
Viruses of the Fall Armyworm Spodoptera frugiperda: A Review with Prospects for Biological Control



Abstract
1. Introduction
2. Current Status of Viruses in the Control of FAW
2.1. Ascoviruses
2.2. Baculoviruses
2.3. Rhabdoviruses
2.4. Other Virus Families
2.5. NGS to Discover New Viruses
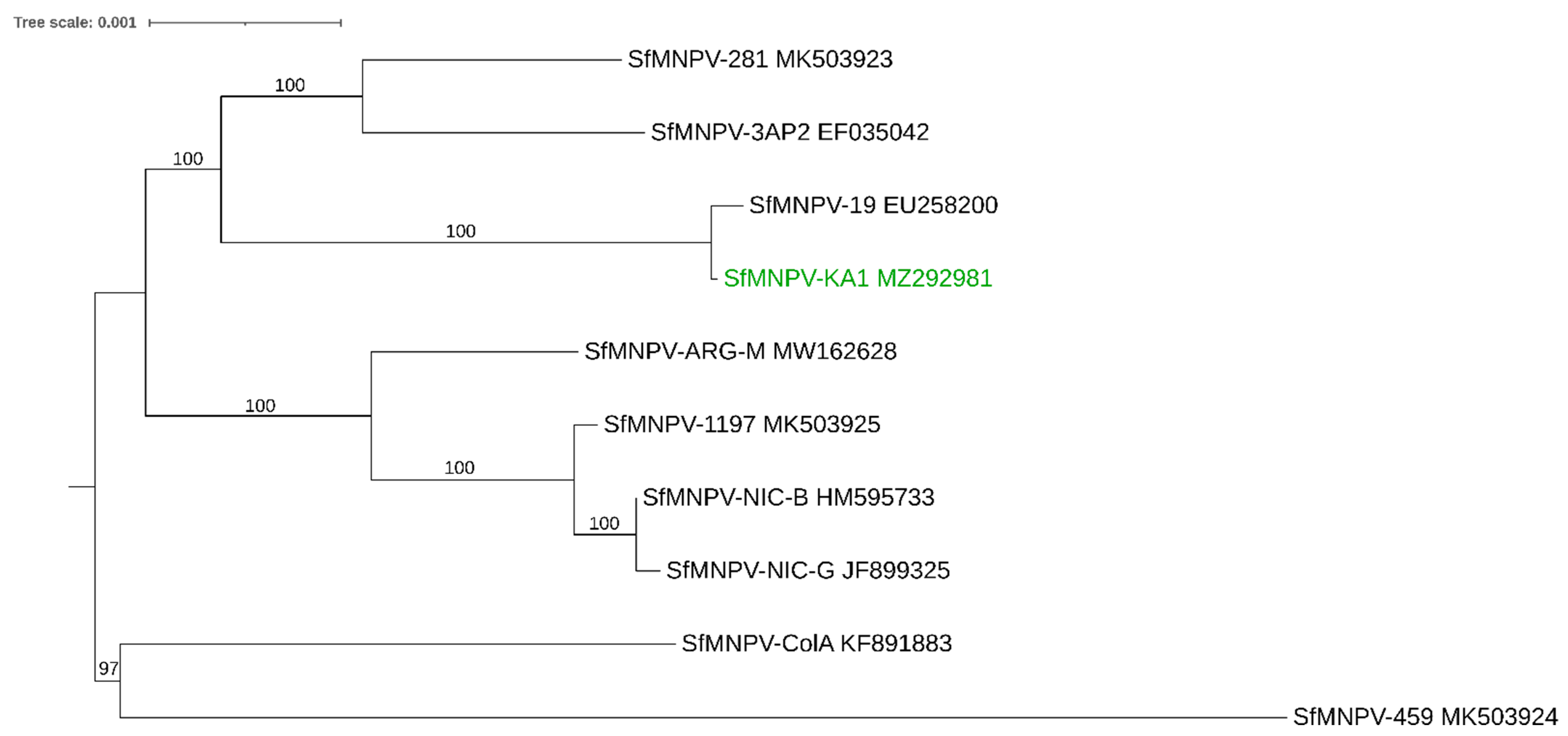
3. SfMNPV: The Most Promising Viral Candidate for the Biological Control of FAW
3.1. In Vitro and In Vivo Plaque Purification
3.2. Specificity of SfMNPV
3.3. Phylogenetic Analysis of SfMNPV and SfGV Isolates
4. Challenges of Using Baculoviruses for Biological Control
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Hruska, A.J.; Gould, F. Fall armyworm (Lepidoptera: Noctuidae) and Diatraea lineolata (Lepidoptera: Pyralidae): Impact of larval population level and temporal occurrence on maize yield in Nicaragua. J. Econ. Entomol. 1997, 90, 611–622. [Google Scholar] [CrossRef]
- Pashley, D.P. Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae): A sibling species complex? Ann. Entomol. Soc. Am. 1986, 79, 898–904. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Meagher, R.L. Review of fall armyworm (Lepidoptera: Noctuidae) genetic complexity and migration. Fla. Entomol. 2008, 91, 546–554. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- FAO FAW Map|Global Action for fall Armyworm Control | Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/fall-armyworm/monitoring-tools/faw-map/en/ (accessed on 4 May 2021).
- Prasanna, E.B.; Huesing, J.E.; Eddy, R.; Peschke, V.M. Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st ed.; CDMX CIMMYT: Veracruz, Mexico, 2018. [Google Scholar]
- CABI. Spodoptera frugiperda (Fall Armyworm) Datasheet. Invasive Species Compendium. Available online: https://www.cabi.org/isc/datasheet/29810 (accessed on 11 May 2021).
- Jing, D.P.; Guo, J.F.; Jiang, Y.Y.; Zhao, J.Z.; Sethi, A.; He, K.L.; Wang, Z.Y. Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques. Insect Sci. 2019, 4, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Kavita, H.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- EPPO. First Report of Spodoptera frugiperda in the Canary Islands, Spain. Available online: https://gd.eppo.int/reporting/article-6992 (accessed on 4 May 2021).
