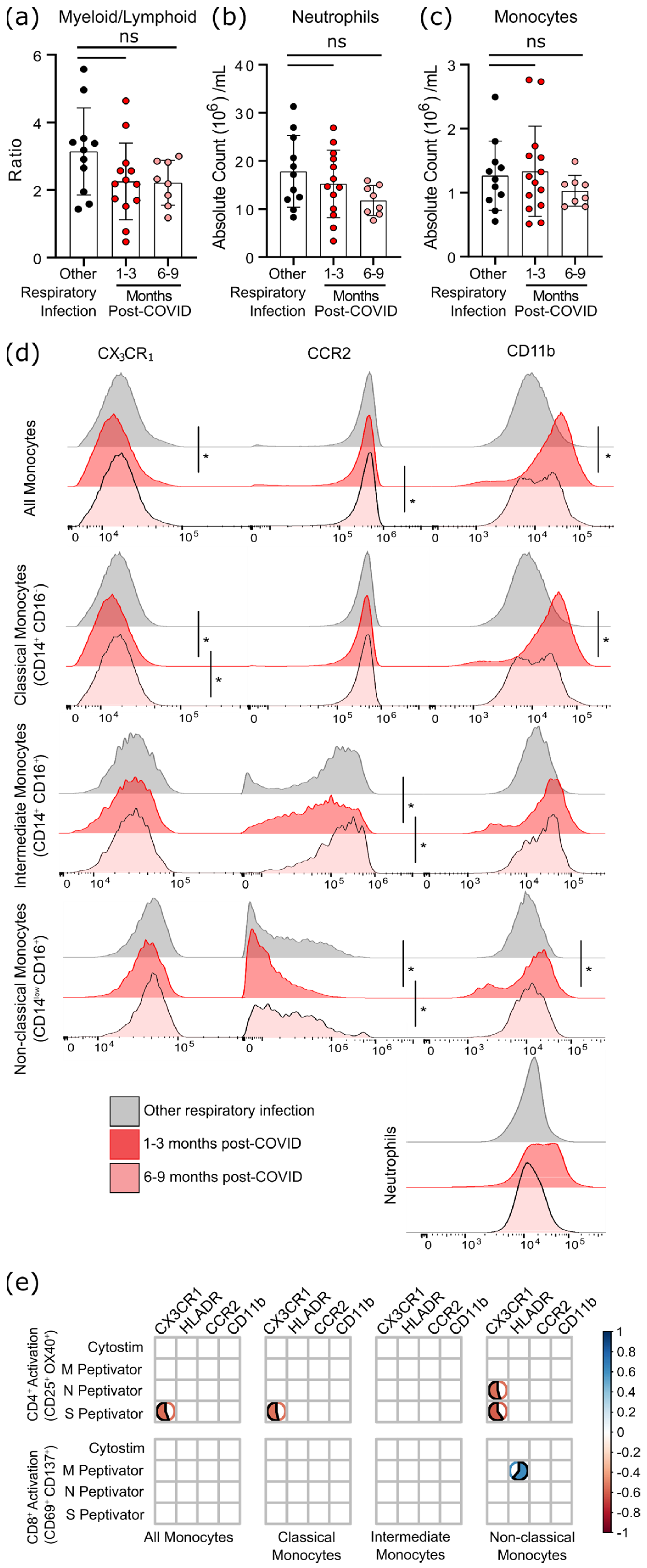
3.4. Survivors of Mild SARS-CoV-2 Infections Have Changes in Circulating Immunophenotype 1–3 Months Post-Symptom Onset
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alon, R.; Sportiello, M.; Kozlovski, S.; Kumar, A.; Reilly, E.C.; Zarbock, A.; Garbi, N.; Topham, D.J. Leukocyte trafficking to the lungs and beyond: Lessons from influenza for COVID-19. Nat. Rev. Immunol. 2021, 21, 49–64. [Google Scholar] [CrossRef]
- Huang, I.; Pranata, R. Lymphopenia in severe coronavirus disease-2019 (COVID-19): Systematic review and meta-analysis. J. Intensive Care 2020, 8, 36. [Google Scholar] [CrossRef]
- Chen, Z.; John Wherry, E. T cell responses in patients with COVID-19. Nat. Rev. Immunol. 2020, 20, 529–536. [Google Scholar] [CrossRef]
- De Candia, P.; Prattichizzo, F.; Garavelli, S.; Matarese, G. T Cells: Warriors of SARS-CoV-2 Infection. Trends Immunol. 2021, 42, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Guo, R.; Lei, L.; Liu, H.; Wang, Y.; Wang, Y.; Qian, H.; Dai, T.; Zhang, T.; Lai, Y. COVID-19 infection induces readily detectable morphologic and inflammation-related phenotypic changes in peripheral blood monocytes. J. Leukoc. Biol. 2021, 109, 13–22. [Google Scholar] [CrossRef]
- Prescott, H.C.; Girard, T.D. Recovery From Severe COVID-19: Leveraging the Lessons of Survival From Sepsis. JAMA 2020, 324, 739–740. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, T.; Knight, M.; A’Court, C.; Buxton, M.; Husain, L. Management of post-acute COVID-19 in primary care. BMJ 2020, 370, m3026. [Google Scholar] [CrossRef] [PubMed]
- Ayoubkhani, D.; Khunti, K.; Nafilyan, V.; Maddox, T.; Humberstone, B.; Diamond, I.; Banerjee, A. Post-COVID syndrome in individuals admitted to hospital with COVID-19: Retrospective cohort study. BMJ 2021, 372, n693. [Google Scholar] [CrossRef] [PubMed]
- Chopra, V.; Flanders, S.A.; O’Malley, M.; Malani, A.N.; Prescott, H.C. Sixty-Day Outcomes Among Patients Hospitalized With COVID-19. Ann. Intern. Med. 2020, 174, 576–578. [Google Scholar] [CrossRef]
