Molecular Characterization of Hemorrhagic Enteritis Virus (HEV) Obtained from Clinical Samples in Western Canada 2017–2018

Abstract
1. Introduction
2. Materials and Methods
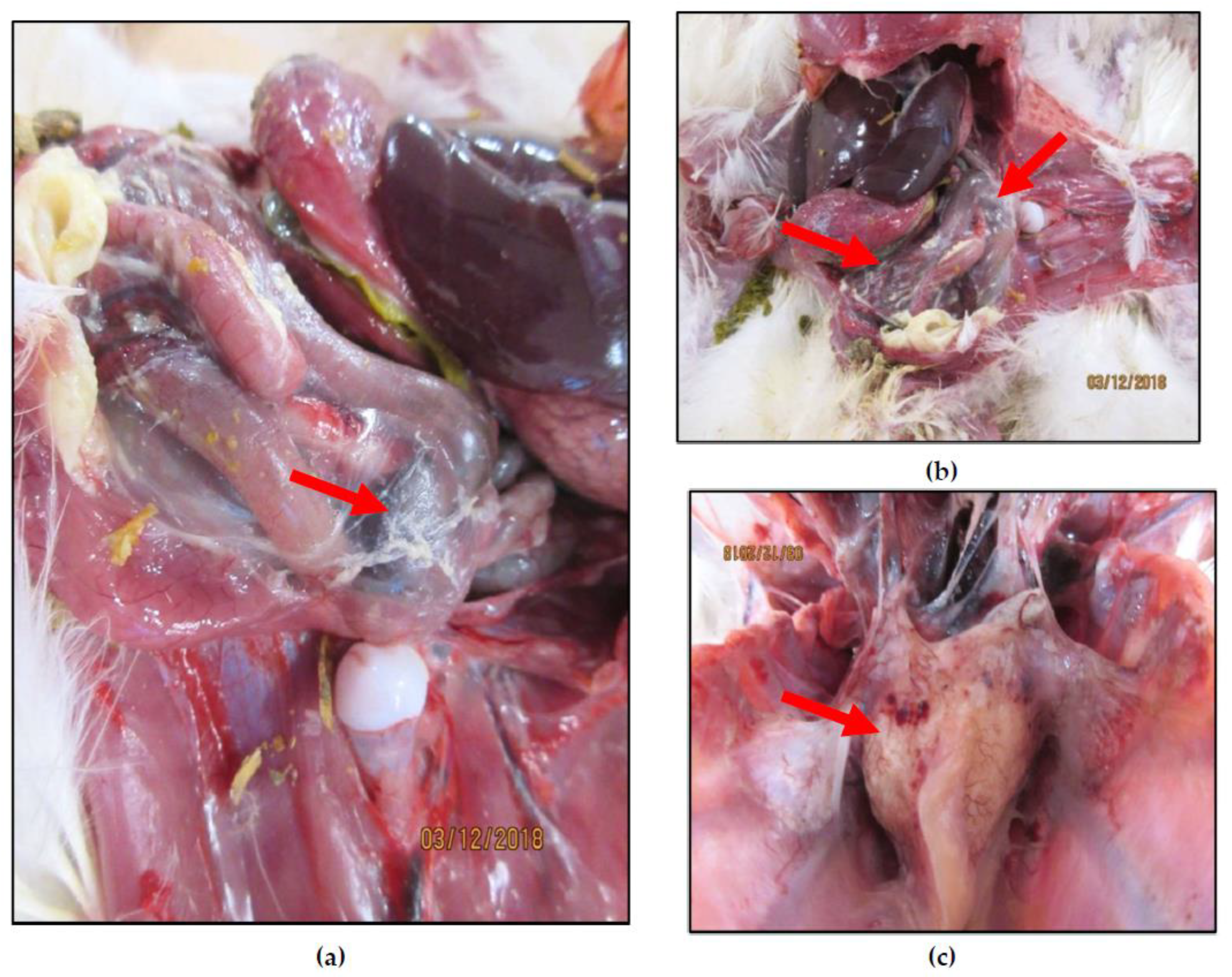
2.1. Sample Collection, Processing, and Ultracentrifugation
2.2. DNA Extraction, PCR, and Sequencing
2.3. Data Analysis
3. Results
3.1. Whole Genome Sequencing
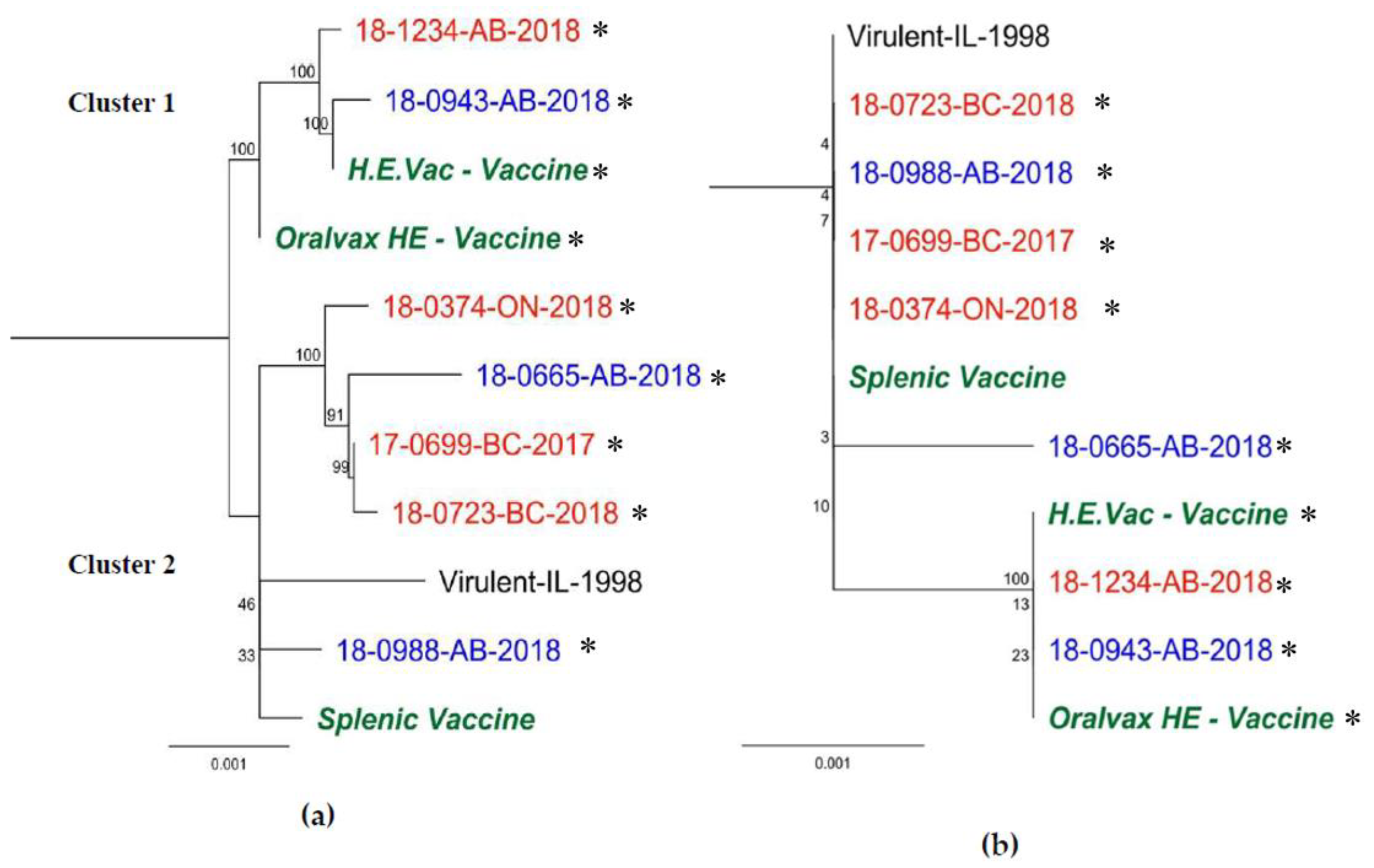
3.2. Hexon Gene
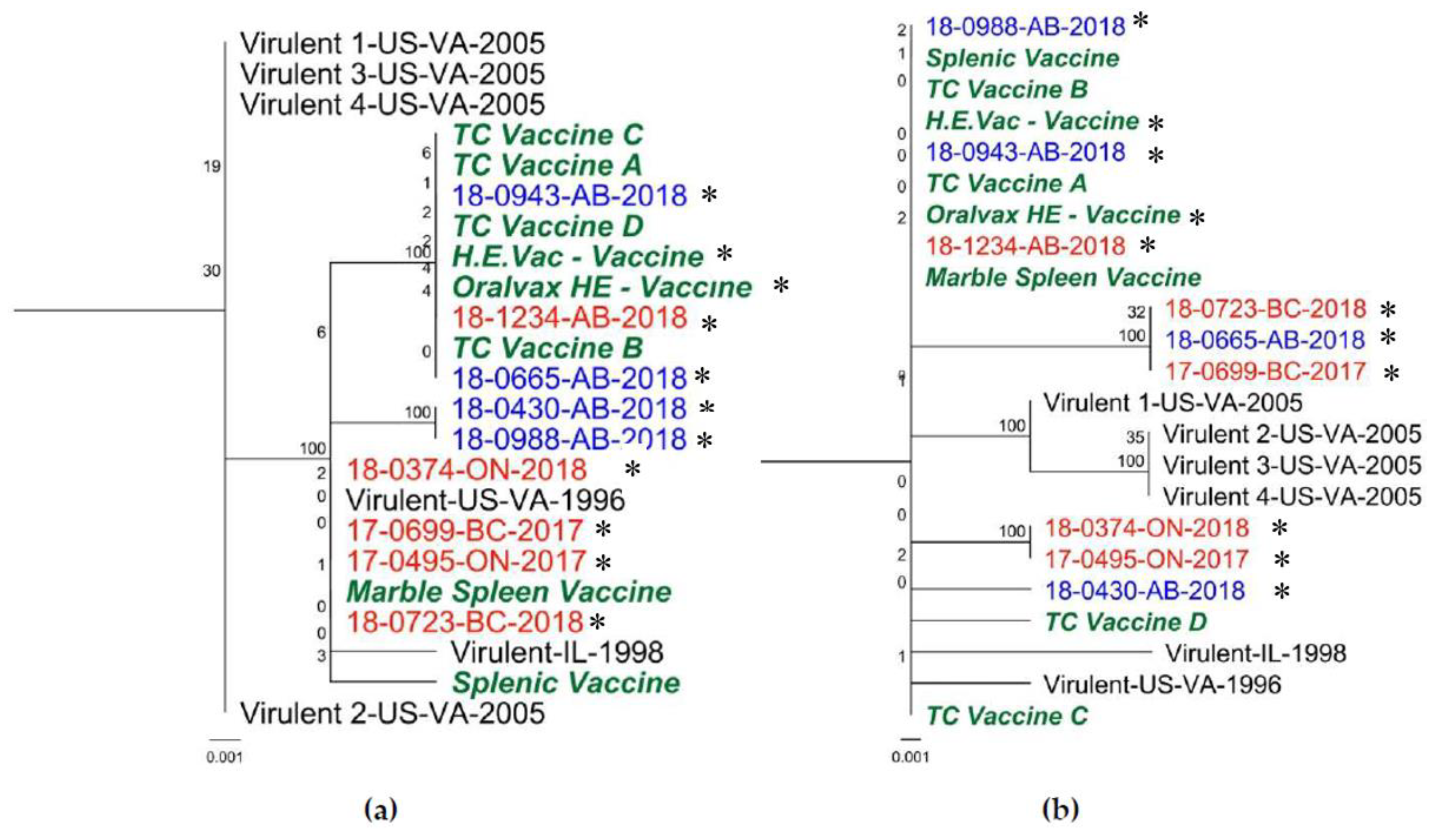
3.3. ORF1 Region
3.4. E3 Gene
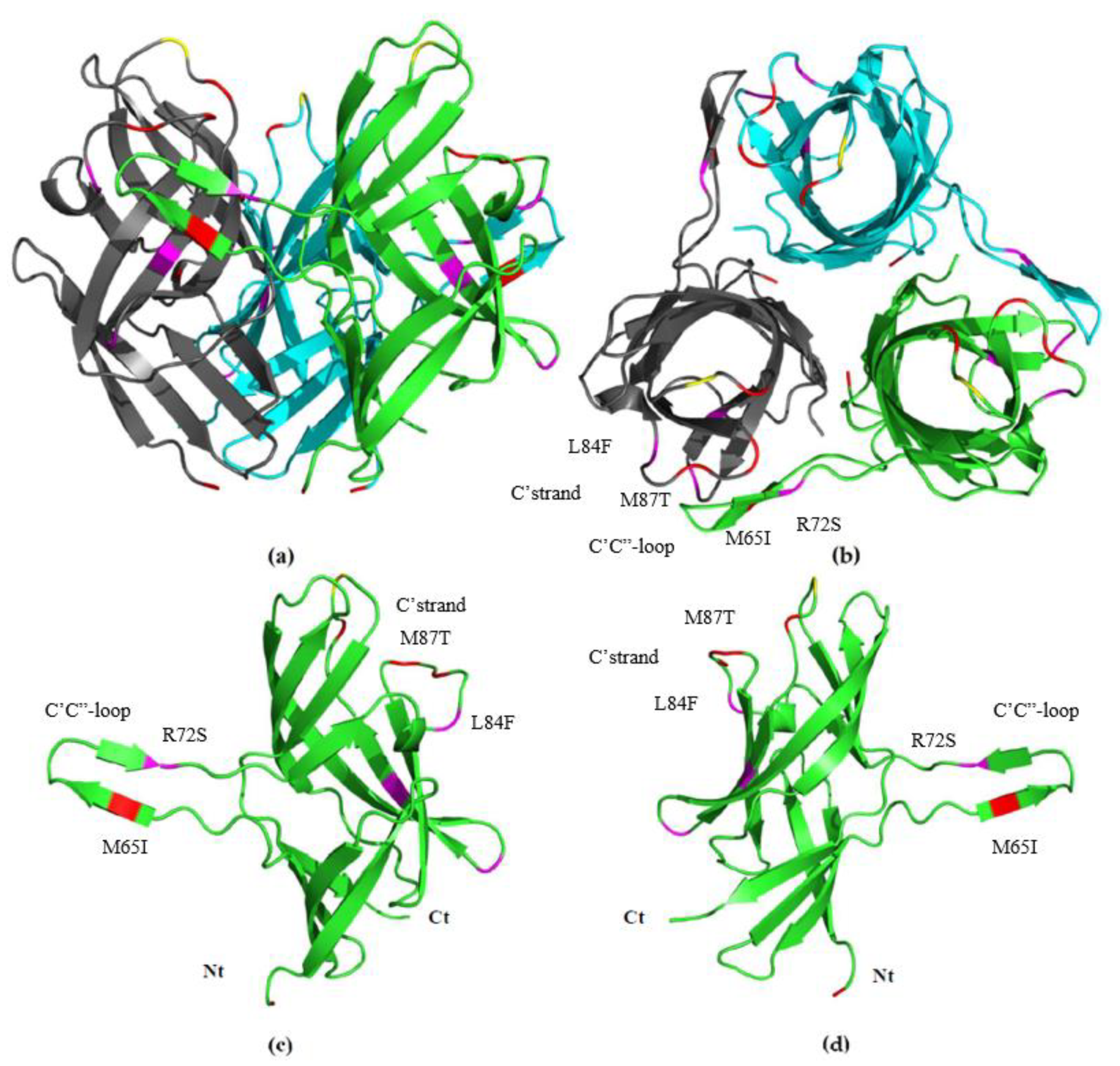
3.5. Fib knob Domain
3.6. pTP
3.7. Prediction of O-Linked Glycosylation Sites in fib Knob by NetOGlyc Service
3.8. Prediction of N-Linked Glycosylation Sites in fib Knob by NetNGlyc Service
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pierson, F.W.; Fitzgerald, S.D. Hemorrhagic Enteritis and Related Infections. In Diseases of Poultry, 13th ed.; Swayne, D.E., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 309–316. [Google Scholar]
- Pitcovski, J.; Mualem, M.; Rei-Koren, Z.; Krispel, S.; Shmueli, E.; Peretz, Y.; Gutter, B.; Gallili, G.; Michael, A.; Goldberg, D. The Complete DNA Sequence and Genome Organization of the Avian Adenovirus, Hemorrhagic Enteritis Virus. Virology 1998, 249, 307–315. [Google Scholar] [CrossRef]
- Pitcovski, J.; Fingerut, E.; Gallili, G.; Eliahu, D.; Finger, A.; Gutter, B. A subunit vaccine against hemorrhagic enteritis adenovirus. Vaccine 2005, 23, 4697–4702. [Google Scholar] [CrossRef]
- Singh, A.K.; Berbís, M.Á.; Ballmann, M.Z.; Kilcoyne, M.; Menendez, M.; Nguyen, T.H.; Joshi, L.; Cañada, F.J.; Jimenez-Barbero, J.; Benkö, M.; et al. Structure and Sialyllactose Binding of the Carboxy-Terminal Head Domain of the Fibre from a Siadenovirus, Turkey Adenovirus. PLoS ONE 2015, 10, e0139339. [Google Scholar] [CrossRef]
- Hurk, J.V.V.D.; Hurk, S.V.D.L.-V.D. Protection of turkeys against haemorrhagic enteritis by monoclonal antibody and hexon immunization. Vaccine 1993, 11, 329–335. [Google Scholar] [CrossRef]
