
Imbalanced Immune Response of T-Cell and B-Cell Subsets in Patients with Moderate and Severe COVID-19

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Characteristics
2.2. Sample Collection
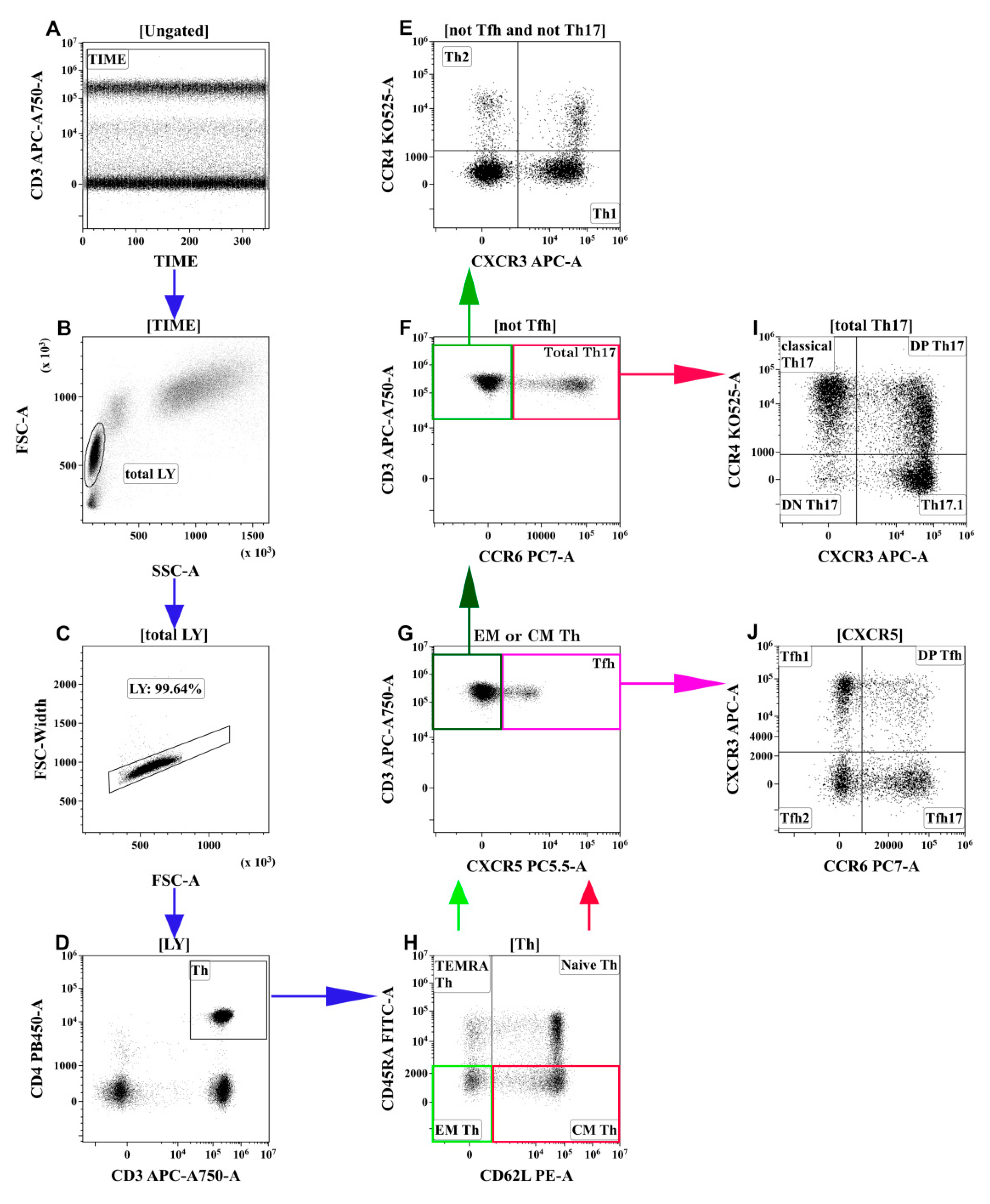
2.3. T-Cell Immunophenotyping by Flow Cytometry
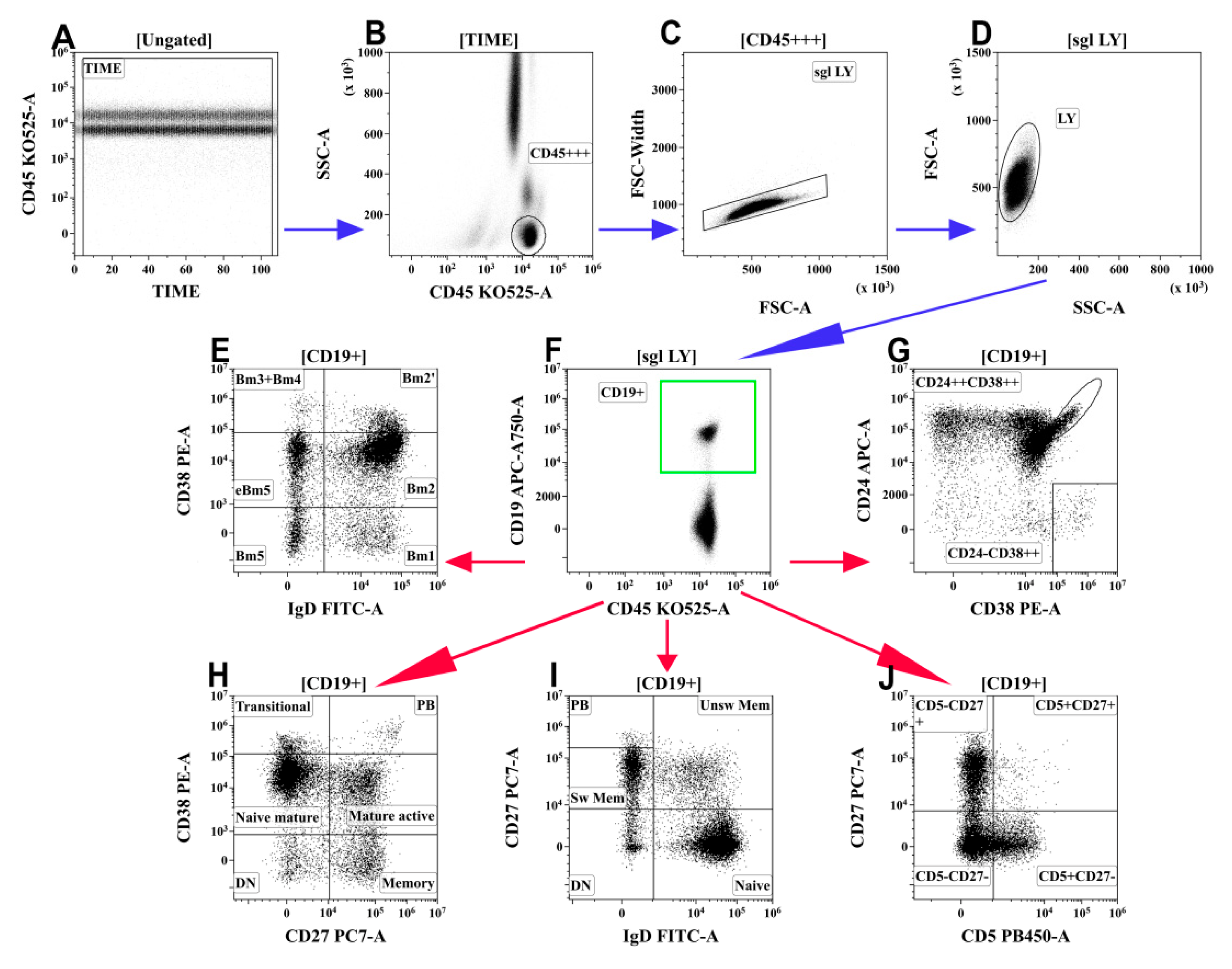