- Donnelly, J.P.; Wang, X.Q.; Iwashyna, T.J.; Prescott, H.C. Readmission and Death after Initial Hospital Discharge among Patients with COVID-19 in a Large Multihospital System. JAMA 2020, 325, 304–306. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Stralin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verschoor, C.P.; Kohli, V.; Balion, C. A comprehensive assessment of immunophenotyping performed in cryopreserved peripheral whole blood. Cytom. B Clin. Cytom. 2018, 94, 662–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, A.; Arnold, D.M.; Smith, J.W.; Moore, J.C.; Zhang, A.; Chagla, Z.; Harvey, B.J.; Stacey, H.D.; Ang, J.C.; Clare, R.; et al. Characteristics of Anti-SARS-CoV-2 Antibodies in Recovered COVID-19 Subjects. Viruses 2021, 13, 697. [Google Scholar] [CrossRef]
- Zaunders, J.J.; Munier, M.L.; Seddiki, N.; Pett, S.; Ip, S.; Bailey, M.; Xu, Y.; Brown, K.; Dyer, W.B.; Kim, M.; et al. High Levels of Human Antigen-Specific CD4+ T Cells in Peripheral Blood Revealed by Stimulated Coexpression of CD25 and CD134 (OX40). J. Immunol. 2009, 183, 2827–2836. [Google Scholar] [CrossRef] [Green Version]
- Sadler, R.; Bateman, E.A.; Heath, V.; Patel, S.Y.; Schwingshackl, P.P.; Cullinane, A.C.; Ayers, L.; Ferry, B.L. Establishment of a healthy human range for the whole blood “OX40” assay for the detection of antigen-specific CD4+ T cells by flow cytometry. Cytom. B Clin. Cytom. 2014, 86, 350–361. [Google Scholar] [CrossRef]
- Seddiki, N.; Cook, L.; Hsu, D.C.; Phetsouphanh, C.; Brown, K.; Xu, Y.; Kerr, S.J.; Cooper, D.A.; Munier, C.M.; Pett, S.; et al. Human antigen-specific CD4(+) CD25(+) CD134(+) CD39(+) T cells are enriched for regulatory T cells and comprise a substantial proportion of recall responses. Eur. J. Immunol. 2014, 44, 1644–1661. [Google Scholar] [CrossRef]
- Wolfl, M.; Kuball, J.; Ho, W.Y.; Nguyen, H.; Manley, T.J.; Bleakley, M.; Greenberg, P.D. Activation-induced expression of CD137 permits detection, isolation, and expansion of the full repertoire of CD8+ T cells responding to antigen without requiring knowledge of epitope specificities. Blood 2007, 110, 201–210. [Google Scholar] [CrossRef]
- Loukov, D.; Karampatos, S.; Maly, M.R.; Bowdish, D.M.E. Monocyte activation is elevated in women with knee-osteoarthritis and associated with inflammation, BMI and pain. Osteoarthr. Cartil. 2018, 26, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Ontario Agency for Health Protection and Promotion (Public Health Ontario). COVID-19 Serosurveillance Summary—Seroprevalence in Ontario: March 27, 2020 to June 30, 2020. Available online: https://www.publichealthontario.ca/-/media/documents/ncov/epi/2020/07/COVID-19-epi-seroprevalence-in-ontario.pdf?la=en (accessed on 31 October 2021).