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.L.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Eschen, R.; Beale, T.; Bonnin, J.M.; Constantine, K.L.; Duah, S.; Finch, E.A.; Makale, F.; Nunda, W.; Ogunmodede, A.; Pratt, C.F.; et al. Towards estimating the economic cost of invasive alien species to African crop and livestock production. CABI Agric. Biosci. 2021, 2, 18. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Goergen, G.; Tounou, K.A.; Agboka, K.; Koffi, D.; Meagher, R.L. Analysis of strain distribution, migratory potential, and invasion history of fall armyworm populations in northern Sub-Saharan Africa. Sci. Rep. 2018, 8, 3710. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, R.N.; Koffi, D.; Agboka, K.; Tounou, K.A.; Banerjee, R.; Jurat-Fuentes, J.L.; Meagher, R.L. Comparative molecular analyses of invasive fall armyworm in Togo reveal strong similarities to populations from the eastern United States and the Greater Antilles. PLoS ONE 2017, 12, e0181982. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, R.N.; Htain, N.N.; Boughton, D.; Zhang, L.; Xiao, Y.; Nagoshi, B.Y.; Mota-Sanchez, D. Southeastern Asia fall armyworms are closely related to populations in Africa and India, consistent with common origin and recent migration. Sci. Rep. 2020, 10, 1421. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Dhanani, I.; Asokan, R.; Mahadevaswamy, H.M.; Kalleshwaraswamy, C.M.; Sharanabasappa; Meagher, R.L. Genetic characterization of fall armyworm infesting South Africa and India indicate recent introduction from a common source population. PLoS ONE 2019, 14, e0217755. [Google Scholar] [CrossRef]
- Schlum, K.A.; Lamour, K.; de Bortoli, C.P.; Banerjee, R.; Meagher, R.; Pereira, E.; Murua, M.G.; Sword, G.A.; Tessnow, A.E.; Viteri Dillon, D.; et al. Whole genome comparisons reveal panmixia among fall armyworm (Spodoptera frugiperda) from diverse locations. BMC Genom. 2021, 22, 179. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wyckhuys, K.A.G.; Jia, X.; Nie, F.; Wu, K. Fall armyworm invasion heightens pesticide expenditure among Chinese smallholder farmers. J. Environ. Manag. 2021, 282, 111949. [Google Scholar] [CrossRef]
- Chimweta, M.; Nyakudya, I.W.; Jimu, L.; Bray Mashingaidze, A. Fall armyworm [Spodoptera frugiperda (J.E. Smith)] damage in maize: Management options for flood-recession cropping smallholder farmers. Int. J. Pest Manag. 2019, 66, 142–154. [Google Scholar] [CrossRef]
- Yu, S.J. Insecticide resistance in the fall armyworm, Spodoptera frugiperda (J. E. Smith). Pestic. Biochem. Physiol. 1991, 39, 84–91. [Google Scholar] [CrossRef]
- Gutiérrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Terán-Santofimio, H.; Rodriguez-Maciel, J.C.; DiFonzo, C. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef]
- Muraro, D.S.; de Neto, D.O.A.; Kanno, R.H.; Kaiser, I.S.; Bernardi, O.; Omoto, C. Inheritance patterns, cross-resistance and synergism in Spodoptera frugiperda (Lepidoptera: Noctuidae) resistant to emamectin benzoate. Pest Manag. Sci. 2021, 77, 5049–5057. [Google Scholar] [CrossRef]
- Michigan State University. Arthropod Pesticide Resistance Database. Available online: https://www.pesticideresistance.org/display.php?page=species&arId=200 (accessed on 27 May 2021).
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; dos Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Bateman, M.L.; Day, R.K.; Rwomushana, I.; Subramanian, S.; Wilson, K.; Babendreier, D.; Luke, B.; Edgington, S. Updated assessment of potential biopesticide options for managing fall armyworm (Spodoptera frugiperda) in Africa. J. Appl. Entomol. 2021, 145, 384–393. [Google Scholar] [CrossRef]
- Hailu, G.; Niassy, S.; Zeyaur, K.R.; Ochatum, N.; Subramanian, S. Maize–legume intercropping and push–pull for management of fall armyworm, stemborers, and striga in Uganda. Agron. J. 2018, 110, 2513–2522. [Google Scholar] [CrossRef]
- Harrison, R.D.; Thierfelder, C.; Baudron, F.; Chinwada, P.; Midega, C.; Schaffner, U.; van den Berg, J. Agro-ecological options for fall armyworm (Spodoptera frugiperda J.E. Smith) management: Providing low-cost, smallholder friendly solutions to an invasive pest. J. Environ. Manag. 2019, 243, 318–330. [Google Scholar] [CrossRef]
- Prasad, V.; Srivastava, S. Insect Viruses. In Ecofriendly Pest Management for Food Security; Academic Press: London, UK, 2016; pp. 411–442. ISBN 9780128032664. [Google Scholar]
- Moscardi, F.; De Souza, M.L.; De Castro, M.E.B.; Lara Moscardi, M.; Szewczyk, B. Baculovirus pesticides: Present state and future perspectives. In Microbes and Microbial Technology: Agricultural and Environmental Applications; Springer: New York, NY, USA, 2011; pp. 415–445. ISBN 9781441979308. [Google Scholar]
- Federici, B.A. Pathogens of insects. In Encyclopedia of Insects; Elsevier: Amsterdam, The Netherlands, 2009; pp. 757–765. ISBN 9780123741448. [Google Scholar]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus insecticides in Latin America: Historical overview, current status and future perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef]
- Valicente, F.H. Entomopathogenic Viruses. In Natural Enemies of Insect Pests in Neotropical Agroecosystems; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 137–150. [Google Scholar]
- Maciel-Vergara, G.; Ros, V.I.D. Viruses of insects reared for food and feed. J. Invertebr. Pathol. 2017, 147, 60–75. [Google Scholar] [CrossRef]
- Miller, D.; O’Reilly, D.; Dall, D. Insect pest control by viruses. In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 1999; pp. 842–849. [Google Scholar]
- Cherry, A.J.; Mercadier, G.; Meikle, W.; Castelo-Branco, M.; Schroer, S. The role of enthomopathogens in DBM biological control. In Improving Biocontrol of Plutella xylostella; Bordat, D., Kirk, A., Eds.; CIRAD: Montpellier, France, 2002. [Google Scholar]
- Federici, B.A.; Bigot, Y. Ascoviruses. In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 186–193. [Google Scholar]
- Moscardi, F. Assessment of the application of baculoviruses for control of lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef]
- Asgari, S.; Bideshi, D.K.; Bigot, Y.; Federici, B.A.; Cheng, X.-W. ICTV Virus Taxonomy Profile: Ascoviridae. J. Gen. Virol. 2017, 98, 4–5. [Google Scholar] [CrossRef]
- Eberle, K.E.; Wennmann, J.T.; Kleespies, R.G.; Jehle, J.A. Basic techniques in insect virology. In Manual of Techniques in Invertebrate Pathology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 15–74. [Google Scholar]
- Hamm, J.J.; Pair, S.D.; Marti, O.G. Incidence and host range of a new ascovirus isolated from fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 1986, 69, 524. [Google Scholar] [CrossRef]
- Xue, J.L.; Cheng, X.W. Comparative analysis of a highly variable region within the genomes of Spodoptera frugiperda ascovirus 1d (SfAV-1d) and SfAV-1a. J. Gen. Virol. 2011, 92, 2797–2802. [Google Scholar] [CrossRef]
