
Novel Virulent Bacteriophage ΦSG005, Which Infects Streptococcus gordonii, Forms a Distinct Clade among Streptococcus Viruses

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Bacteriophage Isolation
2.3. Bacteriophage Preparation
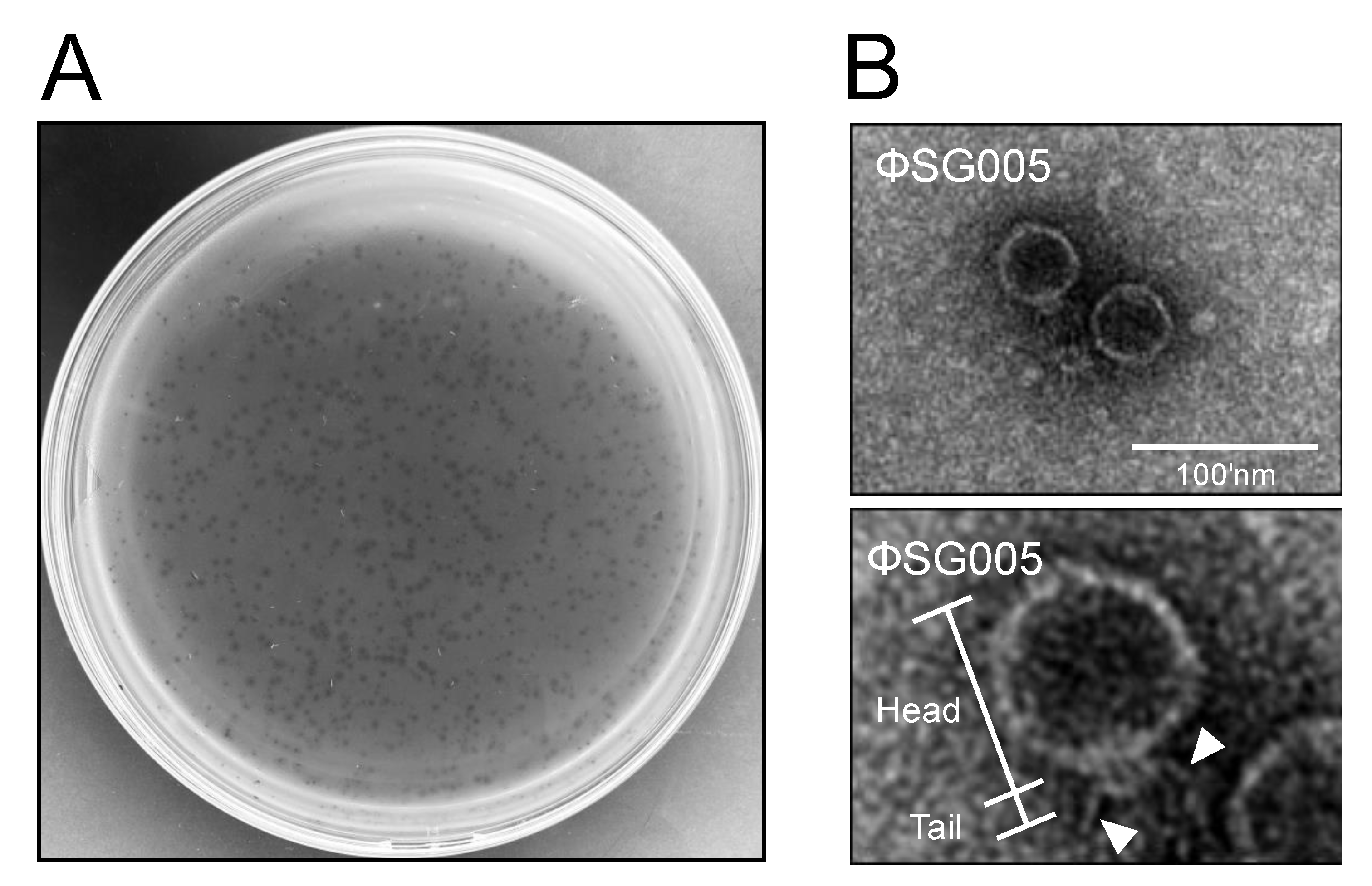
2.4. Transmission Electron Microscopic Imaging
2.5. Whole-Genome Sequencing
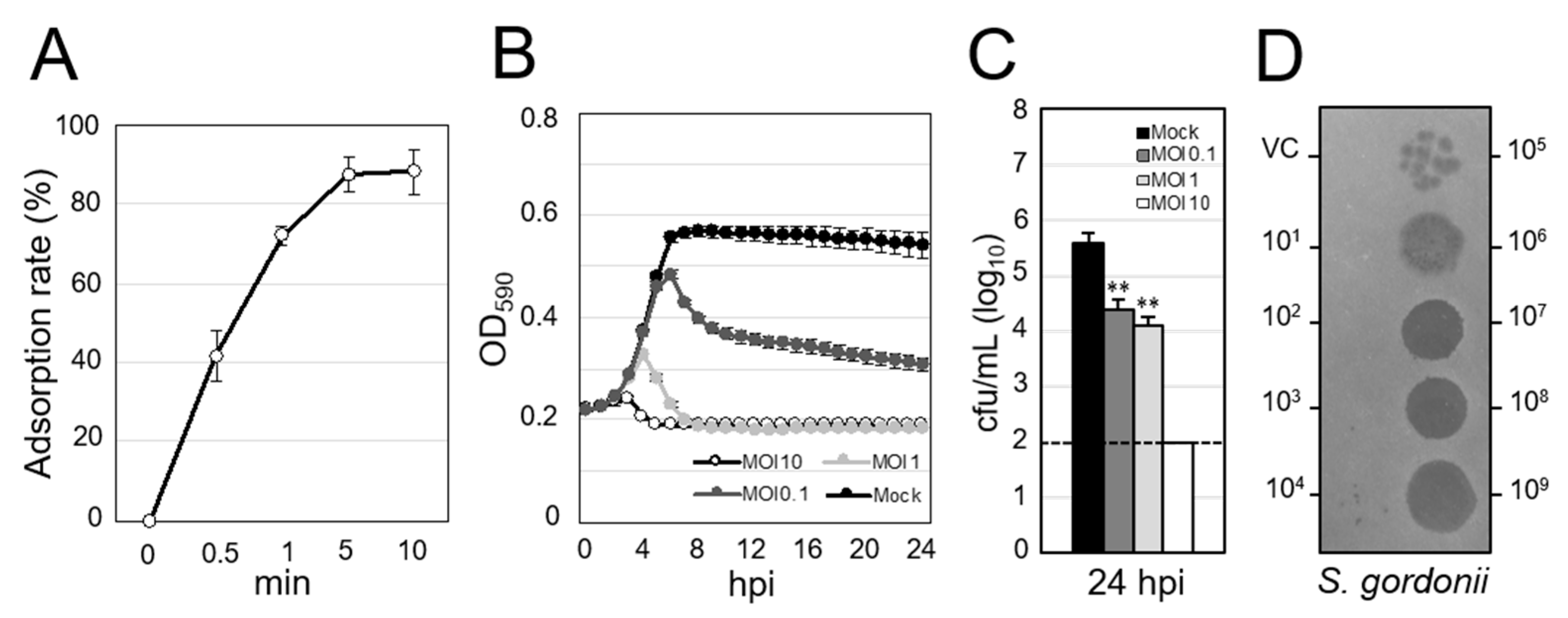
2.6. Adsorption Assay
2.7. Co-Culture of the Phage and Host Strain
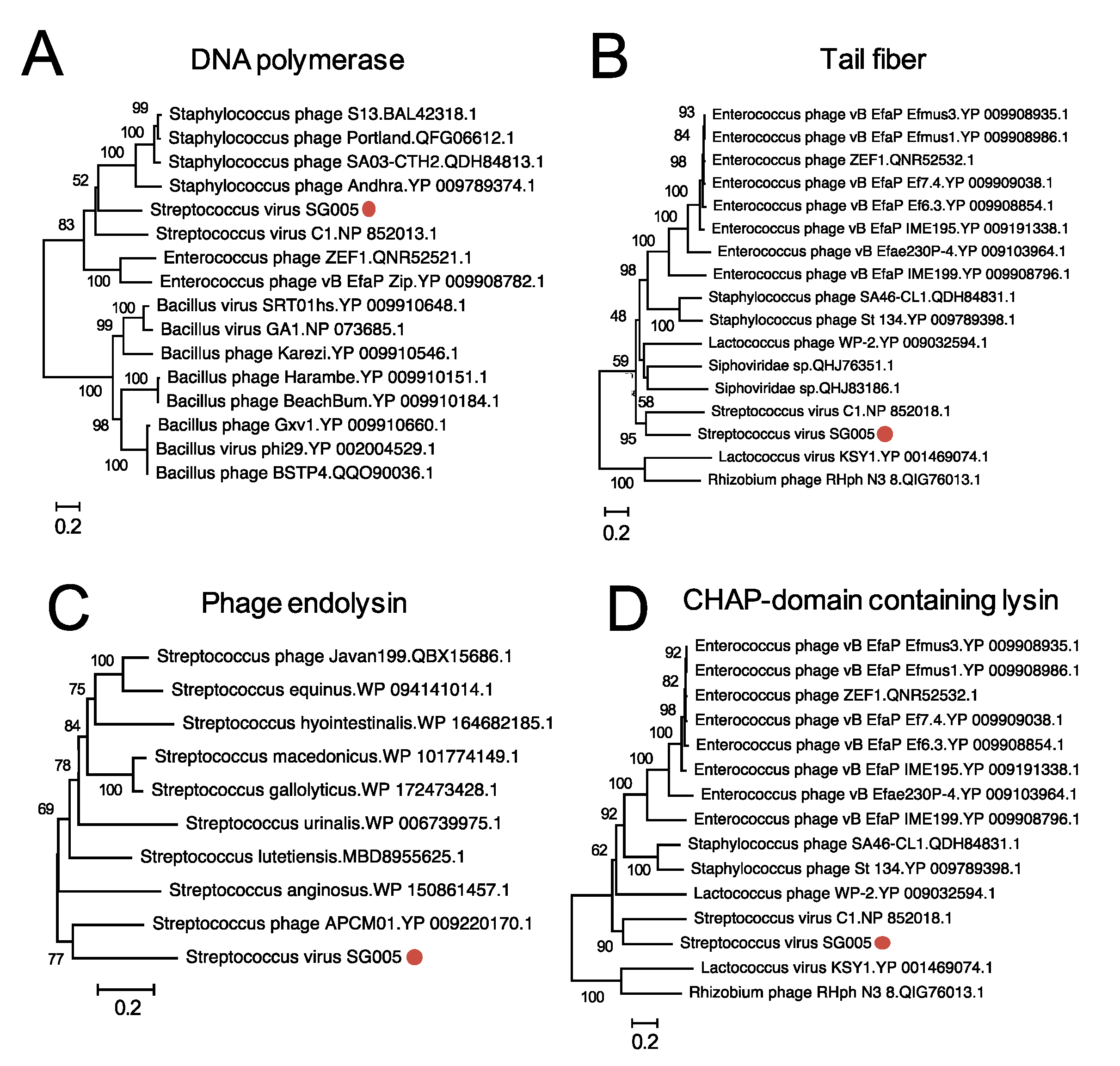
2.8. Bioinformatics Analysis
2.9. Statistical Analysis
3. Results
3.1. Isolation of Streptococcus Virus ΦSG005
3.2. Lytic Activity of Streptococcus Virus ΦSG005
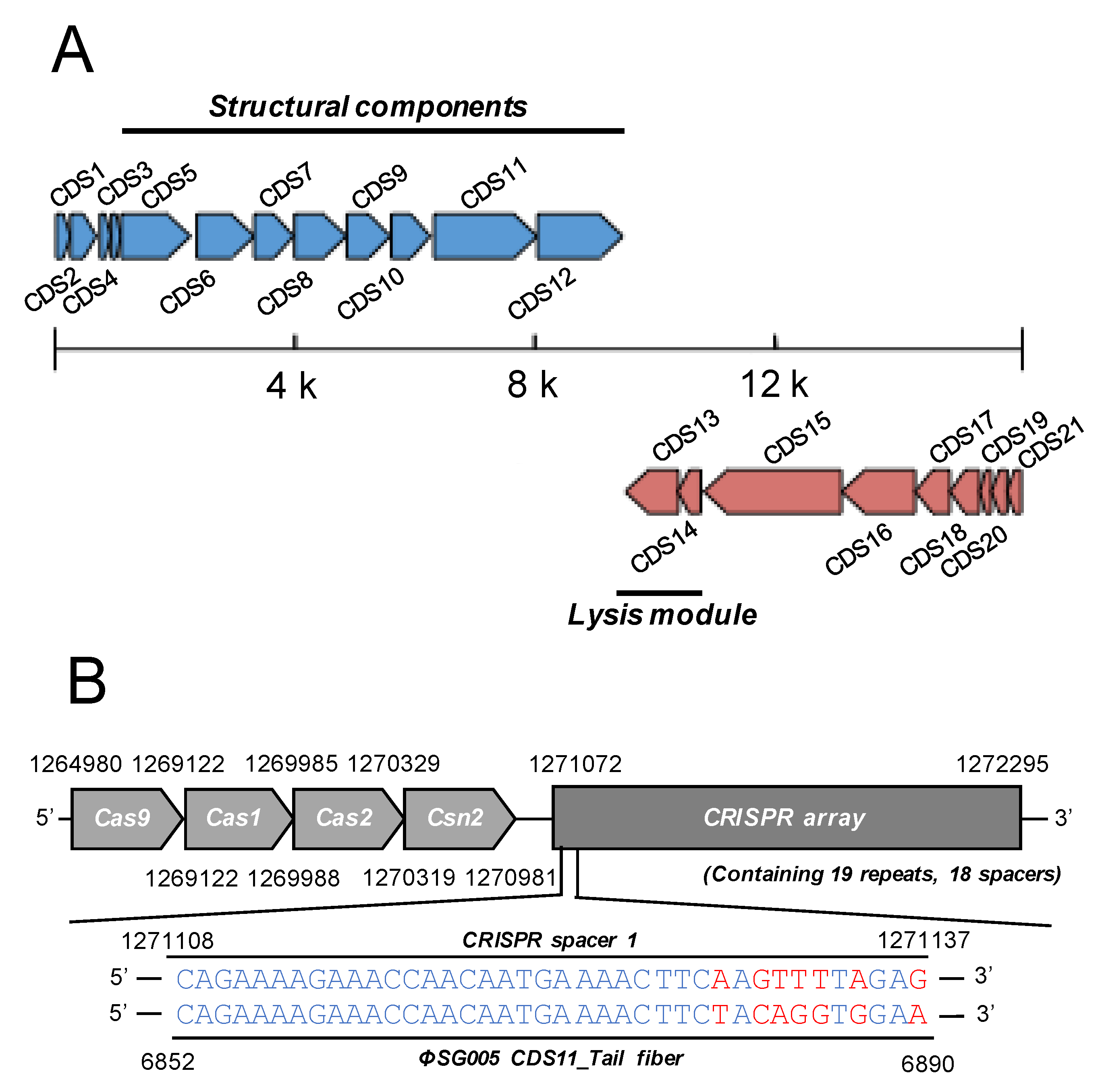
3.3. Genome Organization of ΦSG005
3.4. Prophage Elements in S. gordonii ATCC10558
3.5. Molecular Characterization of ΦSG005
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hampton, H.G.; Watson, B.N.J.; Fineran, P.C. The arms race between bacteria and their phage foes. Nature 2020, 577, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell. Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.F.; Su, B.Y.; Ma, C.H.; Rowley, P.A.; Jayaram, M. A bipartite thermodynamic-kinetic contribution by an activating mutation to rdf-independent excision by a phage serine integrase. Nucleic Acids Res. 2020, 48, 6413–6430. [Google Scholar] [CrossRef] [PubMed]
- Synnott, A.J.; Kuang, Y.; Kurimoto, M.; Yamamichi, K.; Iwano, H.; Tanji, Y. Isolation from sewage influent and characterization of novel staphylococcus aureus bacteriophages with wide host ranges and potent lytic capabilities. Appl. Env. Microbiol. 2009, 75, 4483–4490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furusawa, T.; Iwano, H.; Hiyashimizu, Y.; Matsubara, K.; Higuchi, H.; Nagahata, H.; Niwa, H.; Katayama, Y.; Kinoshita, Y.; Hagiwara, K.; et al. Phage therapy is effective in a mouse model of bacterial equine keratitis. Appl. Env. Microbiol. 