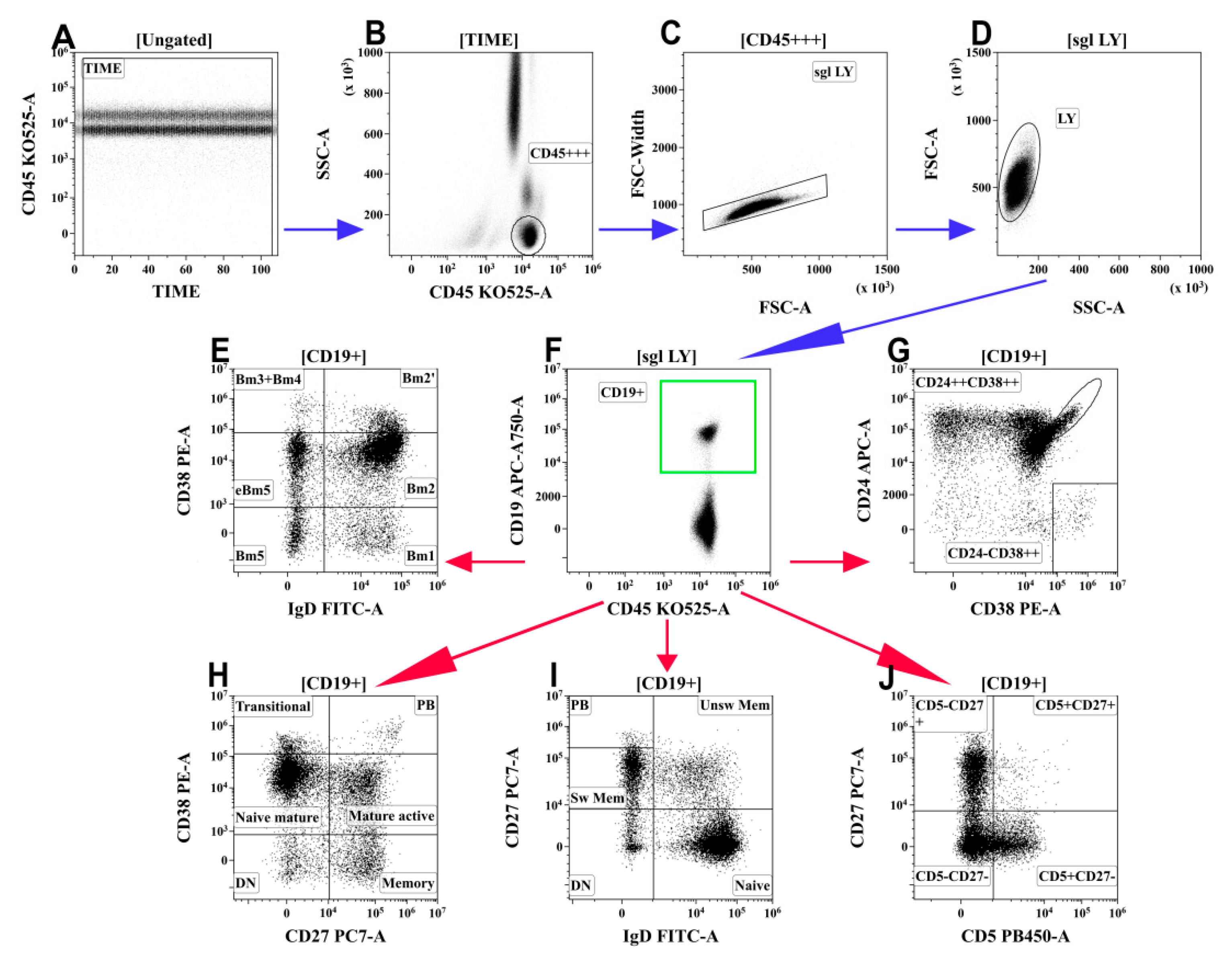
2.4. B-Cell Immunophenotyping by Flow Cytometry
2.5. Statistical Analysis
3. Results
3.1. Routine Laboratory Tests
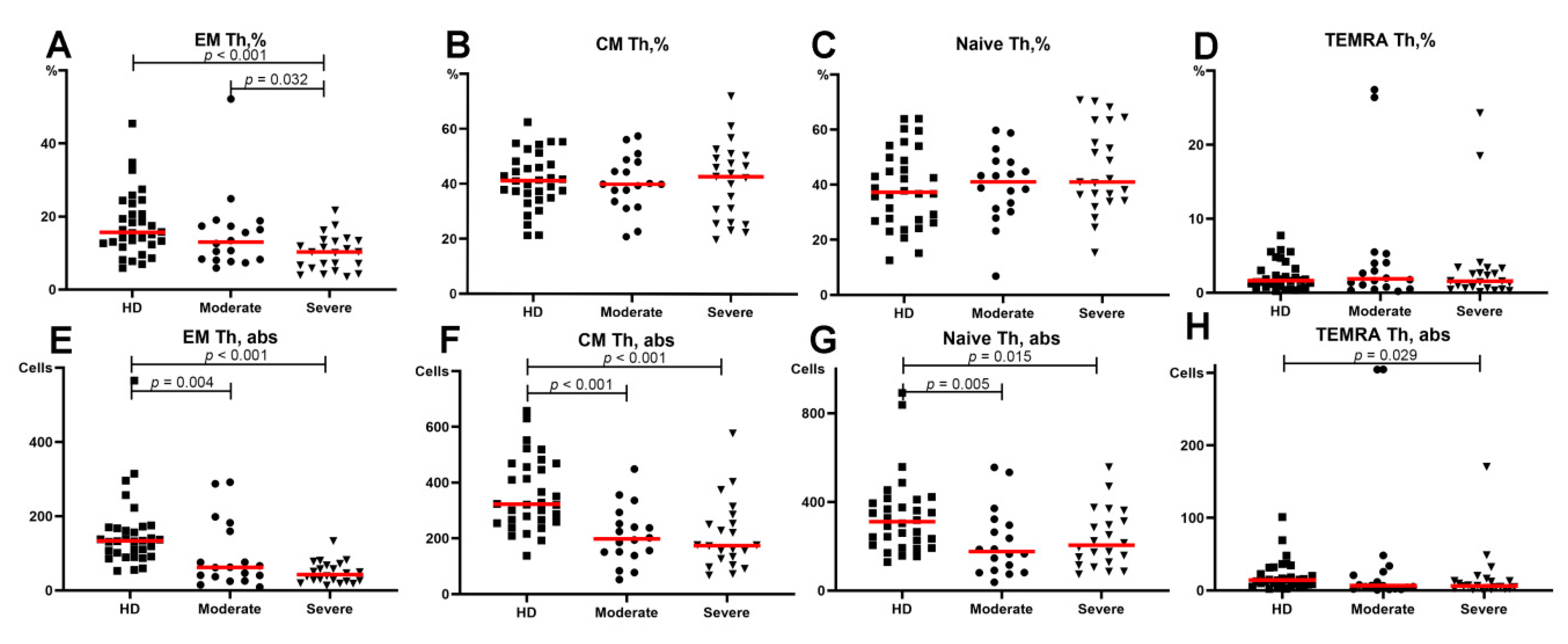
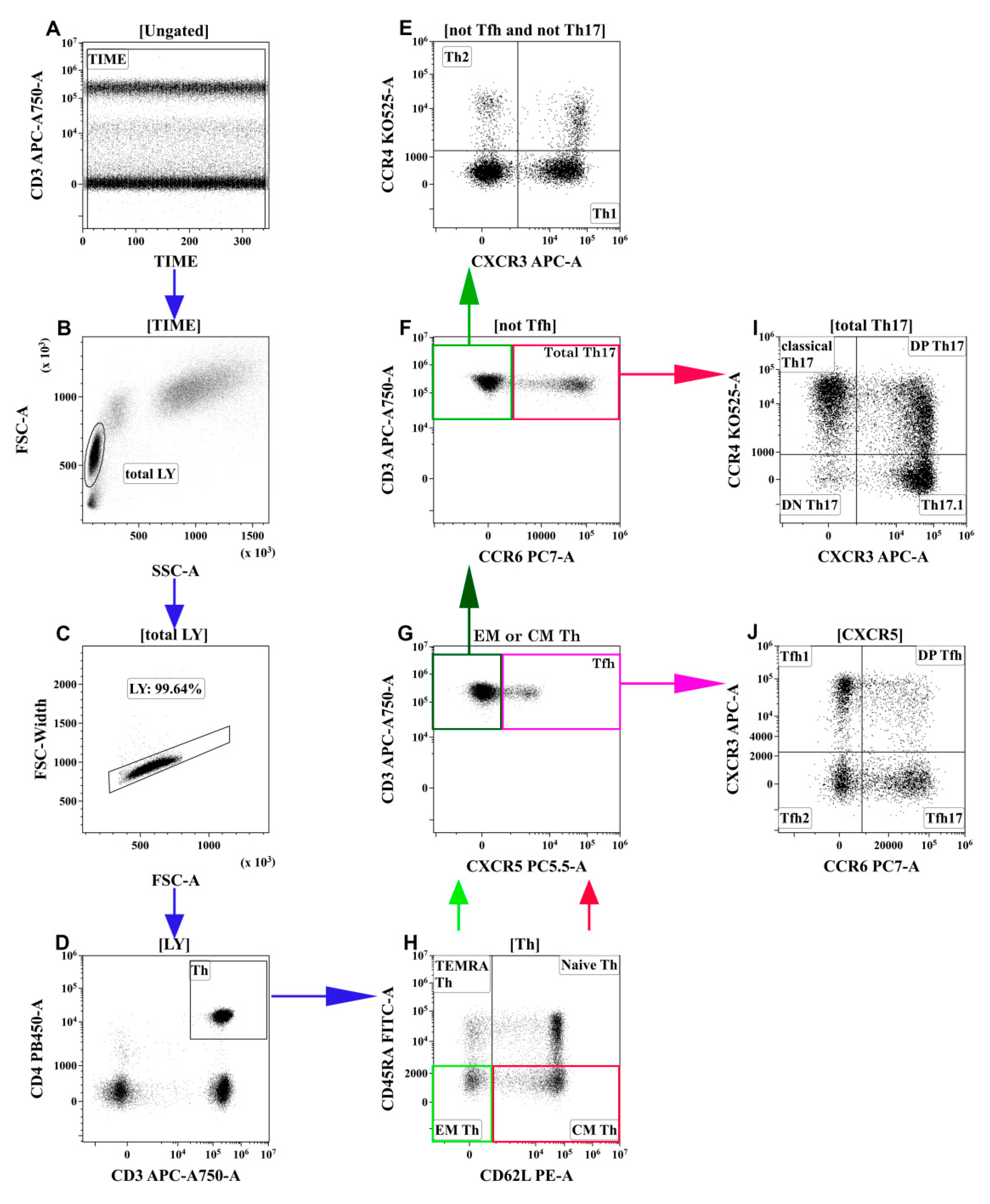
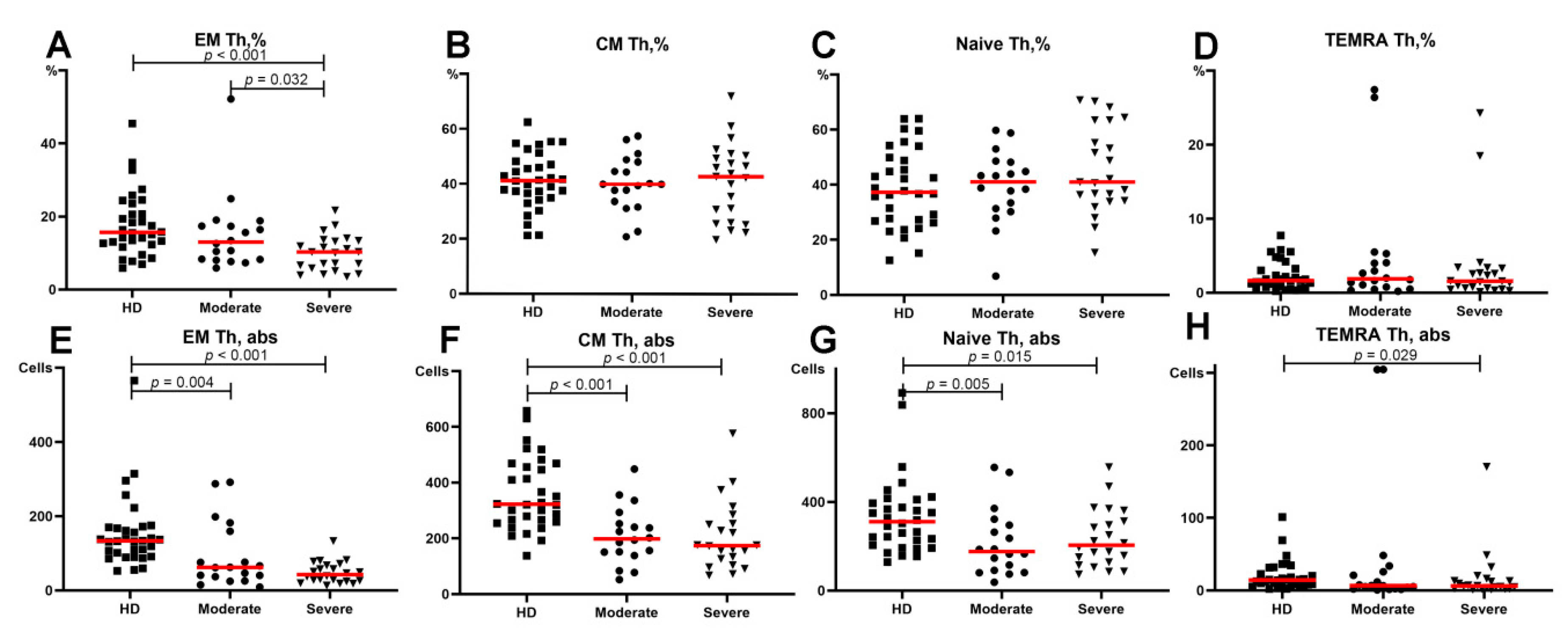
3.2. Alterations in the Main T-Cell Subsets of Patients with COVID-19