- Lezama-Gutiérrez, R.; Hamm, J.J.; Molina-Ochoa, J.; López-Edwards, M.; Pescador-Rubio, A.; González-Ramirez, M.; Styer, E.L. Occurrence of entomopathogens of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Mexican states of Michoacán, Colima, Jalisco and Tamaulipas. Fla. Entomol. 2001, 84, 23. [Google Scholar] [CrossRef]
- Hamm, J.J.; Styer, E.L.; Federici, B.A. Comparison of field-collected ascovirus isolates by DNA hybridization, host range, and histopathology. J. Invertebr. Pathol. 1998, 72, 138–146. [Google Scholar] [CrossRef]
- Yu, H.; Yang, C.J.; Li, N.; Zhao, Y.; Chen, Z.M.; Yi, S.J.; Li, Z.Q.; Adang, M.J.; Huang, G.H. Novel strategies for the biocontrol of noctuid pests (Lepidoptera) based on improving ascovirus infectivity using Bacillus thuringiensis. Insect Sci. 2020, 28, 1452–1467. [Google Scholar] [CrossRef]
- Bohorova, N.; Cabrera, M.; Abarca, C.; Quintero, R.; Maciel, A.M.; Brito, R.M.; Hoisington, D.; Bravo, A. Susceptibility of four tropical lepidopteran maize pests to Bacillus thuringiensis CryI-type insecticidal toxins. J. Econ. Entomol. 1997, 90, 412–415. [Google Scholar] [CrossRef]
- Ríos-Díez, J.D.; Siegfried, B.; Saldamando-Benjumea, C.I. Susceptibility of Spodoptera frugiperda (Lepidoptera: Noctuidae) strains from central Colombia to Cry1Ab and Cry1Ac entotoxins of Bacillus thuringiensis. Southwest. Entomol. 2012, 37, 281–293. [Google Scholar] [CrossRef]
- Raghunandan, B.; Patel, N.; Dave, H.; Mehta, D. Natural occurrence of nucleopolyhedrovirus infecting fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in Gujarat, India. Entomol. Zool. Stud. 2019, 7, 1040–1043. [Google Scholar]
- Chen, Z.W.; Yang, Y.C.; Zhang, J.F.; Jin, M.H.; Xiao, Y.T.; Xia, Z.C.; Liu, Y.Y.; Yu, S.Z.; Yang, Y.Z.; Wang, Y.; et al. Susceptibility and tissue specificity of Spodoptera frugiperda to Junonia coenia densovirus. J. Integr. Agric. 2021, 20, 840–849. [Google Scholar] [CrossRef]
- Mutuel, D.; Ravallec, M.; Chabi, B.; Multeau, C.; Salmon, J.M.; Fournier, P.; Ogliastro, M. Pathogenesis of Junonia coenia densovirus in Spodoptera frugiperda: A route of infection that leads to hypoxia. Virology 2010, 403, 137–144. [Google Scholar] [CrossRef]
- Ma, H.; Galvin, T.A.; Glasner, D.R.; Shaheduzzaman, S.; Khan, A.S. Identification of a novel rhabdovirus in Spodoptera frugiperda cell lines. J. Virol. 2014, 88, 6576–6585. [Google Scholar] [CrossRef]
- Xu, P.; Yang, L.; Yang, X.; Li, T.; Graham, R.I.; Wu, K.; Wilson, K. Novel partiti-like viruses are conditional mutualistic symbionts in their normal lepidopteran host, African armyworm, but parasitic in a novel host, fall armyworm. PLoS Pathog. 2020, 16, e1008467. [Google Scholar] [CrossRef]
- Ros, V.I.D. Baculoviruses: General Features (Baculoviridae). In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 739–746. [Google Scholar]
- Harrison, R.L.; Herniou, E.A.; Jehle, J.A.; Theilmann, D.A.; Burand, J.P.; Becnel, J.J.; Krell, P.J.; van Oers, M.M.; Mowery, J.D.; Bauchan, G.R. ICTV virus taxonomy profile: Baculoviridae. J. Gen. Virol. 2018, 99, 1185–1186. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.; Hoover, K. Baculoviruses and other occluded insect viruses. In Insect Pathology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 73–131. [Google Scholar]
- Rohrmann, G. Introduction to the baculoviruses, their taxonomy, and evolution. In Baculovirus Molecular Biology; National Center for Biotechnology Information: Bethesda, MD, USA, 2019; pp. 1–21. [Google Scholar]
- Sajjan, D.B.; Hinchigeri, S.B. Structural organization of baculovirus occlusion bodies and protective role of multilayered polyhedron envelope protein. Food Environ. Virol. 2016, 8, 86–100. [Google Scholar] [CrossRef]
- Popham, H.J.R.; Nusawardani, T.; Bonning, B.C. Introduction to the use of baculoviruses as biological insecticides. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2016; Volume 1350, pp. 383–392. [Google Scholar]
- Simón, O.; Williams, T.; López-Ferber, M.; Caballero, P. Virus entry or the primary infection cycle are not the principal determinants of host specificity of Spodoptera spp. nucleopolyhedroviruses. J. Gen. Virol. 2004, 85, 2845–2855. [Google Scholar] [CrossRef]
- Thézé, J.; Bézier, A.; Periquet, G.; Drezen, J.-M.; Herniou, E.A. Paleozoic origin of insect large dsDNA viruses. Proc. Natl. Acad. Sci. USA 2011, 108, 15931. [Google Scholar] [CrossRef]
- Jakubowska, A.K.; Lynn, D.E.; Herrero, S.; Vlak, J.M.; Van Oers, M.M. Host-range expansion of Spodoptera exigua multiple nucleopolyhedrovirus to Agrotis segetum larvae when the midgut is bypassed. J. Gen. Virol. 2010, 91, 898–905. [Google Scholar] [CrossRef]
- Szewczyk, B.; Rabalski, L.; Krol, E.; Sihler, W.; Lobo De Souza, M. Baculovirus biopesticides-a safe alternative to chemical protection of plants. J. Biopestic. 2009, 2, 209–216. [Google Scholar]
- Cuartas-Otálora, P.E.; Gómez-Valderrama, J.A.; Ramos, A.E.; Barrera-Cubillos, G.P.; Villamizar-Rivero, L.F. Bio-insecticidal potential of nucleopolyhedrovirus and granulovirus mixtures to control the fall armyworm Spodoptera frugiperda (J.E. smith, 1797) (lepidoptera: Noctuidae). Viruses 2019, 11, 684. [Google Scholar] [CrossRef]
- Behle, R.W.; Popham, H.J.R. Laboratory and field evaluations of the efficacy of a fast-killing baculovirus isolate from Spodoptera frugiperda. J. Invertebr. Pathol. 2012, 109, 194–200. [Google Scholar] [CrossRef]
- Han, Y.; van Houte, S.; Drees, G.F.; van Oers, M.M.; Ros, V.I.D. Parasitic manipulation of host behaviour: Baculovirus SeMNPV EGT facilitates tree-top disease in Spodoptera exigua larvae by extending the time to death. Insects 2015, 6, 716–731. [Google Scholar] [CrossRef]
- Gasque, S.N.; van Oers, M.M.; Ros, V.I. Where the baculoviruses lead, the caterpillars follow: Baculovirus-induced alterations in caterpillar behaviour. Curr. Opin. Insect Sci. 2019, 33, 30–36. [Google Scholar] [CrossRef]
- Feng, G.; Yu, Q.; Hu, C.; Wang, Y.; Yuan, G.; Chen, Q.; Yang, K.; Pang, Y. Apoptosis is induced in the haemolymph and fat body of Spodoptera exigua larvae upon oral inoculation with Spodoptera litura nucleopolyhedrovirus. J. Gen. Virol. 2007, 88, 2185–2193. [Google Scholar] [CrossRef]
- Gelernter, W.D.; Federici, B.A. Isolation, identification, and determination of virulence of a nuclear polyhedrosis virus from the beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae). Environ. Entomol. 1986, 15, 240–245. [Google Scholar] [CrossRef]
- Murillo, R.; Muñoz, D.; Caballero, P. Host range and biological activity of three Spodoptera nucleopolyhedrovirus genotypic variants and the effect of tinopal LPW on the most active variant. Int. J. Pest Manag. 2010, 49, 147–153. [Google Scholar] [CrossRef]
- AgBiTech. AgBiTech Global Innovation. Harnessing Nature to Control Insect Pests. Available online: https://www.agbitech.com/innovation (accessed on 27 April 2021).