- Dhama, K.; Gowthaman, V.; Karthik, K.; Tiwari, R.; Sachan, S.; Kumar, M.A.; Munuswamy, P.; Malik, Y.S.; Singh, R.K.; Munir, M. Haemorrhagic enteritis of turkeys—Current knowledge. Vet. Q. 2017, 37, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Tykałowski, B.; Śmiałek, M.; Koncicki, A.; Ognik, K.; Zduńczyk, Z.; Jankowski, J. The immune response of young turkeys to haemorrhagic enteritis virus infection at different levels and sources of methionine in the diet. BMC Vet. Res. 2019, 15, 387. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, S.D.; Rautenschlein, S.; Mahsoub, H.M.; Pierson, F.W.; Reed, W.M.; Jack, S.W. Adenovirus Infections. In Diseases of Poultry; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 321–363. [Google Scholar]
- Koncicki, A.; Tykałowski, B.; Stenzel, T.; Śmiałek, M.; Pestka, D. Effect of infection of turkeys with haemorrhagic enteritis adenovirus isolate on the selected parameters of cellular immunity and the course of colibacillosis. Pol. J. Vet. Sci. 2012, 15, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Kumar, A. Haemorrhagic enteritis of turkeys and related infections of pheasants and domestic fowl: A review. World’s Poult. Sci. J. 1998, 54, 253–269. [Google Scholar] [CrossRef]
- Kaboudi, K. Virus-induced immunosuppression in turkeys (Meleagris gallopavo): A review. Open Vet. J. 2019, 9, 349–360. [Google Scholar] [CrossRef]
- Giovanardi, D.; Lupini, C.; Pesente, P.; Rossi, G.; Ortali, G.; Catelli, E. Longitudinal field studies of Avian Metapneumovirus and Turkey Hemorrhagic Enteritis Virus in turkeys suffering from colibacillosis associated mortality. Vet. Res. Commun. 2014, 38, 129–137. [Google Scholar] [CrossRef]
- Tykałowski, B.; Koncicki, A. Studies concerning the role of Hemorrhagic Enteritis Virus in the pathology of turkeys conducted in the Department of Poultry Diseases in Olsztyn over the last 30 years. Med. Weter 2017, 73, 522–527. [Google Scholar] [CrossRef]
- Saif, Y.M. Infectious bursal disease and hemorrhagic enteritis. Poult. Sci. 1998, 77, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Hoerr, F.J. Clinical aspects of immunosuppression in poultry. Avian Dis. 2010, 54, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Iltis, J.P.; Jakowski, R.M.; Wyand, D.S. Transmission of marble spleen disease in turkeys and pheasants. Am. J. Vet. Res. 1975, 36, 97–101. [Google Scholar] [PubMed]
- Gross, W.B. Lesions of Hemorrhagic Enteritis. Avian Dis. 1967, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Gross, W.B.; Moore, W.E.C. Hemorrhagic Enteritis of Turkeys. Avian Dis. 1967, 11, 296. [Google Scholar] [CrossRef]