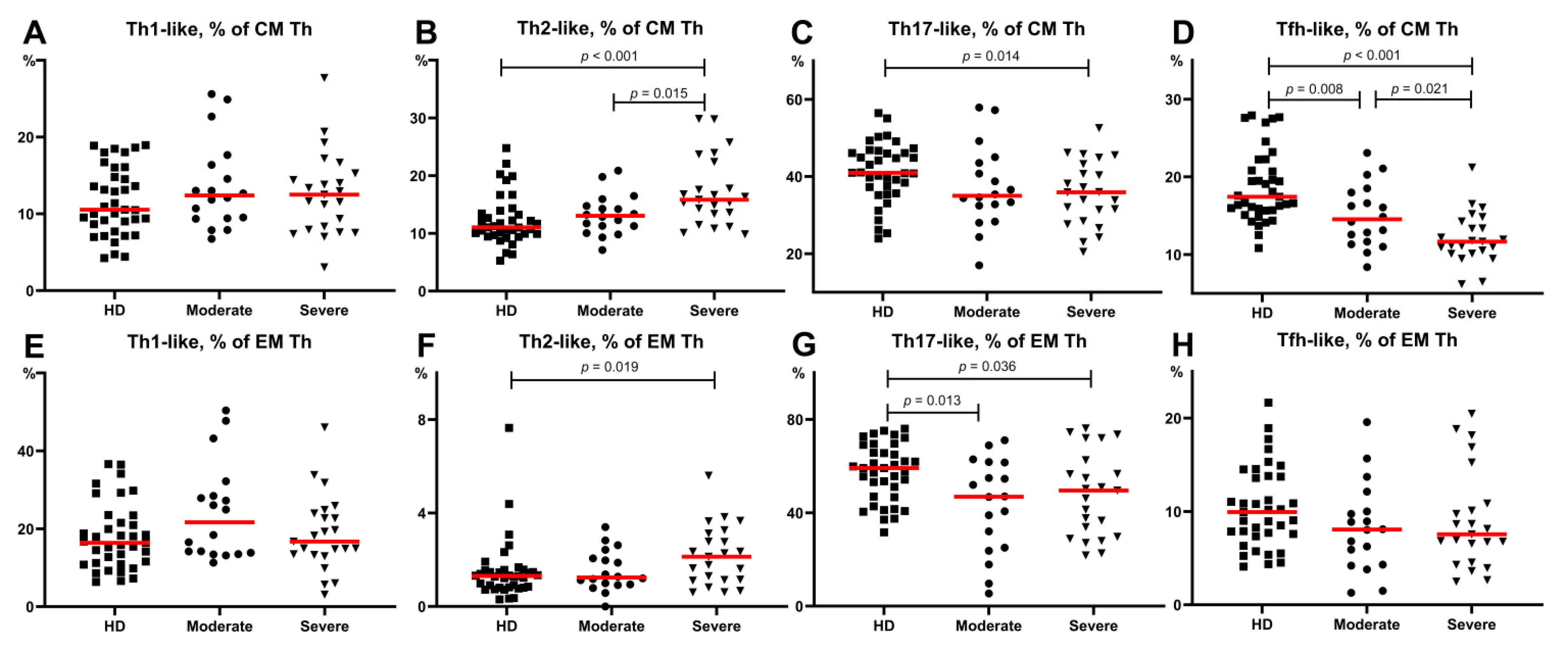
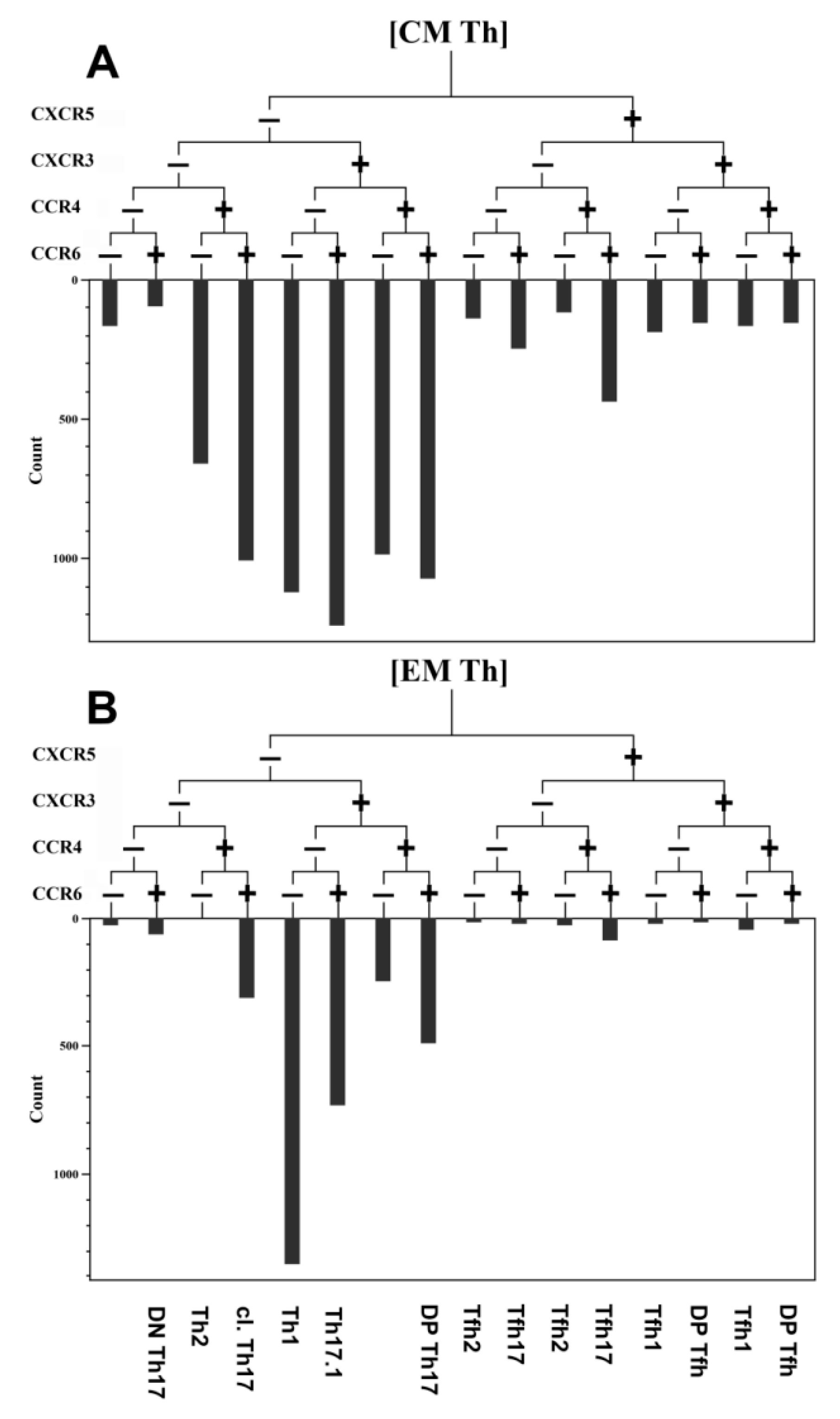
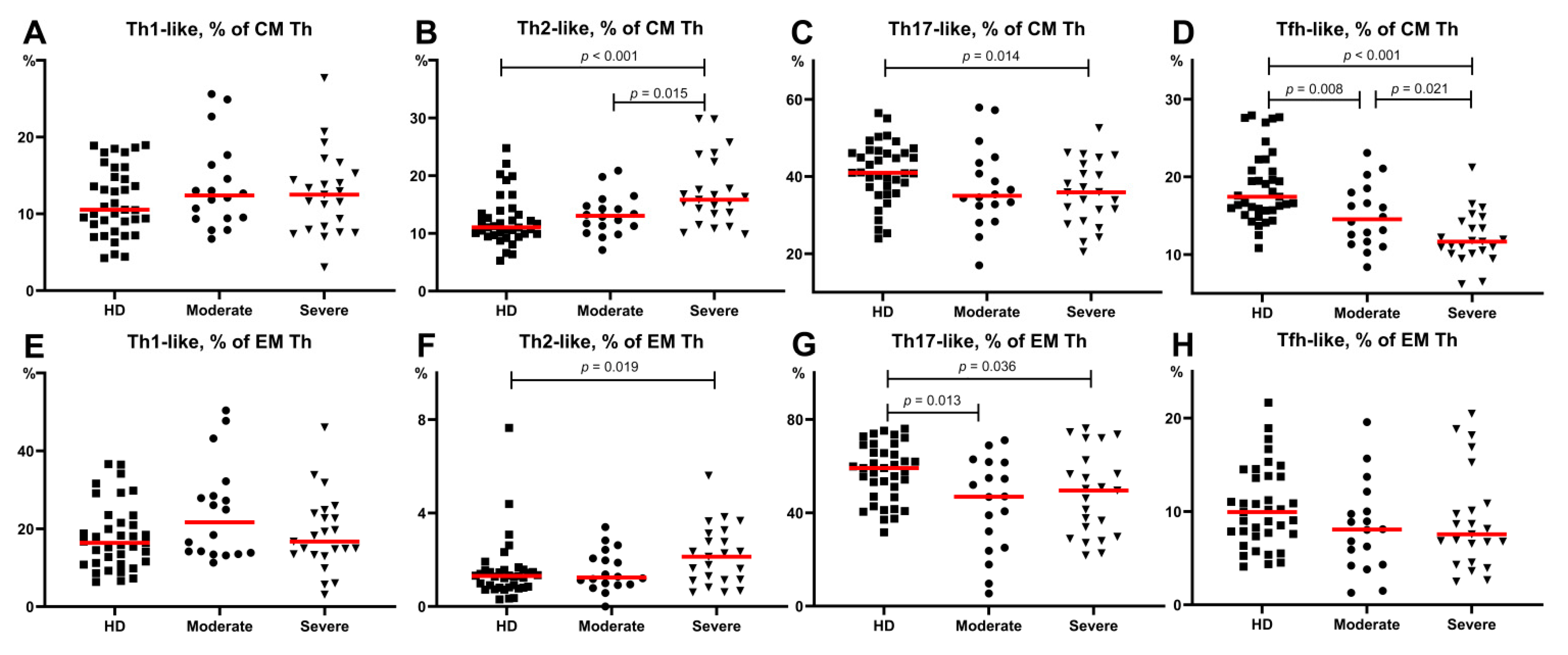
3.3. Imbalance of Main Polarized Th Cell Subsets in Patients with COVID-19
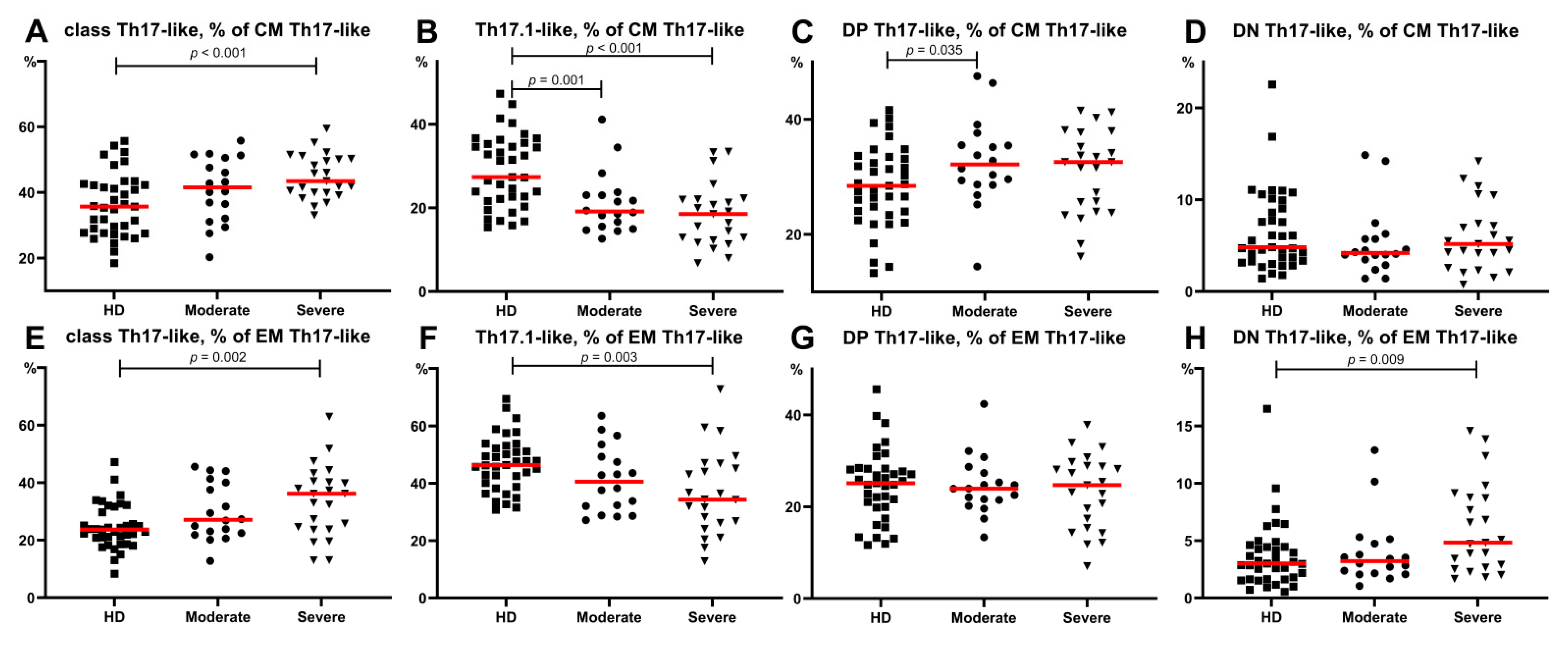
3.4. Imbalance of Th17-Like Subsets in Patients with COVID-19
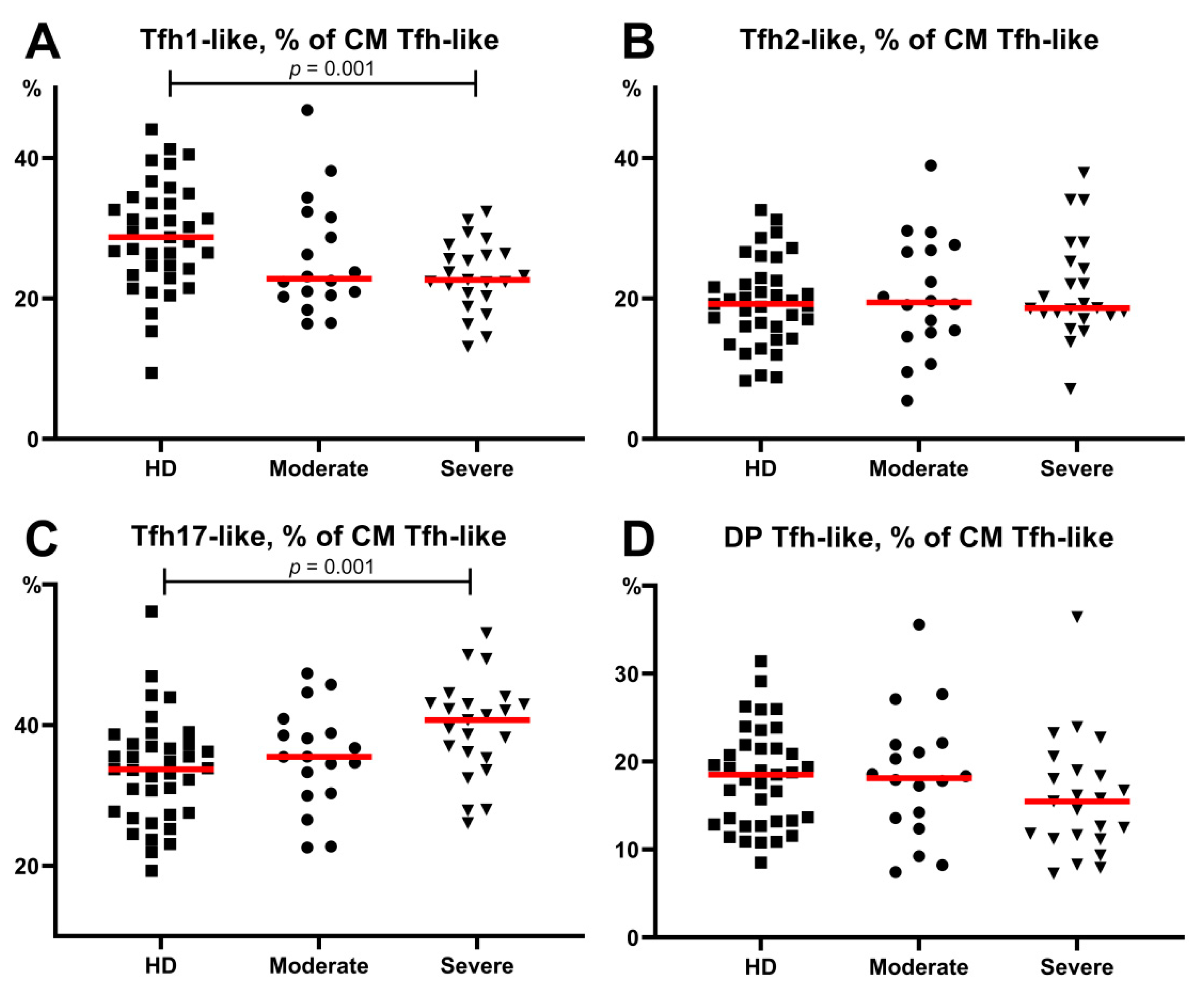