- Ministério da Agricultura Pecuária e Abastecimento. Registered SfMNPV Active Ingredient Products in Brazil. Available online: https://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons (accessed on 14 August 2021).
- CABI. BioProtection Portal. Available online: https://bioprotectionportal.com/ (accessed on 17 August 2021).
- US EPA. Biopesticide Active Ingredients. Available online: https://www.epa.gov/ingredients-used-pesticide-products/biopesticide-active-ingredients (accessed on 24 August 2021).
- Barrera, G.; Simón, O.; Villamizar, L.; Williams, T.; Caballero, P. Spodoptera frugiperda multiple nucleopolyhedrovirus as a potential biological insecticide: Genetic and phenotypic comparison of field isolates from Colombia. Biol. Control 2011, 58, 113–120. [Google Scholar] [CrossRef]
- Vieira, C.M.; Tuelher, E.S.; Valicente, F.H.; Wolff, J.L.C. Characterization of a Spodoptera frugiperda multiple nucleopolyhedrovirus isolate that does not liquefy the integument of infected larvae. J. Invertebr. Pathol. 2012, 111, 189–192. [Google Scholar] [CrossRef]
- Wolff, J.L.C.; Valicente, F.H.; Martins, R.; de Oliveira, J.V.C.; de Zanotto, P.M.A. Analysis of the genome of Spodoptera frugiperda nucleopolyhedrovirus (SfMNPV-19) and of the high genomic heterogeneity in group II nucleopolyhedroviruses. J. Gen. Virol. 2008, 89, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Popham, H.J.R.; Rowley, D.L.; Harrison, R.L. Differential insecticidal properties of Spodoptera frugiperda multiple nucleopolyhedrovirus isolates against corn-strain and rice-strain fall armyworm, and genomic analysis of three isolates. J. Invertebr. Pathol. 2021, 183, 107561. [Google Scholar] [CrossRef]
- Andermatt Biocontrol AG Technical Information: Littovir®. Available online: https://www.andermattbiocontrol.com/media/pdf/products/bio-insecticides/baculovirus/littovir/Technical_Information_Littovir.pdf (accessed on 17 June 2021).
- Fall Armyworm: Impacts and Implications for Africa. Evidence Note 2018. Available online: https://www.invasive-species.org/wp-content/uploads/sites/2/2019/02/FAW-Evidence-Note-October-2018.pdf (accessed on 17 June 2021).
- Kuzmin, I.V.; Novella, I.S.; Dietzgen, R.G.; Padhi, A.; Rupprecht, C.E. The rhabdoviruses: Biodiversity, phylogenetics, and evolution. Infect. Genet. Evol. 2009, 9, 541–553. [Google Scholar] [CrossRef]
- Longdon, B.; Obbard, D.J.; Jiggins, F.M. Sigma viruses from three species of Drosophila form a major new clade in the rhabdovirus phylogeny. Proc. R. Soc. B Biol. Sci. 2010, 277, 35–44. [Google Scholar] [CrossRef]
- Schroeder, L.; Mar, T.B.; Haynes, J.R.; Wang, R.; Wempe, L.; Goodin, M.M.; Goodin, M. Host range and population survey of Spodoptera frugiperda rhabdovirus. J. Virol. 2019, 93, e02028-18. [Google Scholar] [CrossRef] [PubMed]
- Longdon, B.; Murray, G.G.R.; Palmer, W.J.; Day, J.P.; Parker, D.J.; Welch, J.J.; Obbard, D.J.; Jiggins, F.M. The evolution, diversity, and host associations of rhabdoviruses. Virus Evol. 2015, 1, vev014. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Becnel, J.J.; Valles, S.M. RNA Viruses Infecting Pest Insects. In Insect Pathology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 133–170. [Google Scholar]
- Bedford, G.O. Biology and management of palm dynastid beetles: Recent advances. Annu. Rev. Entomol. 2013, 58, 353–372. [Google Scholar] [CrossRef] [PubMed]
- Bergoin, M.; Tijssen, P. Biological and molecular properties of densoviruses and their use in protein expression and biological control. In The Insect Viruses; Springer: New York, NY, USA, 1998; pp. 141–169. [Google Scholar]
- Pigeyre, L.; Schatz, M.; Ravallec, M.; Gasmi, L.; Nègre, N.; Clouet, C.; Seveno, M.; El Koulali, K.; Decourcelle, M.; Guerardel, Y.; et al. Interaction of a densovirus with glycans of the peritrophic matrix mediates oral infection of the lepidopteran pest Spodoptera frugiperda. Viruses 2019, 11, 870. [Google Scholar] [CrossRef]
- Jakubowska, A.K.; Nalcacioglu, R.; Millán-Leiva, A.; Sanz-Carbonell, A.; Muratoglu, H.; Herrero, S.; Demirbag, Z. In search of pathogens: Transcriptome-based identification of viral sequences from the pine processionary moth (Thaumetopoea pityocampa). Viruses 2015, 7, 456–479. [Google Scholar] [CrossRef]
- Käfer, S.; Paraskevopoulou, S.; Zirkel, F.; Wieseke, N.; Donath, A.; Petersen, M.; Jones, T.C.; Liu, S.; Zhou, X.; Middendorf, M.; et al. Re-assessing the diversity of negative strand RNA viruses in insects. PLoS Pathog. 2019, 15, e1008224. [Google Scholar] [CrossRef]
- Millán-Leiva, A.; Jakubowska, A.K.; Ferré, J.; Herrero, S. Genome sequence of SeIV-1, a novel virus from the Iflaviridae family infective to Spodoptera exigua. J. Invertebr. Pathol. 2012, 109, 127–133. [Google Scholar] [CrossRef]
- Obbard, D.J.; Shi, M.; Roberts, K.E.; Longdon, B.; Dennis, A.B. A new lineage of segmented RNA viruses infecting animals. Virus Evol. 2020, 6, 61. [Google Scholar] [CrossRef]
- Obbard, D.J.; Palacios, G.; Kuhn, J.H. Expansion of the metazoan virosphere: Progress, pitfalls, and prospects. Curr. Opin. Virol. 2018, 31, 17–23. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Shi, C.; Liu, Y.; Hu, X.; Xiong, J.; Zhang, B.; Yuan, Z. A metagenomic survey of viral abundance and diversity in mosquitoes from hubei province. PLoS ONE 2015, 10, e0129845. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Kim, Y.S.; Wang, Y.; Shin, S.W.; Kim, I.; Tao, X.Y.; Liu, Q.; Roh, J.Y.; Kim, J.S.; Je, Y.H. Complete genome sequence of a novel picorna-like virus isolated from Spodoptera exigua. J. Asia Pac. Entomol. 2012, 15, 259–263. [Google Scholar] [CrossRef]
- Frey, K.G.; Biser, T.; Hamilton, T.; Santos, C.J.; Pimentel, G.; Mokashi, V.P.; Bishop-Lilly, K.A. Bioinformatic characterization of mosquito viromes within the Eastern United States and Puerto Rico: Discovery of novel viruses. Evol. Bioinf. 2016, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fauver, J.R.; Grubaugh, N.D.; Krajacich, B.J.; Weger-Lucarelli, J.; Lakin, S.M.; Fakoli, L.S.; Bolay, F.K.; Diclaro, J.W.; Dabiré, K.R.; Foy, B.D.; et al. West African Anopheles gambiae mosquitoes harbor a taxonomically diverse virome including new insect-specific flaviviruses, mononegaviruses, and totiviruses. Virology 2016, 498, 288–299. [Google Scholar] [CrossRef]
- Carballo, A.; Williams, T.; Murillo, R.; Caballero, P. Iflavirus covert infection increases susceptibility to nucleopolyhedrovirus disease in Spodoptera exigua. Viruses 2020, 12, 509. [Google Scholar] [CrossRef] [PubMed]
- Virto, C.; Navarro, D.; Tellez, M.M.; Herrero, S.; Williams, T.; Murillo, R.; Caballero, P. Natural populations of Spodoptera exigua are infected by multiple viruses that are transmitted to their offspring. J. Invertebr. Pathol. 2014, 122, 22–27. [Google Scholar] [CrossRef]