- Itakura, C.; Carlson, H.C.; Lang, G.N. Experimental transmission of haemorrhagic enteritis of turkeys. Avian pathology. Avian Pathol. 1974, 3, 279–292. [Google Scholar] [CrossRef]
- Beach, N.M.; Duncan, R.; Larsen, C.; Meng, X.-J.; Sriranganathan, N.; Pierson, F. Persistent Infection of Turkeys with an Avirulent Strain of Turkey Hemorrhagic Enteritis Virus. Avian Dis. Dig. 2009, 4, 370–375. [Google Scholar] [CrossRef]
- Domermuth, C.H.; Gross, W.B. Effect of Chlorine on the Virus of Hemorrhagic Enteritis of Turkeys. Avian Dis. 1972, 16, 952. [Google Scholar] [CrossRef]
- Domermuth, C.H.; Gross, W.B. Effect of Disinfectants and Drying on the Virus of Hemorrhagic Enteritis of Turkeys. Avian Dis. 1971, 15, 94. [Google Scholar] [CrossRef]
- Fitzgerald, S.D.; Reed, W.M. Pathogenesis of Marble Spleen Disease in Bursectomized and Non-Bursectomized Ring-Necked Pheasants Following Oral Inoculation with Cell-Culture-Propagated Virus. Avian Dis. 1991, 35, 579. [Google Scholar] [CrossRef]
- Hussain, I.; Choi, C.U.; Rings, B.S.; Shaw, D.P.; Nagaraja, K.V. Pathogenesis of Hemorrhagic Enteritis Virus Infection in Turkeys. J. Vet. Med. Ser. B 1993, 40, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Rautenschlein, S.; Suresh, M.; Neumann, U.; Sharma, J. Comparative pathogenesis of haemorrhagic enteritis virus (HEV) infection in Turkeys and chickens. J. Comp. Pathol. 1998, 119, 251–261. [Google Scholar] [CrossRef]
- Suresh, M.; Sharma, J. Hemorrhagic enteritis virus induced changes in the lymphocyte subpopulations in turkeys and the effect of experimental immunodeficiency on viral pathogenesis. Vet. Immunol. Immunopathol. 1995, 45, 139–150. [Google Scholar] [CrossRef]
- Rautenschlein, S.; Suresh, M.; Sharma, J.M. Pathogenic avian adenovirus type II induces apoptosis in turkey spleen cells. Arch. Virol. 2000, 145, 1671–1683. [Google Scholar] [CrossRef] [PubMed]
- Rautenschlein, S. Immunopathogenesis of haemorrhagic enteritis virus (HEV) in turkeys. Dev. Comp. Immunol. 2000, 24, 237–246. [Google Scholar] [CrossRef]
- Sponenberg, D.P.; Domermuth, C.H.; Larsen, C.T. Field Outbreaks of Colibacillosis of Turkeys Associated with Hemorrhagic Enteritis Virus. Avian Dis. 1985, 29, 838. [Google Scholar] [CrossRef]