3.5. Alterations in Tfh-Like Subsets in Patients with COVID-19
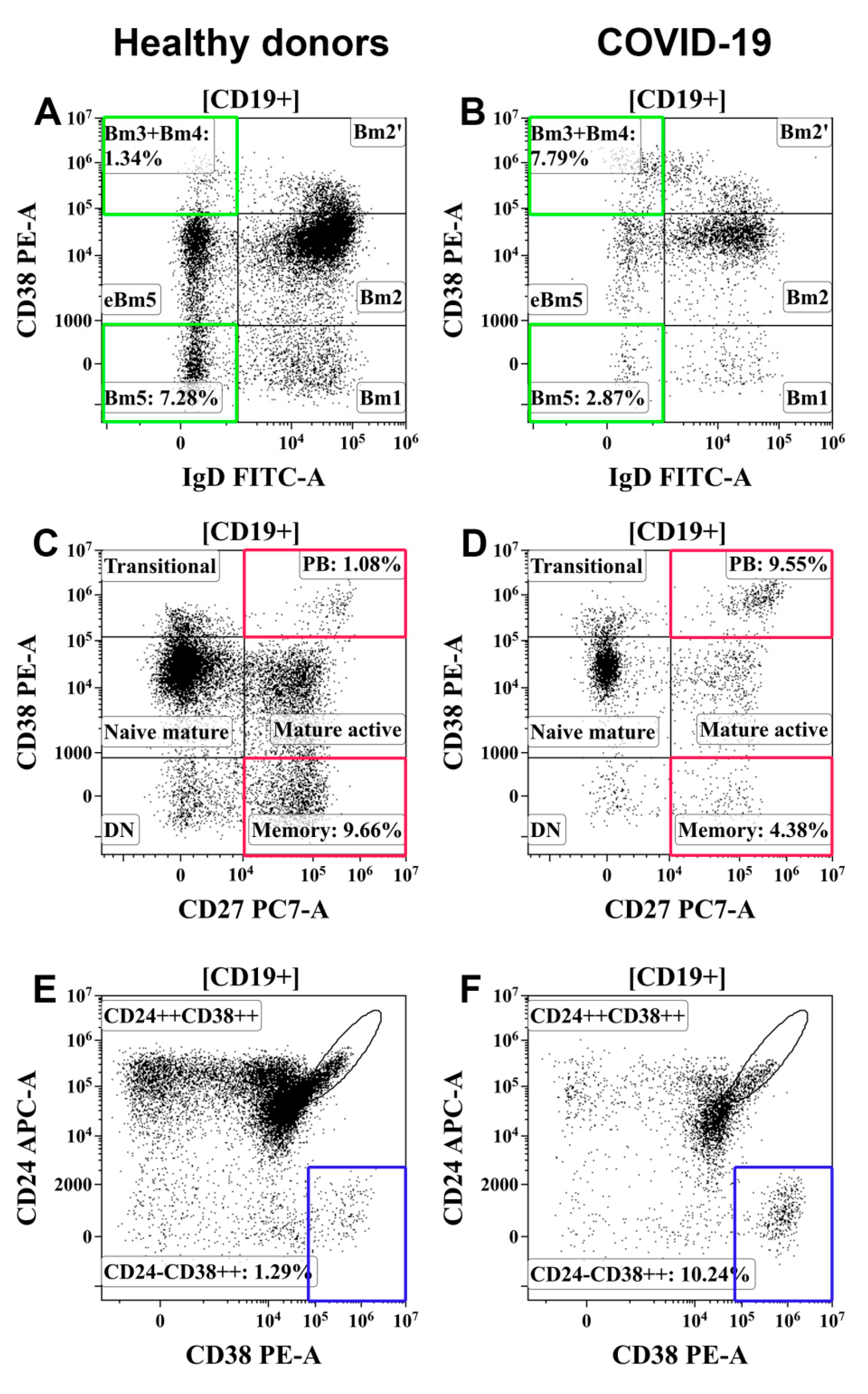
3.6. Alterations in Peripheral Blood B-Cell Subsets from Patients with COVID-19
4. Discussion
5. Conclusions
- Th2-like cells were predominant within central and effector memory Th cells in patients with severe COVID-19.
- The relative number of Th17-like cells was decreased within central and effector memory Th cells in patients with severe COVID-19.
- “Classical” Th17-like cells represented the most predominant Th17-like subset in patients with severe COVID-19, whereas the levels of “non-classical” Th17-like or Th17.1-like cells were dramatically decreased.