2016, 82, 5332–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiki, J.; Furusawa, T.; Munby, M.; Kawaguchi, C.; Matsuda, Y.; Shiokura, Y.; Nakamura, K.; Nakamura, T.; Sasaki, M.; Usui, M.; et al. Susceptibility of pseudomonas aeruginosa veterinary isolates to pbunavirus pb1-like phages. Microbiol. Immunol. 2020, 64, 778–782. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Llorente, C.; Lang, S.; Brandl, K.; Chu, H.; Jiang, L.; White, R.C.; Clarke, T.H.; Nguyen, K.; Torralba, M.; et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature 2019, 575, 505–511. [Google Scholar] [CrossRef]
- Veyrand-Quiros, B.; Guzman-Villanueva, L.T.; Reyes, A.G.; Rodriguez-Jaramillo, C.; Salas-Leiva, J.S.; Tovar-Ramirez, D.; Balcazar, J.L.; Quiroz-Guzman, E. Assessment of bacteriophage vb_pd_pdcc-1 on bacterial dynamics during ontogenetic development of the longfin yellowtail (seriola rivoliana). Appl. Microbiol. Biotechnol. 2021, 105, 2877–2887. [Google Scholar] [CrossRef]
- Shimamori, Y.; Mitsunaka, S.; Yamashita, H.; Suzuki, T.; Kitao, T.; Kubori, T.; Nagai, H.; Takeda, S.; Ando, H. Staphylococcal phage in combination with staphylococcus epidermidis as a potential treatment for staphylococcus aureus-associated atopic dermatitis and suppressor of phage-resistant mutants. Viruses 2020, 13, 7. [Google Scholar] [CrossRef]
- Furfaro, L.L.; Chang, B.J.; Payne, M.S. Applications for bacteriophage therapy during pregnancy and the perinatal period. Front. Microbiol. 2017, 8, 2660. [Google Scholar] [CrossRef] [Green Version]