- Carballo, A.; Murillo, R.; Jakubowska, A.; Herrero, S.; Williams, T.; Caballero, P. Co-infection with iflaviruses influences the insecticidal properties of Spodoptera exigua multiple nucleopolyhedrovirus occlusion bodies: Implications for the production and biosecurity of baculovirus insecticides. PLoS ONE 2017, 12, e0177301. [Google Scholar] [CrossRef]
- García-Banderas, D.; Tamayo-Mejía, F.; Pineda, S.; de la Rosa, J.I.F.; Lasa, R.; Chavarrieta-Yáñez, J.M.; Gervasio-Rosas, E.; Zamora-Avilés, N.; Morales, S.I.; Ramos-Ortiz, S.; et al. Biological characterization of two Spodoptera frugiperda nucleopolyhedrovirus isolates from Mexico and evaluation of one isolate in a small-scale field trial. Biol. Control 2020, 149, 104316. [Google Scholar] [CrossRef]
- Harrison, R.L.; Puttler, B.; Popham, H.J.R. Genomic sequence analysis of a fast-killing isolate of Spodoptera frugiperda multiple nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 775–790. [Google Scholar] [CrossRef]
- Ordóñez-García, M.; Rios-Velasco, C.; Berlanga-Reyes, D.I.; Acosta-Muñiz, C.H.; Salas-Marina, M.Á.; Cambero-Campos, O.J. Occurrence of natural enemies of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Chihuahua, Mexico. Fla. Entomol. 2015, 98, 843–847. [Google Scholar] [CrossRef]
- Escribano, A.; Williams, T.; Goulson, D.; Cave, R.D.; Chapman, J.W.; Caballero, P. Selection of a nucleopolyhedrovirus for Control of Spodoptera frugiperda (Lepidoptera: Noctuidae): Structural, genetic, and biological comparison of four isolates from the Americas. J. Econ. Entomol. 1999, 92, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Fuxa, J.R. Spodoptera frugiperda susceptibility to nuclear polyhedrosis virus isolates with reference to insect migration. Environ. Entomol. 1987, 16, 218–223. [Google Scholar] [CrossRef]
- Lei, C.; Yang, J.; Wang, J.; Hu, J.; Sun, X. Molecular and biological characterization of Spodoptera frugiperda multiple nucleopolyhedrovirus field isolate and genotypes from China. Insects 2020, 11, 777. [Google Scholar] [CrossRef]
- Firake, D.M.; Behere, G.T. Natural mortality of invasive fall armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) in maize agroecosystems of northeast India. Biol. Control 2020, 148, 104303. [Google Scholar] [CrossRef]
- Sivakumar, G.; Kannan, M.; Ramesh Babu, S.; Mohan, M.; Sampath Kumar, M.; Raveendran, P.; Venkatesan, T.; Rangeshwaran, R.; Chandish, R.B.; RamKumar, P. Isolation and characterization of indigenous nucleopolyhedrovirus infecting fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in India. Curr. Sci. 2020, 119, 860–864. [Google Scholar] [CrossRef]
- Wennmann, J.; Tepa-Yotto, G.; Jehle, J.; Goergen, G. Genome Sequence of a Spodoptera frugiperda multiple nucleopolyhedrovirus isolated from fall armyworm (Spodoptera frugiperda) in Nigeria, West Africa. Microbiol. Resour. Announc. 2021, 10, e00565-21. [Google Scholar] [CrossRef] [PubMed]
- Barreto, M.R.; Guimaraes, C.T.; Teixeira, F.F.; Paiva, E.; Valicente, F.H. Effect of baculovirus Spodoptera isolates in Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) larvae and their characterization by RAPD. Neotrop. Entomol. 2005, 34, 67–75. [Google Scholar] [CrossRef]
- Niz, J.M.; Salvador, R.; Ferrelli, M.L.; Sciocco De Cap, A.; Romanowski, V.; Berretta, M.F. Genetic variants in Argentinean isolates of Spodoptera frugiperda Multiple Nucleopolyhedrovirus. Virus Genes 2020, 56, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Simón, O.; Williams, T.; López-Ferber, M.; Caballero, P. Genetic structure of a Spodoptera frugiperda nucleopolyhedrovirus population: High prevalence of deletion genotypes. Appl. Environ. Microbiol. 2004, 70, 5579–5588. [Google Scholar] [CrossRef]
- Barrera, G.; Williams, T.; Villamizar, L.; Caballero, P.; Simón, O. Deletion genotypes reduce occlusion body potency but increase occlusion body production in a Colombian Spodoptera frugiperda nucleopolyhedrovirus population. PLoS ONE 2013, 8, e77271. [Google Scholar] [CrossRef]
- Rowley, D.L.; Farrar, R.R.; Blackburn, M.B.; Harrison, R.L. Genetic and biological variation among nucleopolyhedrovirus isolates from the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Virus Genes 2010, 40, 458–468. [Google Scholar] [CrossRef]
- Ordóñez-García, M.; Rios-Velasco, C.; de Ornelas-Paz, J.J.; Bustillos-Rodríguez, J.C.; Acosta-Muñiz, C.H.; Berlanga-Reyes, D.I.; Salas-Marina, M.Á.; Cambero-Campos, O.J.; Gallegos-Morales, G. Molecular and morphological characterization of multiple nucleopolyhedrovirus from Mexico and their insecticidal activity against Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Appl. Entomol. 2020, 144, 123–132. [Google Scholar] [CrossRef]
- Ferrelli, M.L.; Pidre, M.L.; Ghiringhelli, P.D.; Torres, S.; Fabre, M.L.; Masson, T.; Cédola, M.T.; Sciocco-Cap, A.; Romanowski, V. Genomic analysis of an Argentinean isolate of Spodoptera frugiperda granulovirus reveals that various baculoviruses code for Lef-7 proteins with three F-box domains. PLoS ONE 2018, 13, e0202598. [Google Scholar] [CrossRef] [PubMed]
- Cuartas, P.E.; Barrera, G.P.; Belaich, M.N.; Barreto, E.; Ghiringhelli, P.D.; Villamizar, L.F. The complete sequence of the first Spodoptera frugiperda betabaculovirus genome: A natural multiple recombinant virus. Viruses 2015, 7, 394–421. [Google Scholar] [CrossRef]
- Cuartas, P.; Barrera, G.; Barreto, E.; Villamizar, L. Characterisation of a Colombian granulovirus (Baculoviridae: Betabaculovirus) isolated from Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae. Biocontrol Sci. Technol. 2014, 24, 1265–1285. [Google Scholar] [CrossRef]
- Simón, O.; Williams, T.; López-Ferber, M.; Caballero, P. Deletion of egt is responsible for the fast-killing phenotype of natural deletion genotypes in a Spodoptera frugiperda multiple nucleopolyhedrovirus population. J. Invertebr. Pathol. 2012, 111, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Simón, O.; Palma, L.; Beperet, I.; Muñoz, D.; López-Ferber, M.; Caballero, P.; Williams, T. Sequence comparison between three geographically distinct Spodoptera frugiperda multiple nucleopolyhedrovirus isolates: Detecting positively selected genes. J. Invertebr. Pathol. 2011, 107, 33–42. [Google Scholar] [CrossRef]
- Barrera, G.P.; Belaich, M.N.; Patarroyo, M.A.; Villamizar, L.F.; Ghiringhelli, P.D. Evidence of recent interspecies horizontal gene transfer regarding nucleopolyhedrovirus infection of Spodoptera frugiperda. BMC Genom. 2015, 16, 1008. [Google Scholar] [CrossRef] [PubMed]
- Masson, T.; Fabre, M.L.; Pidre, M.L.; Niz, J.M.; Berretta, M.F.; Romanowski, V.; Ferrelli, M.L. Genomic diversity in a population of Spodoptera frugiperda nucleopolyhedrovirus. Infect. Genet. Evol. 2021, 90, 104749. [Google Scholar] [CrossRef] [PubMed]
- SfMNPV. Product Formulations in Brazil. Available online: https://agrofit.agricultura.gov.br/agrofit_cons/!ap_produto_form_consulta_cons (accessed on 14 August 2021).