- The frequencies of circulating follicular Th-like cells were decreased within the central memory Th cells in patients with moderate and severe COVID-19, and Tfh17-like cells represented the most predominant Tfh-like subset in patients with severe COVID-19.
- Memory B cells were decreased and circulating plasma cell precursors were increased in the peripheral blood of patients with moderate and severe COVID-19.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fu, Y.; Cheng, Y.; Wu, Y. Understanding SARS-CoV-2-mediated inflammatory responses: From mechanisms to potential therapeutic tools. Virol. Sin. 2020, 35, 266–271. [Google Scholar] [CrossRef] [Green Version]
- Yang, M. Cell Pyroptosis, a Potential Pathogenic Mechanism of 2019-nCoV Infection (29 January 2020). Available online: https://ssrn.com/abstract=3527420 (accessed on 16 July 2021).
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of immune response in patients with COVID-19 in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; et al. Complex Immune Dysregulation in COVID-19 Patients with Severe Respiratory Failure. Cell Host Microbe 2020, 27, 992–1000. [Google Scholar] [CrossRef]
- Malkova, A.; Kudlay, D.; Kudryavtsev, I.; Starshinova, A.; Yablonskiy, P.; Shoenfeld, Y. Immunogenetic predictors of severe covid-19. Vaccines 2021, 9, 211. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, Y.D.R.M. Optimizing a Multicolor Immunophenotyping Assay. Clin. Lab. Med. 2007, 27, 469–485. [Google Scholar] [CrossRef] [Green Version]
- Nurkhametova, D.; Kudryavtsev, I.; Khayrutdinova, O.; Serebryakova, M.; Altunbaev, R.; Malm, T.; Giniatullin, R. Purinergic Profiling of Regulatory T-cells in Patients with Episodic Migraine. Front. Cell. Neurosci. 2018, 12, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallusto, F.; Lenig, D.; Förster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef]
- Barbarash, L.; Kudryavtsev, I.; Rutkovskaya, N.; Golovkin, A.T. Cell Response in Patients with Implanted Biological and Mechanical Prosthetic Heart Valves. Mediat. Inflamm. 2016, 2016, 1937564. [Google Scholar] [CrossRef]
- Kudryavtsev, I.; Serebriakova, M.; Zhiduleva, E.; Murtazalieva, P.; Titov, V.; Malashicheva, A.; Shishkova, A.; Semenova, D.; Irtyuga, O.; Isakov, D.; et al. CD73 Rather Than CD39 Is Mainly Involved in Controlling Purinergic Signaling in Calcified Aortic Valve Disease. Front. Genet. 2019, 10, 604. [Google Scholar] [CrossRef] [PubMed]
- Maecker, H.T.; McCoy, J.P.; Nussenblatt, R. Standardizing immunophenotyping for the Human Immunology Project. Nat. Rev. Immunol. 2012, 12, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finak, G.; Langweiler, M.; Jaimes, M.; Malek, M.; Taghiyar, J.; Korin, Y.; Raddassi, K.; Devine, L.; Obermoser, G.; Pekalski, M.; et al. Standardizing Flow Cytometry Immunophenotyping Analysis from the Human ImmunoPhenotyping Consortium. Sci. Rep. 2016, 6, 20686. [Google Scholar] [CrossRef] [Green Version]
- Brodie, T.; Brenna, E.; Sallusto, F. OMIP- 018, Chemokine receptor expression on human T helper cells. Cytom. Part. A 2013, 83, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [Green Version]
- Geginat, J.; Paroni, M.; Maglie, S.; Alfen, J.S.; Kastirr, I.; Gruarin, P.; De Simone, M.; Pagani, M.; Abrignani, S. Plasticity of human CD4 T cell subsets. Front. Immunol. 2014, 5, 630. [Google Scholar] [CrossRef]
- Caza, T.; Landas, S. Functional and Phenotypic Plasticity of CD4(+) T Cell Subsets. Biomed. Res. Int. 2015, 2015, 521957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosmi, L.; Maggi, L.; Santarlasci, V.; Liotta, F.; Annunziato, F. T helper cells plasticity in inflammation. Cytom. Part. A 2014, 85, 36–42. [Google Scholar] [CrossRef]
- Lord, G.M.; Rao, R.M.; Choe, H.; Sullivan, B.M.; Lichtman, A.H.; Luscinskas, F.W.; Glimcher, L.H. T-bet is required for optimal proinflammatory CD4+ T-cell trafficking. Blood 2005, 106, 3432–3439. [Google Scholar] [CrossRef] [PubMed]
- Sundrud, M.S.; Grill, S.M.; Ni, D.; Nagata, K.; Alkan, S.S.; Subramaniam, A.; Unutmaz, D. Genetic reprogramming of primary human T cells reveals functional plasticity in Th cell differentiation. J. Immunol. 2003, 171, 3542–3549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K.; Yoshitomi, H.; Hashimoto, M.; Maeda, S.; Teradaira, S.; Sugimoto, N.; Yamaguchi, T.; Nomura, T.; Ito, H.; Nakamura, T.; et al. Preferential recruitment of CCR6-expressing Th17 cells to inflamed joints via CCL20 in rheumatoid arthritis and its animal model. J. Exp. Med. 2007, 204, 2803–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitfeld, D.; Ohl, L.; Kremmer, E.; Ellwart, J.; Sallusto, F.; Lipp, M.; Förster, R. Follicular B helper T cells express CXC chemokine receptor 5, localize to B cell follicles, and support immunoglobulin production. J. Exp. Med. 2000, 192, 1545–1552. [Google Scholar] [CrossRef]
- Morita, R.; Schmitt, N.; Bentebibel, S.E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human Blood CXCR5+CD4+ T Cells Are Counterparts of T Follicular Cells and Contain Specific Subsets that Differentially Support Antibody Secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S.T. Follicular Helper Cell Biology: A Decade of Discovery and Diseases. Immunity 2019, 50, 1132–1148. [Google Scholar] [CrossRef]
- Bohnhorst, J.Ø.; Bjørgan, M.B.; Thoen, J.E.; Natvig, J.B.; Thompson, K.M. Bm1–Bm5 Classification of Peripheral Blood B Cells Reveals Circulating Germinal Center Founder Cells in Healthy Individuals and Disturbance in the B Cell Subpopulations in Patients with Primary Sjögren’s Syndrome. J. Immunol. 2001, 167, 3610–3618. [Google Scholar] [CrossRef]
- Sanz, I.; Wei, C.; Lee, F.E.H.; Anolik, J. Phenotypic and functional heterogeneity of human memory B cells. Semin. Immunol. 2008, 20, 67–82. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Cai, X.; Wang, H.; He, G.; Lin, Y.; Lu, B.; Chen, C.; Pan, Y.; Hu, X. Abnormalities of peripheral blood system in patients with COVID-19 in Wenzhou, China. Clin. Chim. Acta 2020, 507, 174–180. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Version, T. Diagnosis and Treatment Plan for COVID-19 (Trial Version 6). Chin. Med. J. 2020, 133, 1087–1095. [Google Scholar]
- Wei, L.-L.; Wang, W.-J.; Chen, D.-X.; Xu, B. Dysregulation of the immune response affects the outcome of critical COVID-19 patients. J. Med. Virol. 2020, 92, 2768–2776. [Google Scholar] [CrossRef] [PubMed]
- Mahnke, Y.D.; Brodie, T.M.; Sallusto, F.; Roederer, M.L.E. The who’s who of T-cell differentiation: Human memory T-cell subsets. Eur. J. Immunol. 2013, 43, 2797–2809. [Google Scholar] [CrossRef]
