Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features


Abstract
1. Introduction
2. Feline Enteric Coronavirus and Feline Infectious Peritonitis Virus
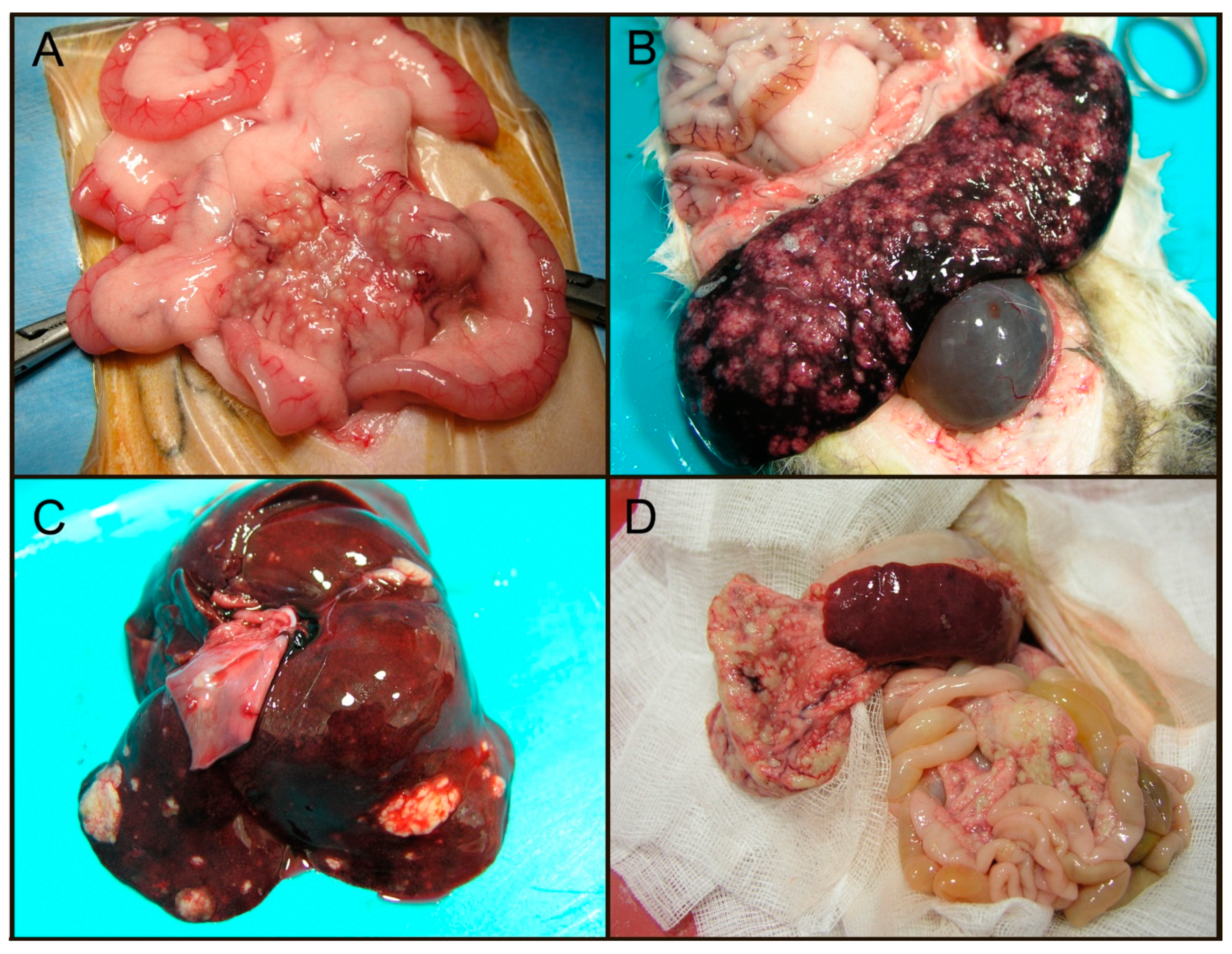
2.1. Epidemiology, Clinical and Pathologic Features
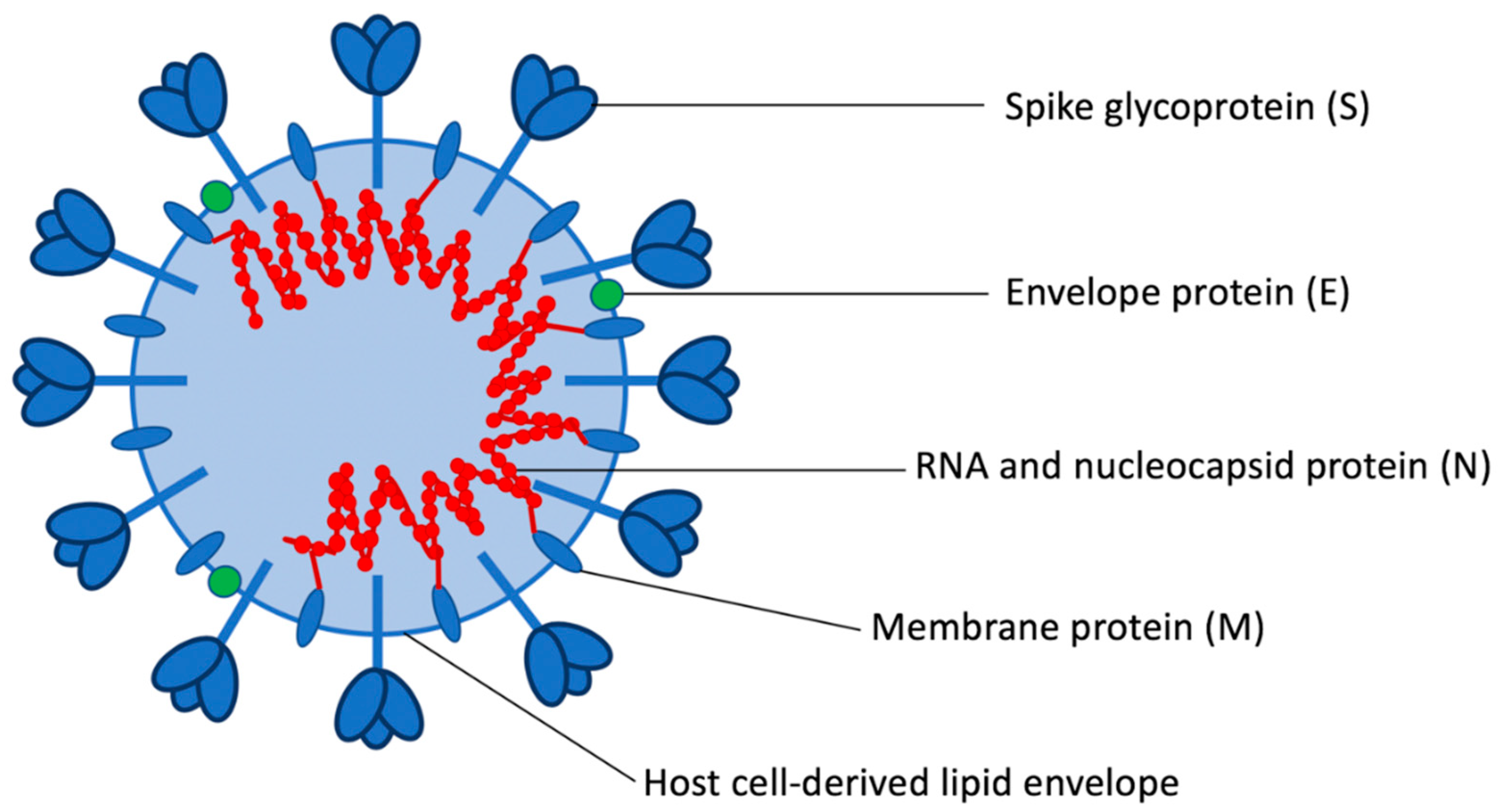
2.2. Virology
3. Ferret Enteric Coronavirus and Ferret Systemic Coronavirus
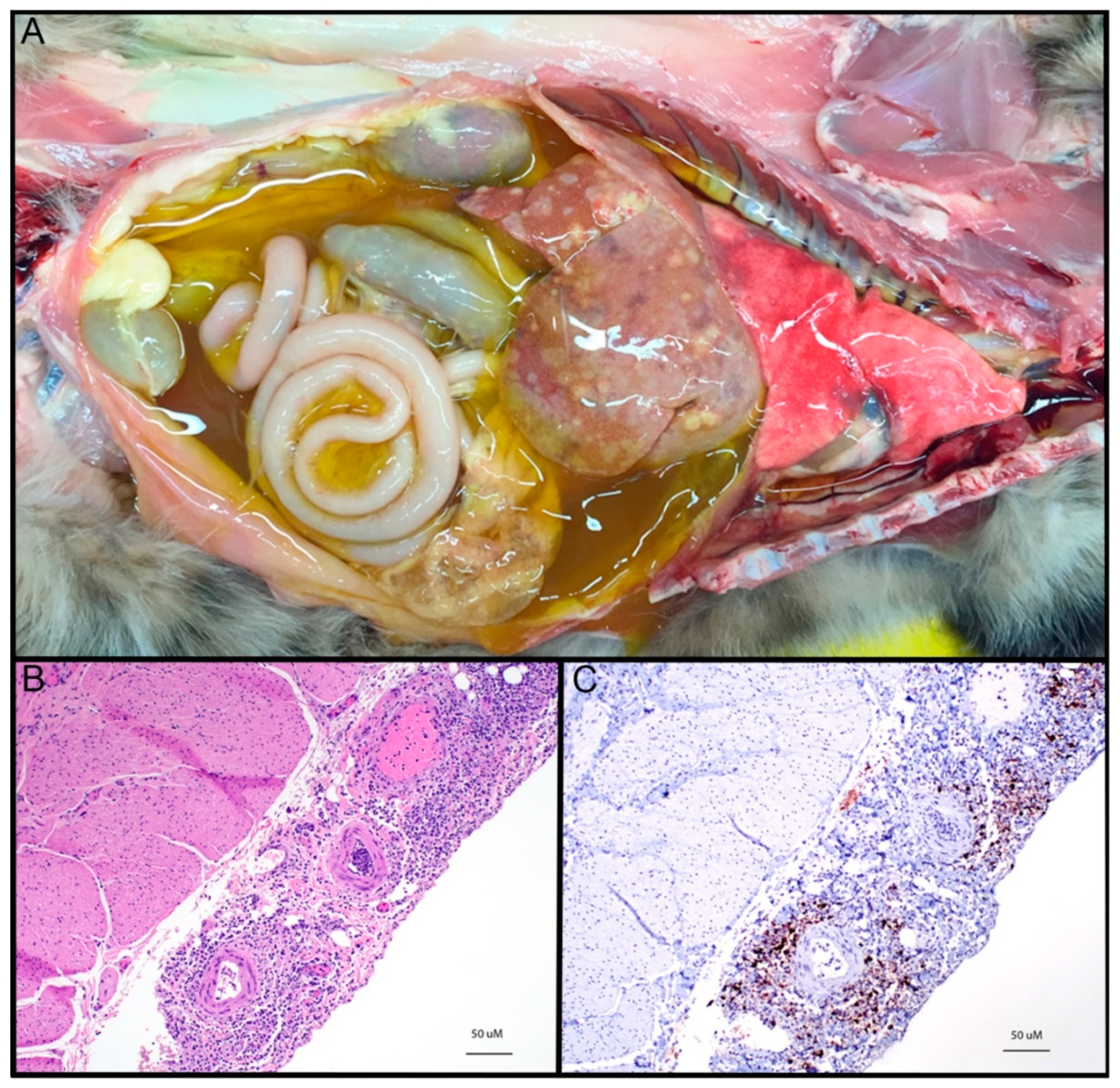