- EU. Pesticides Database (v.2.2) Active substance. Available online: https://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/active-substances/?event=as.details&as_id=1173 (accessed on 17 August 2021).
- CABI. BioProtection Portal. Indústria e Comércio de Fertilizantes e Insumos Microbiológicos Ltd.a. Available online: https://app.bioprotectionportal.com/brazil/product?regnum=39817&distributor=undefined&company=Simbiose (accessed on 11 August 2021).
- Andermatt Biocontrol AG Spodovir Plus—Controls Fall Armyworms (Spodoptera frugiperda). Available online: https://www.andermattbiocontrol.com/sites/products/bio-insecticides/spodovir-plus.html (accessed on 24 June 2021).
- Simón, O.; Chevenet, F.; Williams, T.; Caballero, P.; López-Ferber, M. Physical and partial genetic map of Spodoptera frugiperda nucleopolyhedrovirus (SfMNPV) genome. Virus Genes 2005, 30, 403–417. [Google Scholar] [CrossRef]
- Shapiro, D.I.; Fuxa, J.R.; Braymer, H.D.; Pashley, D.P. DNA restriction polymorphism in wild isolates of Spodoptera frugiperda nuclear polyhedrosis virus. J. Invertebr. Pathol. 1991, 58, 96–105. [Google Scholar] [CrossRef]
- Serrano, A.; Williams, T.; Simón, O.; López-Ferber, M.; Caballero, P.; Muñoz, D. Analagous population structures for two alphabaculoviruses highlight a functional role for deletion mutants. Appl. Environ. Microbiol. 2013, 79, 1118–1125. [Google Scholar] [CrossRef]
- Wilson, K.R.; O’Reilly, D.R.; Hails, R.S.; Cory, J.S. Age-related effects of the Autographa californica multiple nucleopolyhedrovirus egt gene in the cabbage looper (Trichoplusia ni). Biol. Control 2000, 19, 57–63. [Google Scholar] [CrossRef]
- O’Reilly, D.R.; Miller, L.K. A baculovirus blocks insect molting by producing ecdysteroid UDP-glucosyl transferase. Science 1989, 245, 1110–1112. [Google Scholar] [CrossRef]
- Goulson, D. Can host susceptibility to baculovirus infection be predicted from host taxonomy or life history? Environ. Entomol. 2003, 32, 61–70. [Google Scholar] [CrossRef]
- Doyle, C.J.; Hirst, M.L.; Cory, J.S.; Entwistle, P.F. Risk assessment studies: Detailed host range testing of wild-type cabbage moth, Mamestra brassicae (Lepidoptera: Noctuidae) nuclear polyhedrosis virus. Appl. Environ. Microbiol. 1990, 56, 2704–2710. [Google Scholar] [CrossRef] [PubMed]
- Cisneros, J.; Pérez, J.A.; Penagos, D.I.; Ruiz, V.J.; Goulson, D.; Caballero, P.; Cave, R.D.; Williams, T. Formulation of a nucleopolyhedrovirus with boric acid for control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize. Biol. Control 2002, 23, 87–95. [Google Scholar] [CrossRef]
- Armenta, R.; Martínez, A.M.; Chapman, J.W.; Magallanes, R.; Goulson, D.; Caballero, P.; Cave, R.D.; Cisneros, J.; Valle, J.; Castillejos, V.; et al. Impact of a nucleopolyhedrovirus bioinsecticide and selected synthetic insecticides on the abundance of insect natural enemies on maize in Southern Mexico. J. Econ. Entomol. 2003, 96, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.; Goulson, D.; Caballero, P.; Cisneros, J.; Martínez, A.M.; Chapman, J.W.; Roman, D.X.; Cave, R.D. Evaluation of a baculovirus bioinsecticide for small-scale maize growers in Latin America. Biol. Control 1999, 14, 67–75. [Google Scholar] [CrossRef]
- Méndez, W.A.; Valle, J.; Ibarra, J.E.; Cisneros, J.; Penagos, D.I.; Williams, T. Spinosad and nucleopolyhedrovirus mixtures for control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize. Biol. Control 2002, 25, 195–206. [Google Scholar] [CrossRef]
- Figueroa, J.I.; Coronado, R.E.; Pineda, S.; Chavarrieta, J.M.; Martínez-Castillo, A.M. Mortality and food consumption in Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae treated with spinosad alone or in mixtures with a nucleopolyhedrovirus. Fla. Entomol. 2015, 98, 1009–1011. [Google Scholar] [CrossRef]
- Guido-Cira, N.D.; Tamez-Guerra, P.; Mireles-Martínez, M.; Villegas-Mendoza, J.M.; Rosas-García, N.M. Activity of Bacillus thuringiensis- and baculovirus-based formulations to Spodoptera species. Southwest. Entomol. 2017, 42, 191–201. [Google Scholar] [CrossRef]
- Farrar, R.R.; Shapiro, M.; Shepard, B.M. Activity of the nucleopolyhedrovirus of the fall armyworm (Lepidoptera: Noctuidae) on foliage of transgenic sweet corn expressing a CryIa(b) toxin. Environ. Entomol. 2004, 33, 982–989. [Google Scholar] [CrossRef][Green Version]
- Farrar, R.R.; Shapiro, M.; Shepard, B.M. Enhanced activity of the nucleopolyhedrovirus of the fall armyworm (Lepidoptera: Noctuidae) on Bt-transgenic and nontransgenic sweet corn with a fluorescent brightener and a feeding stimulant. Environ. Entomol. 2005, 34, 825–832. [Google Scholar] [CrossRef]
- Farrar, R.R.; Merle Shepard, B.; Shapiro, M.; Hassell, R.L.; Schaffer, M.L.; Smith, C.M. Supplemental control of lepidopterous pests on Bt transgenic sweet corn with biologically-based spray treatments. J. Insect Sci. 2009, 9, 1536–2442. [Google Scholar] [CrossRef] [PubMed]
- Erlandson, M.; Newhouse, S.; Moore, K.; Janmaat, A.; Myers, J.; Theilmann, D. Characterization of baculovirus isolates from Trichoplusia ni populations from vegetable greenhouses. Biol. Control 2007, 41, 256–263. [Google Scholar] [CrossRef]
- Simón, O.; Palma, L.; Williams, T.; López-Ferber, M.; Caballero, P. Analysis of a naturally-occurring deletion mutant of Spodoptera frugiperda multiple nucleopolyhedrovirus reveals sf58 as a new per os infectivity factor of lepidopteran-infecting baculoviruses. J. Invertebr. Pathol. 2012, 109, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*, Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Swofford, D.L.; Sullivan, J. Phylogeny inference based on parsimony and other methods using PAUP. In The Phylogenetic Handbook. A Practical Approach to Phylogenetic Analysis and Hypothesis Testing.; Lemey, P., Salemi, M., Vandamme, A.-M., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 267–312. [Google Scholar]