- Sallusto, F. Heterogeneity of Human CD4+ T Cells Against Microbes. Annu. Rev. Immunol. 2016, 34, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Mathew, D.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Greenplate, A.R.; Wu, J.E.; Alanio, C.; Kuri-Cervantes, L.; Pampena, M.; D’Andrea, K.; et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science 2020, 369, eabc8511. [Google Scholar] [CrossRef] [PubMed]
- de Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Gozzi, L.; Iannone, A.; Lo Tartaro, D.; Mattioli, M.; et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11, 3434. [Google Scholar] [CrossRef]
- Gutiérrez-Bautista, J.F.; Rodriguez-Nicolas, A.; Rosales-Castillo, A.; Jiménez, P.; Garrido, F.; Anderson, P.; Ruiz-Cabello, F.; López-Ruz, M. Negative Clinical Evolution in COVID-19 Patients Is Frequently Accompanied with an Increased Proportion of Undifferentiated Th Cells and a Strong Underrepresentation of the Th1 Subset. Front. Immunol. 2020, 11, 596553. [Google Scholar] [CrossRef]
- Spoerl, S.; Kremer, A.N.; Aigner, M.; Eisenhauer, N.; Koch, P.; Meretuk, L.; Löffler, P.; Tenbusch, M.; Maier, C.; Überla, K.; et al. Upregulation of CCR4 in activated CD8+ T cells indicates enhanced lung homing in patients with severe acute SARS-CoV-2 infection. Eur. J. Immunol. 2021, 51, 1436–1448. [Google Scholar] [CrossRef]
- Saris, A.; Reijnders, T.D.Y.; Nossent, E.J.; Schuurman, A.R.; Verhoeff, J.; Van Asten, S.; Bontkes, H.; Blok, S.; Duitman, J.; Bogaard, H.; et al. Distinct cellular immune profiles in the airways and blood of critically ill patients with COVID-19. Thorax 2021, 76, 1010–1019. [Google Scholar] [CrossRef]
- Zhou, Y.; Fu, B.; Zheng, X.; Wang, D.; Zhao, C.; Qi, Y.; Sun, R.; Tian, Z.; Xu, X.; Wei, H. Aberrant pathogenic GM-CSF+ T cells and inflammatory CD14+ CD16+ monocytes in severe pulmonary syndrome patients of a new coronavirus. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunologic features in severe and moderate Coronavirus Disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Fu, B.; Zheng, X.; Wang, D.; Zhao, C.; Qi, Y.; Sun, R.; Tian, Z.; Xu, X.; Wei, H. Pathogenic T-cells and inflammatory monocytes incite inflammatory storms in severe COVID-19 patients. Natl. Sci. Rev. 2020, 7, 998–1002. [Google Scholar] [CrossRef] [Green Version]
- Abdoli, A.; Ardakani, H.M. Helminth infections and immunosenescence: The friend of my enemy. Exp. Gerontol. 2020, 133, 110852. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Edwards, J.E. Type 1/type 2 immunity in infectious diseases. Clin. Infect. Dis. 2001, 32, 76–102. [Google Scholar] [CrossRef] [PubMed]
- Li, C.K.; Wu, H.; Yan, H.; Ma, S.; Wang, L.; Zhang, M.; Tang, X.; Temperton, N.; Weiss, R.; Brenchley, J.; et al. T Cell Responses to Whole SARS Coronavirus in Humans. J. Immunol. 2008, 181, 5490–5500. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.J.; Busse, W.W.; Bacharier, L.B.; Kattan, M.; O’Connor, G.T.; Wood, R.A.; Visness, C.; Durham, S.; Larson, D.; Esnault, S.; et al. Association of respiratory allergy, asthma, and expression of the SARS-CoV-2 receptor ACE2. J. Allergy Clin. Immunol. 2020, 146, 203–206.e3. [Google Scholar] [CrossRef]
- Shu, C.C.; Wu, M.F.; Wang, J.Y.; Lai, H.C.; Lee, L.N.; Chiang, B.L.; Yu, C.J. Decreased T helper 17 cells in tuberculosis is associated with increased percentages of programmed death ligand 1, T helper 2 and regulatory T cells. Respir. Res. 2017, 18, 128. [Google Scholar] [CrossRef] [Green Version]
- Kudryavtsev, I.V.; Serebriakova, M.K.; Starshinova, A.A.; Zinchenko, Y.S.; Basantsova, N.Y.; Belyaeva, E.N.; Pavlova, M.V.; Yablonskiy, P.K. Altered peripheral blood Th17 and follicular T-helper subsets in patients with pulmonary tuberculosis. Russ. J. Infect. Immun. 2019, 9, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, R.K.; Al Heialy, S.; Hamid, Q. Implications of preexisting asthma on COVID-19 pathogenesis. Am. J. Physiol. -Lung Cell. Mol. Physiol. 2021, 320, L880–L891. [Google Scholar] [CrossRef]
- Gil-Etayo, F.J.; Suàrez-Fernández, P.; Cabrera-Marante, O.; Arroyo, D.; Garcinuño, S.; Naranjo, L.; Pleguezuelo, D.; Allende, L.; Mancebo, E.; Lalueza, A.; et al. T-Helper Cell Subset Response Is a Determining Factor in COVID-19 Progression. Front. Cell. Infect. Microbiol. 2021, 11, 79. [Google Scholar] [CrossRef]
- Roncati, L.; Nasillo, V.; Lusenti, B.; Riva, G. Signals of Th2 immune response from COVID-19 patients requiring intensive care. Ann. Hematol. 2020, 99, 1419–1420. [Google Scholar] [CrossRef]
- Roncati, L.; Ligabue, G.; Fabbiani, L.; Malagoli, C.; Gallo, G.; Lusenti, B.; Nasillo, V.; Manenti, A.; Maiorana, A. Type 3 hypersensitivity in COVID-19 vasculitis. Clin. Immunol. 2020, 217, 108487. [Google Scholar] [CrossRef]
- Schultheiß, C.; Paschold, L.; Simnica, D.; Mohme, M.; Willscher, E.; von Wenserski, L.; Scholz, R.; Wieters, I.; Dahlke, C.; Tolosa, E.; et al. Next-Generation Sequencing of T and B Cell Receptor Repertoires from COVID-19 Patients Showed Signatures Associated with Severity of Disease. Immunity 2020, 53, 442–455. [Google Scholar] [CrossRef]
- Mahmoud Salehi Khesht, A.; Karpisheh, V.; Qubais Saeed, B.; Olegovna Zekiy, A.; Yapanto, L.M.; Nabi Afjadi, M.; Aksoun, M.; Nasr Esfahani, M.; Aghakhani, F.; Movahed, M.; et al. Different T cell related immunological profiles in COVID-19 patients compared to healthy controls. Int. Immunopharmacol. 2021, 97, 107828. [Google Scholar] [CrossRef]
- Shibabaw, T. Inflammatory cytokine: Il-17a signaling pathway in patients present with covid-19 and current treatment strategy. J. Inflamm. Res. 2020, 13, 673. [Google Scholar] [CrossRef] [PubMed]
- Orlov, M.; Wander, P.L.; Morrell, E.D.; Mikacenic, C.; Wurfel, M.M. A Case for Targeting Th17 Cells and IL-17A in SARS-CoV-2 Infections. J. Immunol. 2020, 205, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Osaga, S.; Maeda, T.; Takeda, N.; Tamechika, S.Y.; Naniwa, T.; Niimi, A. Circulating Th17.1 cells as candidate for the prediction of therapeutic response to abatacept in patients with rheumatoid arthritis: An. exploratory research. PLoS ONE 2019, 14, e0215192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacleche, V.S.; Goulet, J.P.; Gosselin, A.; Monteiro, P.; Soudeyns, H.; Fromentin, R.; Jenabian, M.; Vartanian, S.; Deeks, S.; Chomont, N.; et al. New insights into the heterogeneity of Th17 subsets contributing to HIV-1 persistence during antiretroviral therapy. Retrovirology 2016, 13, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, M.; Maggi, L.; Micheletti, A.; Lazzeri, E.; Tamassia, N.; Costantini, C.; Cosmi, L.; Lunardi, C.; Annunziato, F.; Romagnani, S.; et al. Evidence for a cross-talk between human neutrophils and Th17 cells. Blood J. Am. Soc. Hematol. 2010, 115, 335–343. [Google Scholar] [CrossRef]
- Kaneko, N.; Kuo, H.-H.H.; Boucau, J.; Farmer, J.R.; Allard-Chamard, H.; Mahajan, V.S.; Piechocka-Trocha, A.; Lefteri, K.; Osborn, M.; Bals, J.; et al. Loss of Bcl-6-Expressing T Follicular Helper Cells and Germinal Centers in COVID-19. Cell 2020, 183, 143–157.e13. [Google Scholar] [CrossRef]