- Larsen, C.T.; Domermuth, C.H.; Sponenberg, D.P.; Gross, W.B. Colibacillosis of turkeys exacerbated by hemorrhagic enteritis virus. Laboratory studies. Avian Dis. 1985, 29, 729–732. [Google Scholar] [CrossRef]
- Ranganathan, S.; Gribskov, M.; Nakai, K.; Schönbach, C. Encyclopedia of Bioinformatics and Computational Biology; Ranganathan, S., Gribskov, M., Nakai, K., Schönbach, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Maurier, F.; Beury, D.; Fléchon, L.; Varré, J.-S.; Touzet, H.; Goffard, A.; Hot, D.; Caboche, S. A complete protocol for whole-genome sequencing of virus from clinical samples: Application to coronavirus OC43. Virology 2019, 531, 141–148. [Google Scholar] [CrossRef]
- Nazerian, K.; Fadly, A.M. Further Studies on in vitro and in vivo Assays of Hemorrhagic Enteritis Virus (HEV). Avian Dis. 1987, 31, 234. [Google Scholar] [CrossRef]
- Van den Hurk, J.V. Propagation of Group II Avian Adenoviruses in Turkey and Chicken Leukocytes. Avian Dis. 1990, 34, 12. [Google Scholar] [CrossRef] [PubMed]
- Domermuth, C.H.; Gross, W.B.; Douglass, C.S.; DuBose, R.T.; Harris, J.R.; Davis, R.B. Vaccination for Hemorrhagic Enteritis of Turkeys. Avian Dis. 1977, 21, 557. [Google Scholar] [CrossRef] [PubMed]
- Weier, S. Improved immunoprophylaxis against Haemorrhagic Enteritis Virus (HEV) in turkeys with repeated drinking water vaccination. Prakt. Tierarzt. 2013, 94, 739. [Google Scholar]
- Barbour, E.K.; Poss, P.E.; Brinton, M.K.; Johnson, J.B.; Nabbut, N.H. Evaluation of cell culture propagated and in vivo propagated hemorrhagic enteritis vaccines in turkeys. Vet. Immunol. Immunopathol. 1993, 35, 375–383. [Google Scholar] [CrossRef]
- Agriculture and Agri-Food Canada (AAFC). Canada’s Poultry and Egg Industry Profile. Available online: https://www.agr.gc.ca/eng/animal-industry/poultry-and-eggs/poultry-and-egg-market-information/industry-profile/?id=1384971854389 (accessed on 15 July 2020).
- Austin, A. US Turkey Production Falls by 5% in 2019. Available online: https://www.wattagnet.com/articles/39503-us-turkey-production-falls-by-5-in-2019?v=preview#:~:text=According%20to%20the%20annual%20survey,industry%20produced%207.451%20billion%20pounds (accessed on 15 July 2020).