- Wang, P.; Granado, R.R. An intestinal mucin is the target substrate for a baculovirus enhancin. Proc. Natl. Acad. Sci. USA 1997, 94, 6977–6982. [Google Scholar] [CrossRef]
- Cubillos, G.P.B.; Gómez-Valderrama, J.A.; Rivero, L.F.V. Efficacy of microencapsulated nucleopolyhedroviruses from Colombia as biological insecticides against Spodoptera frugiperda (Lepidoptera: Noctuidae). Acta Agron. 2017, 66, 267–274. [Google Scholar] [CrossRef]
- Gómez, J.; Guevara, J.; Cuartas, P.; Espinel, C.; Villamizar, L. Microencapsulated Spodoptera frugiperda nucleopolyhedrovirus: Insecticidal activity and effect on arthropod populations in maize. Biocontrol Sci. Technol. 2013, 23, 829–846. [Google Scholar] [CrossRef]
- Wilson, K.; Grzywacz, D.; Curcic, I.; Scoates, F.; Harper, K.; Rice, A.; Paul, N.; Dillon, A. A novel formulation technology for baculoviruses protects biopesticide from degradation by ultraviolet radiation. Sci. Rep. 2020, 10, 13301. [Google Scholar] [CrossRef] [PubMed]
- Villamizar, L.; Barrera, G.; Cotes, A.M.; Martínez, F. Eudragit S100 microparticles containing Spodoptera frugiperda nucleopolyehedrovirus: Physicochemical characterization, photostability and in vitro virus release. J. Microencapsul. 2010, 27, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Poshadri, A.; Kuna, A. Microencapsulation technology: A review. J. Res. ANGRAU 2010, 38, 86–102. [Google Scholar]
- Castillejos, V.; Trujillo, J.; Ortega, L.D.; Santizo, J.A.; Cisneros, J.; Penagos, D.I.; Valle, J.; Williams, T. Granular phagostimulant nucleopolyhedrovirus formulations for control of Spodoptera frugiperda in maize. Biol. Control 2002, 24, 300–310. [Google Scholar] [CrossRef]
- Hawtin, R.E.; Zarkowska, T.; Arnold, K.; Thomas, C.J.; Gooday, G.W.; King, L.A.; Kuzio, J.A.; Possee, R.D. Liquefaction of Autographa californica nucleopolyhedrovirus-infected insects is dependent on the integrity of virus-encoded chitinase and cathepsin genes. Virology 1997, 238, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.; Chan, L.; van Oers, M.M. Production of Entomopathogenic Viruses. In Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens; Academic Press: Cambridge, MA, USA, 2014; pp. 437–482. [Google Scholar]
- De Almeida, A.F.; de Macedo, G.R.; Chan, L.C.L.; da Pedrini, M.R.S. Kinetic analysis of in vitro production of wild-type Spodoptera frugiperda nucleopolyhedrovirus. Braz. Arch. Biol. Technol. 2010, 53, 285–291. [Google Scholar] [CrossRef][Green Version]
- Sihler, W.; de Souza, M.L.; Valicente, F.H.; Falcão, R.; Sanches, M.M. In vitro infectivity of Spodoptera frugiperda multiple nucleopolyhedrovirus to different insect cell lines. Pesqui. Agropecuária Bras. 2018, 53, 1–9. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.W.; Heckel, D.G.; Huber, J.; et al. Rapid emergence of baculovirus resistance in codling moth due to dominant, sex-linked inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Schulze-Bopp, S.; Undorf-Spahn, K.; Fritsch, E. Evidence for a second type of resistance against Cydia pomonella granulovirus in field populations of codling moths. Appl. Env. Microbiol. 2016, 83, e02330-16. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.J.; Schulze-Bopp, S.; Fritsch, E.; Undorf-Spahn, K.; Jehle, J.A. A third type of resistance to Cydia pomonella granulovirus in codling moths shows a mixed Z-linked and autosomal inheritance pattern. Appl. Environ. Microbiol. 2017, 83, e01036-17. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.M.; Eberle, K.E.; Radtke, P.; Jehle, J.A. Baculovirus resistance in codling moth is virus isolate-dependent and the consequence of a mutation in viral gene pe38. Proc. Natl. Acad. Sci. USA 2014, 111, 15711–15716. [Google Scholar] [CrossRef]
- Fan, J.; Jehle, J.A.; Wennmann, J.T. Population structure of Cydia pomonella granulovirus isolates revealed by quantitative analysis of genetic variation. Virus Evol. 2021, 7, veaa073. [Google Scholar] [CrossRef]
- Alletti, G.G.; Sauer, A.J.; Weihrauch, B.; Fritsch, E.; Undorf-Spahn, K.; Wennmann, J.T.; Jehle, J.A. Using next generation sequencing to identify and quantify the genetic composition of resistance-breaking commercial isolates of Cydia pomonella Granulovirus. Viruses 2017, 9, 250. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Mitchell, F.L.; Richter, A.R. Resistance of Spodoptera frugiperda [Lep.: Noctuidae] to a nuclear polyhedrosis virus in the field and laboratory. Entomophaga 1988, 33, 55–63. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Richter, A.R. Reversion of resistance by Spodoptera frugiperda to nuclear polyhedrosis virus. J. Invertebr. Pathol. 1989, 53, 52–56. [Google Scholar] [CrossRef]
- Carradore, R. Making viruses more socially acceptable: A sociological framework for virus-based plant protection products. Teor. Praksa 2019, 56, 798–814. [Google Scholar]
- Cherry, A.J.; Gwynn, R.L. Perspectives on the development of biological control agents in Africa. Biocontrol Sci. Technol. 2007, 17, 665–676. [Google Scholar] [CrossRef]
- Skovmand, O. Microbial control in Southeast Asia. J. Invertebr. Pathol. 2007, 95, 168–174. [Google Scholar] [CrossRef]