- Gong, F.; Dai, Y.; Zheng, T.; Cheng, L.; Zhao, D.; Wang, H.; Liu, M.; Pei, H.; Jin, T.; Yu, D.; et al. Peripheral CD4+ T cell subsets and antibody response in COVID-19 convalescent individuals. J. Clin. Investig. 2020, 130, 6588–6599. Available online: https://www.jci.org/articles/view/141054 (accessed on 16 July 2021). [CrossRef]
- Kurata, I.; Matsumoto, I.; Sumida, T. T follicular helper cell subsets: A potential key player in autoimmunity. Immunol. Med. 2021, 44, 1–9. [Google Scholar] [CrossRef]
- Schmitt, N.; Bentebibel, S.E.; Ueno, H. Phenotype and functions of memory Tfh cells in human blood. Trends Immunol. 2014, 35, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolla, R.; Vidali, M.; Puricelli, C.; Scotta, A.M.; Pedrinelli, A.; Pergolini, P.; Pirisi, M.; Dianzani, U.; Rigamonti, C. Reduced activity of B-lymphocytes, recognised by Sysmex XN-2000TM haematology analyser, predicts mortality in patients with Coronavirus disease 2019. Int. J. Lab. Hematol. 2021, 43, e5–e8. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, Y.; Liu, H.; Jia, Y.; Li, F.; Wang, W.; Wu, J.; Wan, Z.; Cao, Y.; Zeng, R. Immune dysfunction leads to mortality and organ injury in patients with COVID-19 in China: Insights from ERS-COVID-19 study. Signal. Transduct. Target. Ther. 2020, 5, 62. [Google Scholar] [CrossRef] [PubMed]
- Pence, B.D. Severe COVID-19 and aging: Are monocytes the key? GeroScience 2020, 42, 1051–1061. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comorbidities | Total Number of Patients, n (%) | Moderate, n (% Inside Group) | Severe, n (% Inside Group) |
|---|---|---|---|
| Arterial hypertension | 19 (46) | 7 (39) | 12 (52) |
| Diabetes mellitus | 5 (12) | 2 (11) | 3 (13) |
| Ischemic heart disease | 9 (22) | 4 (22) | 5 (22) |
| Cancer in anamnesis | 3 (7) | 3 (17) | 0 |
| Bronchial asthma | 5 (12) | 2 (11) | 3 (13) |
| Parameter, Units (Reference Value) | Healthy Donors | Patients with COVID-19 | |
|---|---|---|---|
| Moderate (n = 18) | Severe (n = 23) | ||
| Age | 37.0 (32.0;47.0) | 62.5 (52.0;70.0) ^ | 59.0 (53.0;77.0) ^ |
| CT, % lung damage | 40 (32;52) * | 60 (48;68) * | |
| CRP, mg/L (0–5.0) | 64.9 (39.7;80.1) | 71.4 (36.1;109.5) | |
| D-dimer, µg/mL (0–5.0) | 0.55 (0.36;1.25) | 0.62 (0.40;1.48) | |
| Ferritin, ng/mL (30–400) | 372.8 (148.4;901.1) | 422.6 (164.5;713.4) | |
| Fibrinogen, g/L (1.9–4.3) | 6.0 (4.1;6.4) | 6.0 (4.7;7.0) | |
| Troponin I, ng/mL (0–0.034) | 0.006 (0.002;0.014) | 0.002 (0.002;0.002) | |
| Procalcitonin, ng/mL (0–0.5) | 0.07 (0.04;0.15) | 0.06 (0.05;0.18) | |
| ALT, U/L (0–41) | 39.1 (25.2;72.1) | 29.0 (23.0;63.5) | |
| AST, U/L (0–40) | 46.0 (31.0;80.0) | 37.8 (32.0;61.0) | |
| LDH, U/L (135–225) | 335 (325;349) | 332 (267;410) | |
| Haemoglobin, g/L | 139.5 (136.0;148.0) | 128.6 (123.0;140.3) | 141.2 (127.2;148.8) |
| RBC, ×1012 | 5.00 (4.70;5.13) | 4.71 (4.07;5.02) | 4.71 (4.45;4.91) |
| Ht, % | 43.2 (41.5;46.0) * | 38.6 (34.9;42.4) ^ | 40.2 (36.8;42.2) ^ |
| Plt, ×109 | 208 (194;235) | 174 (153;212) | 195 (130;301) |
| WBC, ×109 | 5.8 (5.4;7.7) | 5.1 (3.2;7.3) | 4.4 (3.5;6.1) ^ |
| Lymphocytes, ×109 | 1.95 (1.78;2.21) * | 1.03 (0.62;1.30) ^ | 1.02 (0.75;1.26) ^ |
| Monocytes, ×109 | 0.58 (0.39;0.71) | 0.51 (0.29;0.58) | 0.38 (0.24;0.51) ^ |
| Neutrophils, ×109 | 2.96 (2.78;4.31) | 3.53 (1.98;4.65) | 3.06 (2.31;3.97) |
| Eosinophils, ×109 | 0.11 (0.08;0.23) * | 0.02 (0.01;0.03) ^ | 0.01 (0;0.04) ^ |
| Basophils, ×109 | 0.08 (0.06;1.00) * | 0.03 (0.02;0.04) ^ | 0.01 (0.01;0.04) ^ |
| Parameter | Healthy Donors | COVID-19 Patients | |||
|---|---|---|---|---|---|
| # | % | Severity | # | % | |
| Bm1 | 22.2 (13.2;38.8) | 10.7 (7.9;13.8) | Moderate | 5.0 (3.4;7.7) * | 6.1 (4.2;8.8) * |
| Severe | 4.2 (2.0;8.6) * | 4.5 (1.6;6.4) * | |||
| Bm2 | 127.1 (91.8;184.2) | 58.4 (52.6;63.7) | Moderate | 40.0 (20.8;75.40) * | 53.6 (44.2;61.8) |
| Severe | 73.7 (35.0;111.1) * | 55.4 (48.3;68.9) | |||
| Bm2′ | 19.3 (11.2;25.1) | 8.2 (6.4;10.3) | Moderate | 8.2 (5.0;14.4) * | 10.8 (8.0;14.4) |
| Severe | 13.8 (7.3;21.8) | 11.2 (7.4;16.7) * | |||
| Bm3 + Bm4 | 2.6 (1.8;4.1) | 1.1 (0.8;2.1) | Moderate | 6.7 (3.8;13.3) * | 10.5 (6.9;12.3) * |
| Severe | 6.1 (2.9;12.0) * | 7.6 (3.0;15.2) * | |||
| eBm5 | 25.1 (16.4;37.4) | 11.3 (8.7;14.1) | Moderate | 9.3 (5.7;14.9) * | 11.3 (7.9;15.7) |
| Severe | 8.4 (5.9;15.4) * | 9.1 (5.6;12.1) | |||
| Bm5 | 15.9 (9.9;24.0) | 7.2 (5.1;10.6) | Moderate | 3.6 (2.3;4.9) * | 5.5 (3.8;10.0) * |
| Severe | 3.8 (1.8;6.0) * | 3.3 (1.8;7.1) * | |||
| Parameter | Healthy Donors | COVID-19 Patients | |||
|---|---|---|---|---|---|
| # | % | Severity | # | % | |
| Naïve, IgD+CD27– | 139.5 (89.6;205.5) | 65.4 (55.8;73.4) | Moderate | 49.3 (20.5;87.8) * | 56.1 (46.7;65.4) |
| Severe | 80.5 (35.1;127.7) * | 62.9 (49.2;83.3) | |||
| Unswitched memory, IgD+CD27+ | 27.8 (17.9;42.2) | 14.3 (9.8;17.9) | Moderate | 9.2 (4.4;15.1) * | 11.7 (7.1;19.9) |
| Severe | 7.8 (4.1;11.6) * | 5.6 (4.0;14.0) * | |||
| Class-switched memory, IgD–CD27+ | 35.5 (25.6;52.3) | 16.3 (11.5;20.4) | Moderate | 13.2 (7.5;21.9) * | 16.4 (11.9;23.3) ^ |
| Severe | 9.5 (5.8;22.8) * | 13.1 (5.6;16.6) *,^ | |||
| Double-negative memory, IgD–CD27− | 7.2 (5.6;12.6) | 3.9 (2.3;4.8) | Moderate | 4.1 (3.1;9.5) * | 5.8 (4.4;8.1) * |
| Severe | 3.9 (2.5;11.3) * | 4.3 (2.6;8.5) | |||
| Plasma cell precursors, IgD–CD27++ | 0.8 (0.3;3.3) | 0.4 (0.1;1.8) | Moderate | 3.3 (2.5;9.0) * | 5.9 (3.5;8.1) * |
| Severe | 4.1 (2.0;10.4) * | 4.9 (1.4;12.0) * | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golovkin, A.; Kalinina, O.; Bezrukikh, V.; Aquino, A.; Zaikova, E.; Karonova, T.; Melnik, O.; Vasilieva, E.; Kudryavtsev, I. Imbalanced Immune Response of T-Cell and B-Cell Subsets in Patients with Moderate and Severe COVID-19. Viruses 2021, 13, 1966. https://doi.org/10.3390/v13101966
Golovkin A, Kalinina O, Bezrukikh V, Aquino A, Zaikova E, Karonova T, Melnik O, Vasilieva E, Kudryavtsev I. Imbalanced Immune Response of T-Cell and B-Cell Subsets in Patients with Moderate and Severe COVID-19. Viruses. 2021; 13(10):1966. https://doi.org/10.3390/v13101966
Chicago/Turabian StyleGolovkin, Alexey, Olga Kalinina, Vadim Bezrukikh, Arthur Aquino, Ekaterina Zaikova, Tatyana Karonova, Olesya Melnik, Elena Vasilieva, and Igor Kudryavtsev. 2021. "Imbalanced Immune Response of T-Cell and B-Cell Subsets in Patients with Moderate and Severe COVID-19" Viruses 13, no. 10: 1966. https://doi.org/10.3390/v13101966
APA StyleGolovkin, A., Kalinina, O., Bezrukikh, V., Aquino, A., Zaikova, E., Karonova, T., Melnik, O., Vasilieva, E., & Kudryavtsev, I. (2021). Imbalanced Immune Response of T-Cell and B-Cell Subsets in Patients with Moderate and Severe COVID-19. Viruses, 13(10), 1966. https://doi.org/10.3390/v13101966