- Beach, N.M.; Duncan, R.B.; Larsen, C.T.; Meng, X.-J.; Sriranganathan, N.; Pierson, F.W. Comparison of 12 turkey hemorrhagic enteritis virus isolates allows prediction of genetic factors affecting virulence. J. Gen. Virol. 2009, 90, 1978–1985. [Google Scholar] [CrossRef]
- Alkie, T.N.; Guenther, R.; Rautenschlein, S. Molecular Characterization of Hemorrhagic Enteritis Viruses (HEV) Detected in HEV-Vaccinated Commercial Turkey Flocks in Germany. Avian Dis. 2017, 61, 96–101. [Google Scholar] [CrossRef]
- Hybrid. Technical Guide for Hybrid Turkeys Commercial Products; Hybrid Turkeys, Ed.; Hendrix Genetics: Mauges-sur-Loire, France, 2020. [Google Scholar]
- Boulianne, M.; American Association of Avian Pathologists. Avian Disease Manual, 7th ed.; American Association of Avian Pathologists: Jacksonville, FL, USA, 2013; p. 300. [Google Scholar]
- Mahsoub, H.; Evans, N.P.; Beach, N.M.; Yuan, L.; Zimmerman, K.L.; Pierson, F.W. Real-time PCR-based infectivity assay for the titration of turkey hemorrhagic enteritis virus, an adenovirus, in live vaccines. J. Virol. Methods 2017, 239, 42–49. [Google Scholar] [CrossRef]
- Manninen, A.; Verkade, P.; Le Lay, S.; Torkko, J.; Kasper, M.; Füllekrug, J.; Simons, K. Caveolin-1 Is Not Essential for Biosynthetic Apical Membrane Transport. Mol. Cell. Biol. 2005, 25, 10087–10096. [Google Scholar] [CrossRef]
- Optiprep Application Sheet V07—Purification of Group I (ds)DNA Viruses: Adenovirus and Removal of Helper Virus. Available online: https://www.axis-shield-density-gradient-media.com/V07.pdf (accessed on 1 February 2020).
- Mahsoub, H.M. Real Time PCR-Based Infectivity Assay and Characterization of Cell Surface Receptors for Turkey Hemorrhagic Enteritis Virus; Virginia Tech: Blacksburg, VA, USA, 2016. [Google Scholar]
- Kesmen, Z.; Yetiman, A.E.; Sahin, F.; Yetim, H. Detection of Chicken and Turkey Meat in Meat Mixtures by Using Real-Time PCR Assays. J. Food Sci. 2012, 77, 167–173. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Xu, L.; Benson, S.D.; Burnett, R.M. Nanoporous crystals of chicken embryo lethal orphan (CELO) adenovirus major coat protein, hexon. J. Struct. Biol. 2007, 157, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Rux, J.J.; Kuser, P.R.; Burnett, R.M. Structural and Phylogenetic Analysis of Adenovirus Hexons by Use of High-Resolution X-Ray Crystallographic, Molecular Modeling, and Sequence-Based Methods. J. Virol. 2003, 77, 9553–9566. [Google Scholar] [CrossRef]
- Rux, J.J.; Burnett, R.M. Type-Specific Epitope Locations Revealed by X-Ray Crystallographic Study of Adenovirus Type 5 Hexon. Mol. Ther. 2000, 1, 18–30. [Google Scholar] [CrossRef]
- Chapter 10—Adenoviridae. In Fenner’s Veterinary Virology, 5th ed.; MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 217–227. [Google Scholar]
- Crawford-Miksza, L.; Schnurr, D.P. Analysis of 15 adenovirus hexon proteins reveals the location and structure of seven hypervariable regions containing serotype-specific residues. J. Virol. 1996, 70, 1836–1844. [Google Scholar] [CrossRef]
- Stewart, P.; Fuller, S.; Burnett, R. Difference imaging of adenovirus: Bridging the resolution gap between X-ray crystallography and electron microscopy. EMBO J. 1993, 12, 2589–2599. [Google Scholar] [CrossRef]
- Blom, N.S.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef]
- Beach, N.M. Characterization of Avirulent Turkey Hemorrhagic Enteritis Virus: A Study of the Molecular Basis for Variation in Virulence and the Occurrence of Persistent Infection; Virginia Polytechnic Institute and State University Virginia Tech: Blacksburg, VA, USA, 2006. [Google Scholar]
- Yang, L.; Connaris, H.; Potter, J.A.; Taylor, G.L. Structural characterization of the carbohydrate-binding module of NanA sialidase, a pneumococcal virulence factor. BMC Struct. Biol. 2015, 15, 15. [Google Scholar] [CrossRef]
- Rothe, B.; Roggentin, P.; Schauer, R. The sialidase gene from Clostridium septicum: Cloning, sequencing, expression in Escherichia coli and identification of conserved sequences in sialidases and other proteins. Mol. Genet. Genom. 1991, 226, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Von Itzstein, M.; Dyason, J.C.; Oliver, S.W.; White, H.F.; Wu, W.-Y.; Kok, G.B.; Pegg, M.S. A Study of the Active Site of Influenza Virus Sialidase: An Approach to the Rational Design of Novel Anti-influenza Drugs. J. Med. Chem. 1996, 39, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.J. Factors promoting pathogenicity of influenza virus. Semin. Respir. Infect. 1991, 6, 3–10. [Google Scholar]
- Aboezz, Z.R.; Mahsoub, H.; El-Bagoury, G.; Pierson, F.W. In vitro growth kinetics and gene expression analysis of the turkey adenovirus 3, a siadenovirus. Virus Res. 2019, 263, 47–54. [Google Scholar] [CrossRef]