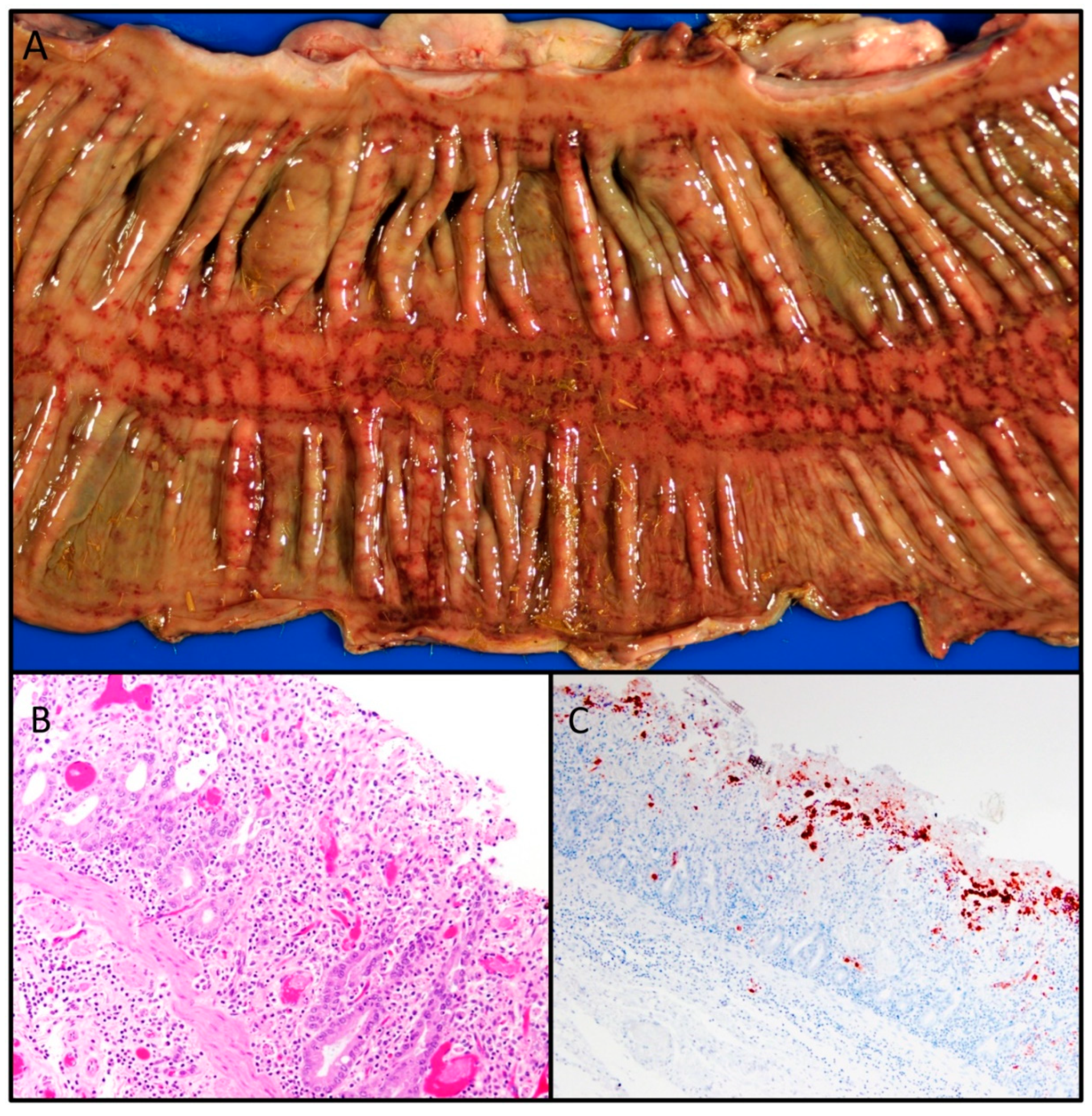
Epidemiology, Virology, and Clinical and Pathologic Features
4. Canine Enteric Coronavirus
4.1. Epidemiology and Clinical Features
4.2. Virology
4.3. Pathology
5. Canine Respiratory Coronavirus
5.1. Epidemiology and Clinical Features
5.2. Virology
5.3. Pathology
6. Equine and Alpaca Coronaviruses
6.1. Epidemiology and Clinical Features
6.2. Virology
6.3. Pathology