- Moodley, A.; Kot, W.; Nalgard, S.; Jakociune, D.; Neve, H.; Hansen, L.H.; Guardabassi, L.; Vogensen, F.K. Isolation and characterization of bacteriophages active against methicillin-resistant staphylococcus pseudintermedius. Res. Vet. Sci. 2019, 122, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Giri, S.S.; Yun, S.; Kim, S.W.; Han, S.J.; Kwon, J.; Oh, W.T.; Lee, S.B.; Park, Y.H.; Park, S.C. Two novel bacteriophages control multidrug- and methicillin-resistant staphylococcus pseudintermedius biofilm. Front. Med. (Lausanne) 2021, 8, 524059. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-rna-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, E.; Richter, H.; van der Oost, J.; White, M.F. Biogenesis pathways of rna guides in archaeal and bacterial crispr-cas adaptive immunity. FEMS Microbiol. Rev. 2015, 39, 428–441. [Google Scholar] [CrossRef] [PubMed]
- van der Ploeg, J.R. Characterization of streptococcus gordonii prophage ph15: Complete genome sequence and functional analysis of phage-encoded integrase and endolysin. Microbiology 2008, 154, 2970–2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, G.; Du, D.; Yu, Y.; Zhang, Y.; Qian, Y.; Zhang, W. Pan-genome analysis of streptococcus suis serotype 2 revealed genomic diversity among strains of different virulence. Transbound Emerg. Dis. 2020, 14, 156. [Google Scholar] [CrossRef]
- Rezaei Javan, R.; Ramos-Sevillano, E.; Akter, A.; Brown, J.; Brueggemann, A.B. Prophages and satellite prophages are widespread in streptococcus and may play a role in pneumococcal pathogenesis. Nat. Commun. 2019, 10, 4852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, O.J.; Kwon, Y.; Park, C.; So, Y.J.; Park, T.H.; Jeong, S.; Im, J.; Yun, C.H.; Han, S.H. Streptococcus gordonii: Pathogenesis and host response to its cell wall components. Microorganisms 2020, 8, 1852. [Google Scholar] [CrossRef]