- Kumar, P.; Hurk, J.V.D.; Ayalew, L.E.; Gaba, A.; Tikoo, S.K. Proteomic analysis of purified turkey adenovirus 3 virions. Vet. Res. 2015, 46, 79. [Google Scholar] [CrossRef][Green Version]
- Arnberg, N.; Mei, Y.-F.; Wadell, G. Fiber Genes of Adenoviruses with Tropism for the Eye and the Genital Tract. Virology 1997, 227, 239–244. [Google Scholar] [CrossRef]
- Zheng, J.; Yamada, Y.; Fung, T.S.; Huang, M.; Chia, R.; Liu, D.X. Identification of N-linked glycosylation sites in the spike protein and their functional impact on the replication and infectivity of coronavirus infectious bronchitis virus in cell culture. Virology 2018, 513, 65–74. [Google Scholar] [CrossRef]
- Van den Hurk, J.V.; van Drunen Little-van den Hurk, S. Characterization of group II avian adenoviruses with a panel of monoclonal antibodies. Can. J. Vet. Res. Rev. Can. Rech. Vet. 1988, 52, 458–467. [Google Scholar]
- Webster, A.; Leith, I.R.; Nicholson, J.; Hounsell, J.; Hay, R.T. Role of preterminal protein processing in adenovirus replication. J. Virol. 1997, 71, 6381–6389. [Google Scholar] [CrossRef]
- Fadly, A.M.; Nazerian, K.; Nagaraja, K.; Below, G. Field Vaccination against Hemorrhagic Enteritis of Turkeys by a Cell-Culture Live-Virus Vaccine. Avian Dis. 1985, 29, 768. [Google Scholar] [CrossRef]
- Sanjuán, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell. Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [PubMed]
- Sanjuán, R.; Nebot, M.R.; Chirico, N.; Mansky, L.M.; Belshaw, R. Viral Mutation Rates. J. Virol. 2010, 84, 9733–9748. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Tissue | Province/Source | Age (Days) | Clinical Case | HE Vaccination Program a | Type of Sequence |
|---|---|---|---|---|---|---|
| H.E. Vac | Vaccine | Arko Labs | N/A | N/A | N/A | Vaccine |
| Oralvax HE | Vaccine | MSD | N/A | N/A | N/A | Vaccine |
| 17-0495 | Spleen | ON | 44 | ↑ Mortality-Surveillance | No | Field |
| 17-0699 | Spleen | BC | 69 | ↑ Mortality-Surveillance | No | Field |
| 18-0374 | Spleen | AB | 52 | ↑ Mortality-Systemic Bacterial Infections. Escherichia coli in Pericardium | No | Field |
| 18-0430 | Spleen | AB | 110 | ↑ Mortality-Cellulitis- Escherichia coli; Staphylococcus aureus; Enterococcus faecalis; Lactobacillus agilis in Subcutaneous tissue | Yes | Field |
| 18-0665 | Spleen | AB | 91 | ↑ Mortality-Systemic Bacterial Infections-Escherichia coli in Air Sac and Liver | Yes | Field |
| 18-0723 | Spleen | BC | 62 | ↑ Mortality-Surveillance Escherichia coli in Air Sac and Liver | No | Field |
| 18-0943 | Spleen | AB | 61 | ↑ Mortality-Gangrenous Dermatitis Escherichia coli; Staphylococcus saprophyticus; Bacillus pumilus; Bacillus altitudinis; Staphylococcus chromogenes; Staphylococcus chromogenes; Clostridium perfringes; Staphylococcus lentus in Subcutaneous tissue | Yes | Vaccine |
| 18-0988 | Spleen | AB | 117 | ↑ Mortality-Cellulitis- Escherichia coli; Staphylococcus aureus; Enterococcus in Subcutaneous tissue | Yes | Field |
| 18-1234 | Spleen | AB | 77 | ↑ Mortality-Systemic Bacterial Infections- Escherichia coli in Pericardium and Air Sac | No | Vaccine |
| Name | Sequence | Target | Reference | Position | Amplicon | Annealing T | Extension |
|---|---|---|---|---|---|---|---|
| THEV-Whole-F1 | ATGCTTGGGAGGGGATTTCG | THEV | This study | 21–40 | 26,129 | 59 °C | 14 min |
| THEV-Whole-R1 | AACCGGAAAAGAAGGCGGAT | THEV | This study | 26,131–26,150 | |||
| Alkie-HEV-ORF1-F1 | CTGACCTTGTCGTCCGTGC | ORF1 | [41] | 283–301 | 1537 | 62 °C | 2 min |
| HEV3’For-951 | TGGCGGCAATGGCTTAGTAA | ORF1 | This study | 951–970 | |||
| Alkie-HEV3’Rev | GGATACAATTGACCATTGGAAG | ORF1 | [41] | 1799–1820 | |||
| THEV-E3-Fw | CTCCCCTAGTCACCTGACCA | E3 | This study | 20,738–20,757 | 1807 | 59 °C | 2 min |
| THEV-E3-Rv | AACGCTTTCCAGGAGTAGCC | E3 | This study | 22,525–22,544 | |||
| THEV-Fib-Fw | GGCTACTCCTGGAAAGCGTT | Fib | This study | 22,525–22,544 | 2021 | 59 °C | 2.5 min |
| THEV-Fib-Rv | GTCAGCTTGCAACCACCAAG | Fib | This study | 24,550–24,569 | |||
| THEV-Fib-Fw | GGCTACTCCTGGAAAGCGTT | Fib | This study | 22,525–22,544 | 1502 | 59 °C | 2 min |
| THEV-Fib-Rv2 | GCGCACCTGCAAAGTCAAAT | Fib | This study | 24,007–24,026 |
| Sequence a | Phylogenetic Tree | GenBank Number | Genome Size (nt) | Paper Published | ||||
|---|---|---|---|---|---|---|---|---|
| Whole Genome | Hexon | ORF1 | E3 | fib Knob Domain | ||||
| H.E.Vac | X | X | X | X | X | MT603863 MT603864 MT603862 MT603869 MT603871 MT603861 MT603865 MT603870 MT603866 MT603867 MT603868 | 26,289 | This study |
| Oralvax HE | X | X | X | X | X | 26,270 | ||
| 17-0495-ON-2017 | X | X | X | 3850 | ||||
| 17-0699-BC-2017 | X | X | X | X | 26,115 | |||
| 18-0374-AB-2018 | X | X | X | X | 25,997 | |||
| 18-0430-AB-2018 | X | X | X | 3850 | ||||
| 18-0665-AB-2018 | X | X | X | X | 25,717 | |||