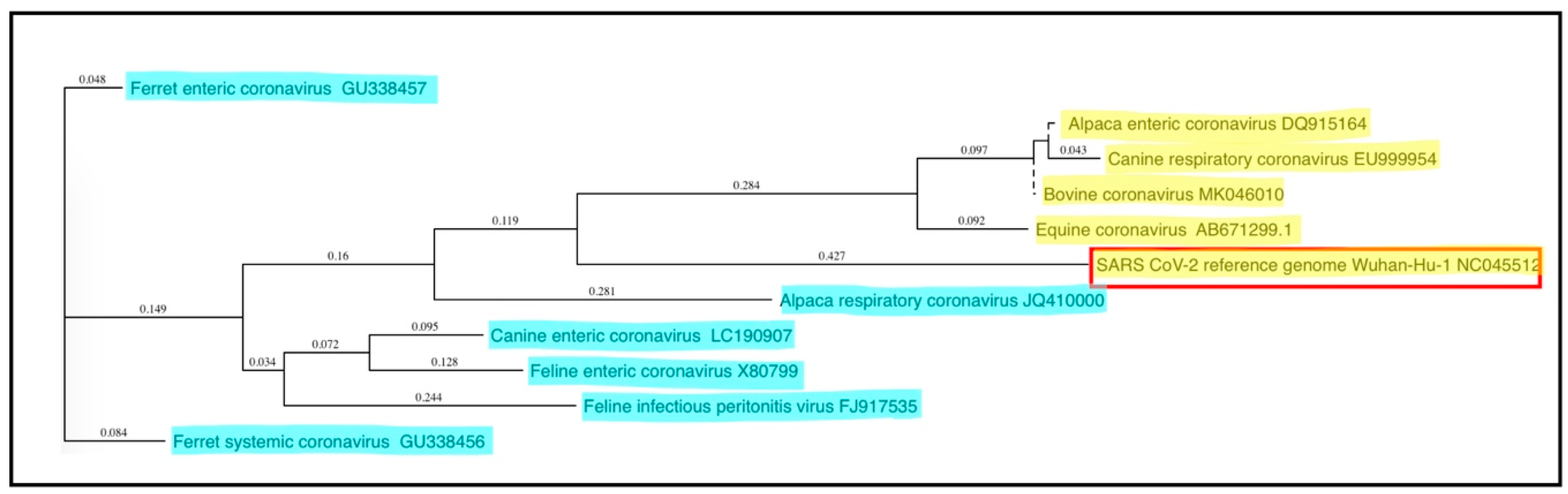
7. Zoonotic Coronaviruses
Funding
Acknowledgments
Conflicts of Interest
References
- Gnirs, K.; Quinton, J.F.; Dally, C.; Nicolier, A.; Ruel, Y. Cerebral pyogranuloma associated with systemic coronavirus infection in a ferret. J. Small Anim. Pract. 2016, 57, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Giannitti, F.; Diab, S.; Mete, A.; Stanton, J.B.; Fielding, L.; Crossley, B.; Sverlow, K.; Fish, S.; Mapes, S.; Scott, L.; et al. Necrotizing Enteritis and Hyperammonemic Encephalopathy Associated with Equine Coronavirus Infection in Equids. Vet. Pathol. 2015, 52, 1148–1156. [Google Scholar] [CrossRef]
- Kipar, A.; Kremendahl, J.; Addie, D.D.; Leukert, W.; Grant, C.K.; Reinacher, M. Fatal enteritis associated with coronavirus infection in cats. J. Comp. Pathol. 1998, 119, 1–14. [Google Scholar] [CrossRef]
- Mitchell, J.A.; Brooks, H.W.; Szladovits, B.; Erles, K.; Gibbons, R.; Shields, S.; Brownlie, J. Tropism and pathological findings associated with canine respiratory coronavirus (CRCoV). Vet. Microbiol. 2013, 162, 582–594. [Google Scholar] [CrossRef]
- Coronaviridae. In Fenner’s Veterinary Virology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 435–461.