- Lacey, L.A.; Shapiro-Ilan, D.I. Microbial control of insect pests in temperate orchard systems: Potential for incorporation into IPM. Annu. Rev. Entomol. 2008, 53, 121–144. [Google Scholar] [CrossRef]
- Moore, S.D.; Kirkman, W.; Richards, G.I.; Stephen, P.R. The Cryptophlebia leucotreta granulovirus—10 Years of commercial field use. Viruses 2015, 7, 1284. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, D. Basic and Applied Research: Baculovirus. In Microbial Control of Insect and Mite Pests: From Theory to Practice; Academic Press: Cambridge, MA, USA, 2017; pp. 27–46. [Google Scholar]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Ravensberg, W.J. Critical Factors in the Successful Commercialization of Microbial Pest Control Products. In A Roadmap to the Successful Development and Commercialization of Microbial Pest Control Products for Control of Arthropods; Springer: Berlin/Heidelberg, Germany, 2011; pp. 295–356. [Google Scholar]


{kind=link}
| Viruses Associated with FAW | Virus Name | Host Stage Infected | Main Symptoms | Key References |
|---|---|---|---|---|
| DNA Viruses | ||||
| Ascoviruses | Spodoptera frugiperda ascovirus (SfAV-1a) | Larvae | Stunting of infected larvae, production of virus filled vesicles, milky-white discoloration, fat body cells infection | [39,43] |
| Baculoviruses | Spodoptera frugiperda multiple nucleopolyhedrovirus (SfMNPV), Spodoptera frugiperda granulovirus (SfGV), Spodoptera littoralis nucleopolyhedrovirus (SpliNPV) | Larvae | Whitish-grey discoloration, swollen body, ruptured integument leading to liquefaction of the larvae | [34,35,50] |
| Densoviruses | Junonia coenia densovirus (JcDV) | Larvae | Anorexia, lethargy, hypoxia, and inhibition of molting | [42,51,52] |
| RNA Viruses | ||||
| Rhabdoviruses | Spodoptera frugiperda rhabdovirus | Sf9 and Sf21 cells | No described symptoms | [53] |
| Partiti-like viruses | Spodoptera exempta virus 1, 2, and 3 (SEIV1-3) | Larvae | Reduced the growth rate and fecundity of FAW larvae and increase susceptibility to baculovirus | [54] |
| SfMNPV | SeMNPV | SpltNPV | SpliNPV | |
|---|---|---|---|---|
| S. frugiperda | +++ | +++ | ||
| S. exigua | + | +++ | +++ | |
| S. litura | +++ | |||
| S. littoralis | + | +++ |
| Baculovirus | Isolate | Plaque Purified Isolate/Genotype | Country of Collection | Key Reference |
|---|---|---|---|---|
| SfMNPV | 1 | 1BP2 | USA | [104] |
| 2 | 2AP2 | USA | [104] | |
| 3 | 3AP2 | USA | [104] | |
| 4 | 4AP2 | USA | [104] | |
| 5 | 5AP1 | USA | [104] | |
| 6 | 6AP1 | USA | [104] | |
| 01 to 22 | Brazil | [78,112] | ||
| 6 nd | Brazil | [77] | ||
| SfHub | SfHub-A, SfHub-E | China | [108] | |
| M | M1, M11 | Argentina | [113] | |
| C | Argentina | [113] | ||
| SfNIC | SfNIC-A to SfNIC-I | Nicaragua | [76,114] | |
| SfCol | SfCol-A to SfCol-G | Colombia | [76,115] | |
| 281, 637, 638, 651, 652, 653, 654, 1197, 2507, 3146 | USA | [79] | ||
| 459, 635, 636 | Colombia | [116] | ||
| SfCH1, SfCH4, SfCH6, SfCH12, SfCH15, SfCH18, SfCH30, SfCH32 | Mexico | [105,117] | ||
| SfGV | ARG | Argentina | [118] | |
| VG008 | Colombia | [119] |
| Product Name | Trademark | Baculovirus Isolate | Country of Registration | Key References |
|---|---|---|---|---|
| Baculomip-SF | Promip Manejo Integrado de Pragas Ltd.a | SfMNPV | Brazil | [125] |
| Baculonat SF | Bionat Soluções Biologicas Ltd.a | SfMNPV | Brazil | [125] |
| CartuchoVIT | Grupo Vitae Ltd.a—ME | SfMNPV-6 | Brazil | [35,73] |
| Cartugen Cartugen CCAB | Agbitech Controles Biológicos Ltd.a | SfMNPV-3AP2 | Brazil | [72,73] |
| Fawligen | AgBiTech Pty Ltd. | SfMNPV-3AP2 | Bangladesh, Kenya, Sri Lanka, USA, Zambia, Australia | [72,74,75] |
| Laphy Protection | Biome Industry Commerce and Distribution—EIRELI | SfMNPV | Brazil | [125] |
| Littovir | Andermatt Biocontrol | SpliNPV | France, Portugal, Tunisia, Bulgaria, Spain, Italy, Morocco, Cameroon | [80,81,126,127] |
| Lungo | Agbitech Controles Biológicos Ltd.a | SfMNPV | Brazil | [125] |
| Spobiol | Colombian Agricultural Research Corporation | SfMNPV | [74] | |
| Spodovir Plus | Andermatt Biocontrol | SfMNPV-6 and -19 | Brazil, Paraguay | [125,128] |
| Surtivo Plus Surtivo Ultra | AgBiTech Pty Ltd. | Autographa californica multiple nucleopolyhedrovirus (AcMNPV), Chrysodeixis includens (ChinNPV), Helicoverpa armigera HearNPV and SfMNPV | Brazil | [73,74] |
| VirControl SF | Symbiosis Industry and Trade of Fertilizers and Microbiological Inputs Ltd.a | SfMNPV-6 | Brazil | [35,73,74] |
| Vir Protection | Biome Industry Commerce and Distribution—EIRELI | SfMNPV | Brazil | [74,125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, A.G.; Wennmann, J.T.; Goergen, G.; Bryon, A.; Ros, V.I.D. Viruses of the Fall Armyworm Spodoptera frugiperda: A Review with Prospects for Biological Control. Viruses 2021, 13, 2220. https://doi.org/10.3390/v13112220
Hussain AG, Wennmann JT, Goergen G, Bryon A, Ros VID. Viruses of the Fall Armyworm Spodoptera frugiperda: A Review with Prospects for Biological Control. Viruses. 2021; 13(11):2220. https://doi.org/10.3390/v13112220
Chicago/Turabian StyleHussain, Ahmed G., Jörg T. Wennmann, Georg Goergen, Astrid Bryon, and Vera I.D. Ros. 2021. "Viruses of the Fall Armyworm Spodoptera frugiperda: A Review with Prospects for Biological Control" Viruses 13, no. 11: 2220. https://doi.org/10.3390/v13112220
APA StyleHussain, A. G., Wennmann, J. T., Goergen, G., Bryon, A., & Ros, V. I. D. (2021). Viruses of the Fall Armyworm Spodoptera frugiperda: A Review with Prospects for Biological Control. Viruses, 13(11), 2220. https://doi.org/10.3390/v13112220