| 18-0723-BC-2018 | X | X | X | X | 26,115 | |||
| 18-0943-AB-2018 | X | X | X | X | 26,289 | |||
| 18-0988-AB-2018 | X | X | X | X | 26,100 | |||
| 18-1234-AB-2018 | X | X | X | X | 26,120 | |||
| Virulent-IL-1998 b | X | X | X | X | X | AF074946 | 26,263 | [2] |
| Splenic Vaccine | X | X | X | X | X | AY849321 | 26,266 | [40] |
| Virulent-US-VA-1996 b | X | X | X | DQ868929 | 3857 | |||
| Virulent1-US-VA-2005 b | X | X | X | DQ868931 | 3857 | |||
| Virulent2-US-VA-2005 b | X | X | X | DQ868932 | 3857 | |||
| Virulent3-US-VA-2005 b | X | X | X | DQ868933 | 3857 | |||
| Virulent4-US-VA-2005 b | X | X | X | DQ868934 | 3857 | |||
| Marble spleen vaccine | X | X | X | DQ868930 | 3857 | |||
| Tissue culture vaccine A | X | X | X | DQ868935 | 3857 | |||
| Tissue culture vaccine B | X | X | X | DQ868936 | 3857 | |||
| Tissue culture vaccine C | X | X | X | DQ868937 | 3857 | |||
| Tissue culture vaccine D | X | X | X | DQ868938 | 3857 | |||
| Case1-DE-1989 | X | KX944266 | 1481 | [41] | ||||
| Case2-DE-2010 | X | KX944267 | 1388 | |||||
| Case3-DE-2012 | X | KX944268 | 1433 | |||||
| Case4-DE-2008 | X | KX944269 | 1412 | |||||
| Case5-DE-2008 | X | KX944270 | 1296 | |||||
| Case6-DE-2008 | X | KX944271 | 1385 | |||||
| Case7-DE-2008 | X | KX944272 | 1492 | |||||
| Case8-DE-2008 | X | KX944273 | 1492 | |||||
| Case9-DE-UNK | X | KX944274 | 1326 | |||||
| Case10-DE-2008 | X | KX944275 | 1326 | |||||
| Case11-DE-2008 | X | KX944276 | 1326 | |||||
| Case12-DE-2008 | X | KX944277 | 1493 | |||||
| Case13-DE-2012 | X | KX944278 | 1274 | |||||
| Case14-DE-2012 | X | KX944279 | 1265 | |||||
| Case15-DE-2012 | X | KX944280 | 1265 | |||||
| Case16-DE-2008 | X | KX944281 | 1385 | |||||
| Case17-DE-2012 | X | KX944282 | 1500 | |||||
| ID | Type of Sequence | O-Glycosylation Site | Score | O-Glyc Results |
|---|---|---|---|---|
| H.E. Vac Oralvax HE TC Vaccine A TC Vaccine B TC Vaccine C TC Vaccine D Marble Spleen Vaccine Splenic Vaccine 18-1234 18-0943 | Vaccine | 13 | 0.76–0.77 | Positive-6 locations |
| 18 | 0.63 | |||
| 19 | 0.61 | |||
| 22 | 0.54 | |||
| 24 | 0.65 | |||
| 26 | 0.71 | |||
| 17-0495 17-0699 18-0374 18-0665 18-0723 18-0988 Virulent-US-VA-2005 Virulent-1-US-VA-2005 Virulent-2-US-VA-2005 Virulent-3-US-VA-2005 Virulent-4-US-VA-2005 | Field | 13 | 0.76–0.77 | Positive-6 locations |
| 18 | 0.62–0.66 | |||
| 19 | 0.59–0.61 | |||
| 22 | 0.53–0.59 | |||
| 24 | 0.62–0.66 | |||
| 26 | 0.65–0.72 | |||
| 18-0430 Virulent-IL-1998 | Field | 10 | 0.5 | Positive-7 locations |
| 13 | 0.77 | |||
| 18 | 0.63–0.64 | |||
| 19 | 0.62 | |||
| 22 | 0.54–0.55 | |||
| 24 | 0.66 | |||
| 26 | 0.71–0.72 |
| ID | Type of Sequence | N-Glycosylation Site | Potential | Agreement | N-Glyc Results c | Comments |
|---|---|---|---|---|---|---|
| H.E. Vac Oralvax HE TC Vaccine A TC Vaccine B TC Vaccine C Marble Spleen Vac. Splenic Vaccine 18-1234 18-0943 18-0988 | Vaccine Vaccine Vaccine Vaccine Vaccine Vaccine Vaccine Vaccine Vaccine Field | 32-NGQF | 0.6786 | (9/9) | ++ | |
| 61-NIGV | 0.7398 | (9/9) | ++ | |||
| 67-NPTF | 0.5111 | (6/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6818 | (9/9) | ++ | Sequon ASN-XAA-SER/THR | ||
| 89-NNTY | 0.6219 | (8/9) | + | |||
| 90-NTYI | 0.6121 | (8/9) | + | |||
| 97-NGGV | 0.6647 | (9/9) | ++ | |||
| 117-NNSS | 0.5110 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4376 | (7/9) | - | |||
| 133-NGNP | 0.1234 | (9/9) | --- | |||
| 135-NPHM | 0.5491 | (7/9) | + | PRO-X1 | ||
| 143-NPVP | 0.1229 | (9/9) | --- | |||
| 148-NIKM | 0.6002 | (8/9) | + | |||
| TC Vaccine D | Vaccine | 32-NGQF | 0.6784 | (9/9) | ++ | |
| 61-NIGV | 0.7398 | (9/9) | ++ | |||
| 67-NPTF | 0.5111 | (6/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6818 | (9/9) | ++ | Sequon ASN-XAA-SER/THR | ||
| 89-NNTY | 0.6219 | (8/9) | + | |||
| 90-NTYI | 0.6121 | (8/9) | + | |||
| 97-NGGV | 0.6648 | (9/9) | ++ | |||
| 117-NNSS | 0.5110 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4377 | (7/9) | - | |||
| 133-NGNP | 0.1234 | (9/9) | --- | |||
| 135-NPHM | 0.5493 | (7/9) | + | PRO-X1 | ||
| 143-NPVS | 0.5489 | (6/9) | + | |||
| 148-NIKM | 0.5516 | (6/9) | + | |||
| Virulent-IL-1998 | Field | 32-NGQF | 0.6786 | (9/9) | ++ | |
| 61-NIGV | 0.7589 | (9/9) | +++ | |||
| 67-NPTF | 0.5039 | (4/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6810 | (9/9) | ++ | Sequon ASN-XAA-SER/THR | ||
| 89-NNTY | 0.6149 | (7/9) | + | |||
| 90-NTYI | 0.5604 | (7/9) | + | |||
| 97-NGGV | 0.6506 | (9/9) | ++ | |||
| 117-NNSS | 0.5112 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4377 | (7/9) | - | |||
| 133-NGNP | 0.1233 | (9/9) | --- | |||
| 135-NPHM | 0.5494 | (7/9) | + | PRO-X1 | ||
| 143-NPVP | 0.1229 | (9/9) | --- | |||
| 148-NIKM | 0.6004 | (8/9) | + | |||
| Virulent-US-VA-1996 | Field | 32-NGQF | 0.6786 | (9/9) | ++ | |