- Government of Canada. Respiratory Virus Report, Week 34—Ending 22 August 2020. Available online: https://www.canada.ca/en/public-health/services/surveillance/respiratory-virus-detections-canada/2019-2020/week-34-ending-august-22-2020.html (accessed on 31 October 2021).
- Honigsbaum, M.; Krishnan, L. Taking pandemic sequelae seriously: From the Russian influenza to COVID-19 long-haulers. Lancet 2020, 396, 1389–1391. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: What do we know about “long COVID”? BMJ 2020, 370, m2815. [Google Scholar] [CrossRef]
- Kratofil, R.M.; Kubes, P.; Deniset, J.F. Monocyte Conversion during Inflammation and Injury. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 35–42. [Google Scholar] [CrossRef] [Green Version]
- El Khoury, J.; Toft, M.; Hickman, S.E.; Means, T.K.; Terada, K.; Geula, C.; Luster, A.D. Ccr2 deficiency impairs microglial accumulation and accelerates progression of Alzheimer-like disease. Nat. Med. 2007, 13, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Auffray, C.; Fogg, D.; Garfa, M.; Elain, G.; Join-Lambert, O.; Kayal, S.; Sarnacki, S.; Cumano, A.; Lauvau, G.; Geissmann, F. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 2007, 317, 666–670. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.; Tacke, R.; Hedrick, C.C.; Hanna, R.N. Nonclassical Patrolling Monocyte Function in the Vasculature. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1306–1316. [Google Scholar] [CrossRef] [Green Version]
- Al-Aly, Z.; Xie, Y.; Bowe, B. High-dimensional characterization of post-acute sequalae of COVID-19. Nature 2021, 594, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.M.; Zhu, J.; Huang, X.; Yang, Y. The Development and Function of Memory Regulatory T Cells after Acute Viral Infections. J. Immunol. 2012, 189, 2805–2814. [Google Scholar] [CrossRef] [PubMed]
- Betts, R.J.; Prabhu, N.; Ho, A.W.S.; Lew, F.C.; Hutchinson, P.E.; Rotzschke, O.; Macary, P.A.; Kemeny, D.M. Influenza A Virus Infection Results in a Robust, Antigen-Responsive, and Widely Disseminated Foxp3+ Regulatory T Cell Response. J. Virol. 2012, 86, 2817–2825. [Google Scholar] [CrossRef] [Green Version]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef]
- Verdoni, L.; Mazza, A.; Gervasoni, A.; Martelli, L.; Ruggeri, M.; Ciuffreda, M.; Bonanomi, E.; D’Antiga, L. An outbreak of severe Kawasaki-like disease at the Italian epicentre of the SARS-CoV-2 epidemic: An observational cohort study. Lancet 2020, 395, 1771–1778. [Google Scholar] [CrossRef]
- Ramos-Casals, M.; Brito-Zerón, P.; Mariette, X. Systemic and organ-specific immune-related manifestations of COVID-19. Nat. Rev. Rheumatol. 2021, 17, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Lanteri, M.C.; O’Brien, K.M.; Purtha, W.E.; Cameron, M.J.; Lund, J.M.; Owen, R.E.; Heitman, J.W.; Custer, B.; Hirschkorn, D.F.; Tobler, L.H.; et al. Tregs control the development of symptomatic West Nile virus infection in humans and mice. J. Clin. Investig. 2009, 119, 3266–3277. [Google Scholar] [CrossRef] [Green Version]