- Back, C.R.; Sztukowska, M.N.; Till, M.; Lamont, R.J.; Jenkinson, H.F.; Nobbs, A.H.; Race, P.R. The streptococcus gordonii adhesin csha protein binds host fibronectin via a catch-clamp mechanism. J. Biol. Chem. 2017, 292, 1538–1549. [Google Scholar] [CrossRef] [Green Version]
- Loo, C.Y.; Corliss, D.A.; Ganeshkumar, N. Streptococcus gordonii biofilm formation: Identification of genes that code for biofilm phenotypes. J. Bacteriol. 2000, 182, 1374–1382. [Google Scholar] [CrossRef] [Green Version]
- Mosailova, N.; Truong, J.; Dietrich, T.; Ashurst, J. Streptococcus gordonii: A rare cause of infective endocarditis. Case Rep. Infect. Dis. 2019, 2019, 7127848. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Hu, H.; Fang, W.; Shi, D.; Liang, C.; Sun, Y.; Gao, G.; Wang, H.; Zhang, Q.; Wang, L.; et al. Detection of pathogens from resected heart valves of patients with infective endocarditis by next-generation sequencing. Int. J. Infect. Dis. 2019, 83, 148–153. [Google Scholar] [CrossRef] [Green Version]
- Washburn, M.R.; White, J.C.; Niven, C.F.; Streptococcus, B.E. Immunological characteristics. J. Bacteriol. 1946, 51, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Bonilla, N.; Rojas, M.I.; Netto Flores Cruz, G.; Hung, S.H.; Rohwer, F.; Barr, J.J. Phage on tap-a quick and efficient protocol for the preparation of bacteriophage laboratory stocks. PeerJ 2016, 4, e2261. [Google Scholar] [CrossRef] [Green Version]
- Khan Mirzaei, M.; Nilsson, A.S. Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Goker, M. Victor: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. Viridic-a novel tool to calculate the intergenomic similarities of prokaryote-infecting viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Ackermann, H.W. 5500 phages examined in the electron microscope. Arch. Virol. 2007, 152, 227–243. [Google Scholar] [CrossRef]