| 61-NIGV | 0.7400 | (9/9) | ++ | |||
| 67-NPTF | 0.5110 | (6/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6818 | (9/9) | ++ | Sequon ASN-XAA-SER/THR b | ||
| 90-NTYI | 0.6771 | (9/9) | ++ | |||
| 97-NGGV | 0.6619 | (9/9) | ++ | |||
| 117-NNSS | 0.5109 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4378 | (7/9) | - | |||
| 133-NGNP | 0.1234 | (9/9) | --- | |||
| 135-NPHM | 0.5493 | (7/9) | + | PRO-X1 | ||
| 143-NPVP | 0.1229 | (9/9) | --- | |||
| 148-NIKM | 0.6001 | (7/9) | + | |||
| 18-0430 | Field | 32-NGQF | 0.6786 | (9/9) | ++ | |
| 61-NIGV | 0.7399 | (9/9) | ++ | |||
| 67-NPTF | 0.5111 | (6/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6817 | (9/9) | ++ | Sequon ASN-XAA-SER/THR b | ||
| 89-NNTY | 0.6220 | (8/9) | + | |||
| 90-NTYI | 0.6122 | (8/9) | + | |||
| 97-NGGV | 0.6647 | (9/9) | ++ | |||
| 117-NNSS | 0.5111 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4379 | (7/9) | - | |||
| 133-NGNP | 0.1278 | (9/9) | --- | |||
| 135-NPHI | 0.5884 | (6/9) | + | PRO-X1 | ||
| 143-NPVP | 0.1086 | (9/9) | --- | |||
| 148-NIKM | 0.5840 | (8/9) | + | |||
| 17-0495 18-0374 | Field Field | 32-NGQF | 0.6786 | (9/9) | ++ | |
| 61-NIGV | 0.7398 | (9/9) | ++ | |||
| 67-NPTF | 0.5110 | (6/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6818 | (9/9) | ++ | Sequon ASN-XAA-SER/THR b | ||
| 89-NNTY | 0.6219 | (8/9) | + | |||
| 90-NTYI | 0.6120 | (8/9) | + | |||
| 97-NGGV | 0.6647 | (9/9) | ++ | |||
| 117-NNSS | 0.5108 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4378 | (7/9) | - | |||
| 133-NDNP | 0.1003 | (9/9) | --- | |||
| 135-NPHM | 0.5519 | (7/9) | + | PRO-X1 | ||
| 143-NPVP | 0.1249 | (9/9) | --- | |||
| 148-NIKM | 0.6000 | (7/9) | + | |||
| 17-0699 18-0723 18-0665 | Field Field Field | 32-NGQF | 0.6786 | (9/9) | ++ | |
| 61-NIGV | 0.7398 | (9/9) | ++ | |||
| 67-NPTF | 0.4794 | (4/9) | - | Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.7094 | (9/9) | ++ | |||
| 89-NNTY | 0.6257 | (8/9) | + | |||
| 90-NTYI | 0.6410 | (8/9) | + | |||
| 97-NGGV | 0.6649 | (9/9) | ++ | |||
| 117-NNSS | 0.5111 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4378 | (7/9) | - | |||
| 133-NGNP | 0.1234 | (9/9) | --- | |||
| 135-NPHM | 0.5494 | (7/9) | + | PRO-X1 | ||
| 143-NPVP | 0.1229 | (9/9) | --- | |||
| 148-NIKM | 0.5999 | (7/9) | + | |||
| Virulent-1-US-VA-2005 | Field | 32-NGQF | 0.6773 | (9/9) | ++ | |
| 61-NIGV | 0.7398 | (9/9) | ++ | |||
| 67-NPTF | 0.5110 | (6/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6818 | (9/9) | ++ | Sequon ASN-XAA-SER/THR b | ||
| 89-NNTY | 0.6219 | (8/9) | + | |||
| 90-NTYI | 0.6120 | (8/9) | + | |||
| 97-NGGV | 0.6647 | (9/9) | ++ | |||
| 117-NNSS | 0.5108 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4378 | (7/9) | - | |||
| 133-NGNP | 0.1234 | (9/9) | --- | |||
| 135-NPHM | 0.5492 | (7/9) | + | PRO-X1 | ||
| 143-NPVP | 0.1229 | (9/9) | --- | |||
| 148-NIKM | 0.6000 | (7/9) | + | |||
| Virulent-2-US-VA-2005 Virulent-3-US-VA-2005 Virulent-4-US-VA-2005 | Field Field Field | 32-NGQF | 0.6771 | (8/9) | + | |
| 61-NIGV | 0.7397 | (9/9) | ++ | |||
| 67-NPTF | 0.5108 | (6/9) | + | PRO-X1 a. Sequon ASN-XAA-SER/THR b | ||
| 73-NKSI | 0.6817 | (9/9) | ++ | Sequon ASN-XAA-SER/THR b | ||
| 89-NNTY | 0.6219 | (8/9) | + | |||
| 90-NTYI | 0.6121 | (8/9) | + | |||
| 97-NGGV | 0.6646 | (9/9) | ++ | |||
| 117-NNSS | 0.5107 | (5/9) | + | Sequon ASN-XAA-SER/THR | ||
| 118-NSSF | 0.4378 | (7/9) | - | |||
| 133-NGNP | 0.1234 | (9/9) | --- | |||
| 135-NPHM | 0.5591 | (7/9) | + | PRO-X1 | ||
| 143-NHVP | 0.1038 | (9/9) | --- | |||
| 148-NIKM | 0.5943 | (6/9) | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomino-Tapia, V.; Mitevski, D.; Inglis, T.; van der Meer, F.; Abdul-Careem, M.F. Molecular Characterization of Hemorrhagic Enteritis Virus (HEV) Obtained from Clinical Samples in Western Canada 2017–2018. Viruses 2020, 12, 941. https://doi.org/10.3390/v12090941
Palomino-Tapia V, Mitevski D, Inglis T, van der Meer F, Abdul-Careem MF. Molecular Characterization of Hemorrhagic Enteritis Virus (HEV) Obtained from Clinical Samples in Western Canada 2017–2018. Viruses. 2020; 12(9):941. https://doi.org/10.3390/v12090941
Chicago/Turabian StylePalomino-Tapia, Victor, Darko Mitevski, Tom Inglis, Frank van der Meer, and Mohamed Faizal Abdul-Careem. 2020. "Molecular Characterization of Hemorrhagic Enteritis Virus (HEV) Obtained from Clinical Samples in Western Canada 2017–2018" Viruses 12, no. 9: 941. https://doi.org/10.3390/v12090941
APA StylePalomino-Tapia, V., Mitevski, D., Inglis, T., van der Meer, F., & Abdul-Careem, M. F. (2020). Molecular Characterization of Hemorrhagic Enteritis Virus (HEV) Obtained from Clinical Samples in Western Canada 2017–2018. Viruses, 12(9), 941. https://doi.org/10.3390/v12090941