- Stukalov, A.; Girault, V.; Grass, V.; Karayel, O.; Bergant, V.; Urban, C.; Haas, D.A.; Huang, Y.; Oubraham, L.; Wang, A.; et al. Multilevel proteomics reveals host perturbations by SARS-CoV-2 and SARS-CoV. Nature 2021, 594, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cui, Y.; Feng, H.; Liu, H.; Liu, X. TGF-β signaling controls Foxp3 methylation and T reg cell differentiation by modulating Uhrf1 activity. J. Exp. Med. 2019, 216, 2819–2837. [Google Scholar] [CrossRef] [PubMed]
- Lüke, F.; Orsó, E.; Kirsten, J.; Poeck, H.; Grube, M.; Wolff, D.; Burkhardt, R.; Lunz, D.; Lubnow, M.; Schmidt, B.; et al. Coronavirus disease 2019 induces multi-lineage, morphologic changes in peripheral blood cells. EJHaem 2020. [Google Scholar] [CrossRef]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419–1440.e23. [Google Scholar] [CrossRef]
- Pérez-Gómez, A.; Vitallé, J.; Gasca-Capote, C.; Gutierrez-Valencia, A.; Trujillo-Rodriguez, M.; Serna-Gallego, A.; Muñoz-Muela, E.; Jimenez-Leon, M.R.; Benhnia, M.R.; Rivas-Jeremias, I.; et al. Dendritic cell deficiencies persist seven months after SARS-CoV-2 infection. Cell Mol. Immunol. 2021, 18, 2128–2139. [Google Scholar] [CrossRef]
- McKechnie, J.L.; Blish, C.A. The Innate Immune System: Fighting on the Front Lines or Fanning the Flames of COVID-19? Cell Host Microbe 2020, 27, 863–869. [Google Scholar] [CrossRef]
- Cabaro, S.; D’Esposito, V.; Di Matola, T.; Sale, S.; Cennamo, M.; Terracciano, D.; Parisi, V.; Oriente, F.; Portella, G.; Beguinot, F.; et al. Cytokine signature and COVID-19 prediction models in the two waves of pandemics. Sci. Rep. 2021, 11, 20793. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Bainton, D.F.; Borregaard, N.; Springer, T.A. Stimulated mobilization of monocyte Mac-1 and p150,95 adhesion proteins from an intracellular vesicular compartment to the cell surface. J. Clin. Investig. 1987, 80, 535–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Other Respiratory Infections (Not COVID-19) n = 11 | COVID-19 Infections n = 22 | Indeterminate n = 5 | p Value 1 | |
|---|---|---|---|---|
| Age (mean ± STDEV) | 55 ± 16 | 55 ± 15 | 57 ± 11 | 0.9249 |
| Sex (% female) | 8 (73%) | 11 (50%) | 3 (60%) | 0.2783 |
| BMI (kg/m2) | 24.0 ± 2.9 | 25.0 ± 3.6 | 24.5 ± 2.3 | 0.4172 |
| Health Conditions (frequency) | ||||
| Asthma | 1 (9%) | 1 (5%) | 1 (20%) | - |
| COPD (including emphysema and chronic bronchitis) | 1 (9%) | 0 (0%) | 0 (0%) | - |
| Other lung disease | 0 (0%) | 1 (5%) | 0 (0%) | - |
| Diabetes | 0 (0%) | 3 (14%) | 0 (0%) | - |
| Hypertension | 1 (9%) | 1 (5%) | 1 (20%) | - |
| Heart disease | 0 (0%) | 2 (9%) | 0 (0%) | - |