- Clark, P.F.; Clark, A.S. A “bacteriophage” active against a hemolytic streptococcus. J. Bacteriol. 1926, 11, 142. [Google Scholar] [CrossRef]
- Biswas, A.; Gagnon, J.N.; Brouns, S.J.; Fineran, P.C.; Brown, C.M. Crisprtarget: Bioinformatic prediction and analysis of crrna targets. RNA Biol. 2013, 10, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Boudry, P.; Semenova, E.; Monot, M.; Datsenko, K.A.; Lopatina, A.; Sekulovic, O.; Ospina-Bedoya, M.; Fortier, L.C.; Severinov, K.; Dupuy, B.; et al. Function of the crispr-cas system of the human pathogen clostridium difficile. MBio 2015, 6, e01112–e01115. [Google Scholar] [CrossRef] [Green Version]
- Rostol, J.T.; Marraffini, L. (ph)ighting phages: How bacteria resist their parasites. Cell. Host Microbe 2019, 25, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. Rna-guided editing of bacterial genomes using crispr-cas systems. Nat. Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef]
- An, S.Y.; Ka, D.; Kim, I.; Kim, E.H.; Kim, N.K.; Bae, E.; Suh, J.Y. Intrinsic disorder is essential for cas9 inhibition of anti-crispr acriia5. Nucleic Acids Res. 2020, 48, 7584–7594. [Google Scholar] [CrossRef]
- Fortier, L.C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- Paul, V.D.; Rajagopalan, S.S.; Sundarrajan, S.; George, S.E.; Asrani, J.Y.; Pillai, R.; Chikkamadaiah, R.; Durgaiah, M.; Sriram, B.; Padmanabhan, S. A novel bacteriophage tail-associated muralytic enzyme (tame) from phage k and its development into a potent antistaphylococcal protein. BMC Microbiol. 2011, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Fujiki, J.; Nakamura, T.; Furusawa, T.; Ohno, H.; Takahashi, H.; Kitana, J.; Usui, M.; Higuchi, H.; Tanji, Y.; Tamura, Y.; et al. Characterization of the lytic capability of a lysk-like endolysin, lys-phisa012, derived from a polyvalent staphylococcus aureus bacteriophage. Pharmacy 2018, 11, 456. [Google Scholar]