| Cancer | 2 (18%) | 2 (9%) | 0 (0%) | - |
| Autoimmune condition | 1 (9%) | 1 (5%) | 1 (20%) | - |
| Medications | ||||
| Number of medications (mean ± STDEV) | 1 ± 2 | 2 ± 2 | 1 ± 1 | 0.2049 |
| Symptoms (frequency) | ||||
| Cough | 6 (55%) | 13 (59%) | 2 (40%) | - |
| Shortness of breath | 4 (36%) | 9 (41%) | 1 (20%) | - |
| Chest pain | 3 (27%) | 9 (41%) | 2 (40%) | - |
| Fever | 3 (27%) | 12 (55%) | 1 (20%) | - |
| Feeling generally unwell | 9 (82%) | 21 (95%) | 2 (40%) | - |
| Abnormally tired | 7 (64%) | 20 (91%) | 2 (40%) | - |
| New confusion | 4 (36%) | 1 (5%) | 1 (20%) | - |
| New generalized muscle aches and pains | 5 (45%) | 16 (73%) | 2 (40%) | - |
| New joint pain | 3 (27%) | 5 (23%) | 2 (40%) | - |
| Earache/infection | 0 (0%) | 1 (5%) | 0 (0%) | - |
| Headache | 6 (55%) | 14 (64%) | 1 (20%) | - |
| Runny/stuffy nose | 7 (64%) | 7 (32%) | 2 (40%) | - |
| Sinus pain | 2 (18%) | 4 (18%) | 1 (20%) | - |
| Sore/scratchy throat | 7 (64%) | 10 (45%) | 2 (40%) | - |
| Loss of appetite | 4 (36%) | 8 (36%) | 2 (40%) | - |
| Loss of taste/smell | 2 (18%) | 11 (50%) | 1 (20%) | - |
| Duration of Symptoms | ||||
| No symptoms—1 week | 4 (36%) | 3 (14%) | 1 (20%) | - |
| 2–4 weeks | 3 (27%) | 11 (50%) | 4 (80%) | - |
| 4 or more weeks | 4 (36%) | 8 (36%) | 0 | - |
| Diagnosis | ||||
| PCR test for SARS-CoV-2 performed | 0 (0%) | 15 (68%) | 0 (0%) | - |
| Told by a health care professional that they had SARS-CoV-2/COVID-19 without a PCR test | 1 (8%) | 5 (25%) | 0 (0%) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kennedy, A.E.; Cook, L.; Breznik, J.A.; Cowbrough, B.; Wallace, J.G.; Huynh, A.; Smith, J.W.; Son, K.; Stacey, H.; Ang, J.; et al. Lasting Changes to Circulating Leukocytes in People with Mild SARS-CoV-2 Infections. Viruses 2021, 13, 2239. https://doi.org/10.3390/v13112239
Kennedy AE, Cook L, Breznik JA, Cowbrough B, Wallace JG, Huynh A, Smith JW, Son K, Stacey H, Ang J, et al. Lasting Changes to Circulating Leukocytes in People with Mild SARS-CoV-2 Infections. Viruses. 2021; 13(11):2239. https://doi.org/10.3390/v13112239
Chicago/Turabian StyleKennedy, Allison E., Laura Cook, Jessica A. Breznik, Braeden Cowbrough, Jessica G. Wallace, Angela Huynh, James W. Smith, Kiho Son, Hannah Stacey, Jann Ang, and et al. 2021. "Lasting Changes to Circulating Leukocytes in People with Mild SARS-CoV-2 Infections" Viruses 13, no. 11: 2239. https://doi.org/10.3390/v13112239
APA StyleKennedy, A. E., Cook, L., Breznik, J. A., Cowbrough, B., Wallace, J. G., Huynh, A., Smith, J. W., Son, K., Stacey, H., Ang, J., McGeer, A., Coleman, B. L., Larché, M., Larché, M., Hambly, N., Nair, P., Ask, K., Miller, M. S., Bramson, J., ... Bowdish, D. M. E. (2021). Lasting Changes to Circulating Leukocytes in People with Mild SARS-CoV-2 Infections. Viruses, 13(11), 2239. https://doi.org/10.3390/v13112239