- Lancefield, R.C. Note on the susceptibility of certain strains of hemolytic streptococcus to a streptococcus bacteriophage. Proc. Soc. Exp. Biol. Med. 1932, 30, 169–171. [Google Scholar] [CrossRef]
- Shwartzman, G. Studies on streptococcus bacteriophage: I. A powerful lytic principle against hemolytic streptococci of erysipelas origin. J. Exp. Med. 1927, 46, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.; Loomis, L.; Fischetti, V.A. Prevention and elimination of upper respiratory colonization of mice by group a streptococci by using a bacteriophage lytic enzyme. Proc. Natl. Acad. Sci. USA 2001, 98, 4107–4112. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Component | Length (bp) | G+C (%) | CDS | Accession No. |
|---|---|---|---|---|---|
| ΦSG005 | Viral genome | 16,127 | 34.5 | 21 | LC_628082 |
| ATCC10558 | Chromosome | 2,187,611 | 49.52 | 2039 | NZ_LS483341.1 |
| CDS | Location (Nucleotides) | Strand | Size (No. of Amino Acids) | Product or Predicted Function |
|---|---|---|---|---|
| 1 | 1–237 | + | 79 | hypothetical protein |
| 2 | 239–670 | + | 144 | anti-CRISPR protein AcrIIA5 |
| 3 | 730–906 | + | 59 | hypothetical protein |
| 4 | 921–1079 | + | 53 | Phage protein |
| 5 | 1079–2239 | + | 387 | Phage capsid and scaffold |
| 6 | 2344–3306 | + | 321 | Phage capsid and scaffold |
| 7 | 3299–3955 | + | 219 | lower collar protein |
| 8 | 3972–4841 | + | 290 | hypothetical protein |
| 9 | 4844–5569 | + | 242 | hypothetical protein |
| 10 | 5687–6234 | + | 216 | hypothetical protein |
| 11 | 6297–8021 | + | 575 | Phage tail fiber |
| 12 | 8023–9450 | + | 476 | CHAP domain-containing phage lysin |
| 13 | 10,375–9485 | - | 297 | Phage endolysin |
| 14 | 10,745–10,362 | - | 128 | holin protein |
| 15 | 13,088–10,797 | - | 764 | DNA polymerase (EC 2.7.7.7), phage-associated |
| 16 | 14,318–13,092 | - | 409 | Phage neck |
| 17 | 14,900–14,322 | - | 193 | hypothetical protein |
| 18 | 15,394–14,903 | - | 164 | hypothetical protein |
| 19 | 15,591–15,400 | - | 64 | hypothetical protein |
| 20 | 15,853–15,593 | - | 87 | hypothetical protein |
| 21 | 16,088–15,855 | - | 78 | hypothetical protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujiki, J.; Yoshida, S.-i.; Nakamura, T.; Nakamura, K.; Amano, Y.; Nishida, K.; Nishi, K.; Sasaki, M.; Iwasaki, T.; Sawa, H.; et al. Novel Virulent Bacteriophage ΦSG005, Which Infects Streptococcus gordonii, Forms a Distinct Clade among Streptococcus Viruses. Viruses 2021, 13, 1964. https://doi.org/10.3390/v13101964
Fujiki J, Yoshida S-i, Nakamura T, Nakamura K, Amano Y, Nishida K, Nishi K, Sasaki M, Iwasaki T, Sawa H, et al. Novel Virulent Bacteriophage ΦSG005, Which Infects Streptococcus gordonii, Forms a Distinct Clade among Streptococcus Viruses. Viruses. 2021; 13(10):1964. https://doi.org/10.3390/v13101964
Chicago/Turabian StyleFujiki, Jumpei, Shin-ichi Yoshida, Tomohiro Nakamura, Keisuke Nakamura, Yurika Amano, Keita Nishida, Keitaro Nishi, Michihito Sasaki, Tomohito Iwasaki, Hirofumi Sawa, and et al. 2021. "Novel Virulent Bacteriophage ΦSG005, Which Infects Streptococcus gordonii, Forms a Distinct Clade among Streptococcus Viruses" Viruses 13, no. 10: 1964. https://doi.org/10.3390/v13101964
APA StyleFujiki, J., Yoshida, S.-i., Nakamura, T., Nakamura, K., Amano, Y., Nishida, K., Nishi, K., Sasaki, M., Iwasaki, T., Sawa, H., Komatsuzawa, H., Hijioka, H., & Iwano, H. (2021). Novel Virulent Bacteriophage ΦSG005, Which Infects Streptococcus gordonii, Forms a Distinct Clade among Streptococcus Viruses. Viruses, 13(10), 1964. https://doi.org/10.3390/v13101964