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Lorusso, A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet. Microbiol. 2020, 244, 108693. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, K.; Shi, Z.-L.; Zhou, P. Bat Coronaviruses in China. Viruses 2019, 11, 210. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS–coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of Coronavirus Cell Entry Mediated by the Viral Spike Protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef]
- Freundt, E.C.; Yu, L.; Park, E.; Lenardo, M.J.; Xu, X.-N. Molecular Determinants for Subcellular Localization of the Severe Acute Respiratory Syndrome Coronavirus Open Reading Frame 3b Protein. J. Virol. 2009, 83, 6631–6640. [Google Scholar] [CrossRef]
- Zhang, X.; Kousoulas, K.G.; Storz, J. The hemagglutinin/esterase gene of human coronavirus strain OC43: Phylogenetic relationships to bovine and murine coronaviruses and influenza C virus. Virology 1992, 186, 318–323. [Google Scholar] [CrossRef]
- Zeng, Q.; Langereis, M.A.; van Vliet, A.L.W.; Huizinga, E.G.; de Groot, R.J. Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 9065–9069. [Google Scholar] [CrossRef] [PubMed]
- De Groot, R.J. Structure, function and evolution of the hemagglutinin-esterase proteins of corona- and toroviruses. Glycoconj. J. 2006, 23, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, I.N.A.; de Vries, R.P.; Gröne, A.; de Haan, C.A.M.; Verheije, M.H. Binding of avian coronavirus spike proteins to host factors reflects virus tropism and pathogenicity. J. Virol. 2011, 85, 8903–8912. [Google Scholar] [CrossRef]
- Walls, A.C.; Tortorici, M.A.; Snijder, J.; Xiong, X.; Bosch, B.-J.; Rey, F.A.; Veesler, D. Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion. Proc. Natl. Acad. Sci. USA 2017, 114, 11157–11162. [Google Scholar] [CrossRef]
- Schiller, J.J.; Kanjanahaluethai, A.; Baker, S.C. Processing of the coronavirus MHV-JHM polymerase polyprotein: Identification of precursors and proteolytic products spanning 400 kilodaltons of ORF1a. Virology 1998, 242, 288–302. [Google Scholar] [CrossRef]
- Sawicki, S.G.; Sawicki, D.L.; Siddell, S.G. A Contemporary View of Coronavirus Transcription. J. Virol. 2007, 81, 20–29. [Google Scholar] [CrossRef]
- Narayanan, K.; Huang, C.; Makino, S. Coronavirus Accessory Proteins. Nidoviruses 2008, 235–244. [Google Scholar] [CrossRef]
- Imbert, I.; Snijder, E.J.; Dimitrova, M.; Guillemot, J.-C.; Lécine, P.; Canard, B. The SARS-Coronavirus PLnc domain of nsp3 as a replication/transcription scaffolding protein. Virus Res. 2008, 133, 136–148. [Google Scholar] [CrossRef]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Smith, E.C.; Case, J.B.; Feng, J.Y.; Jordan, R.; et al. Coronavirus Susceptibility to the Antiviral Remdesivir (GS-5734) Is Mediated by the Viral Polymerase and the Proofreading Exoribonuclease. mBio 2018, 9, e00221–e03188. [Google Scholar] [CrossRef]
- Gordon, C.J.; Tchesnokov, E.P.; Feng, J.Y.; Porter, D.P.; Götte, M. The antiviral compound remdesivir potently inhibits RNA-dependent RNA polymerase from Middle East respiratory syndrome coronavirus. J. Biol. Chem. 2020, 295, 4773–4779. [Google Scholar] [CrossRef] [PubMed]
- Minskaia, E.; Hertzig, T.; Gorbalenya, A.E.; Campanacci, V.; Cambillau, C.; Canard, B.; Ziebuhr, J. Discovery of an RNA virus 3’->5’ exoribonuclease that is critically involved in coronavirus RNA synthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 5108–5113. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, S.G.; Sawicki, D.L. A New Model for Coronavirus Transcription. In Coronaviruses and Arteriviruses; Enjuanes, L., Siddell, S.G., Spaan, W., Eds.; Advances in Experimental Medicine and Biology; Springer US: Boston, MA, USA, 1998; pp. 215–219. ISBN 978-1-4615-5331-1. [Google Scholar]
- Fehr, A.R.; Perlman, S. Coronaviruses: An Overview of Their Replication and Pathogenesis. In Coronaviruses: Methods and Protocols; Maier, H.J., Bickerton, E., Britton, P., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; pp. 1–23. ISBN 978-1-4939-2438-7. [Google Scholar]
- Song, Z.; Xu, Y.; Bao, L.; Zhang, L.; Yu, P.; Qu, Y.; Zhu, H.; Zhao, W.; Han, Y.; Qin, C. From SARS to MERS, Thrusting Coronaviruses into the Spotlight. Viruses 2019, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Krueger, D.K.; Kelly, S.M.; Lewicki, D.N.; Ruffolo, R.; Gallagher, T.M. Variations in Disparate Regions of the Murine Coronavirus Spike Protein Impact the Initiation of Membrane Fusion. J. Virol. 2001, 75, 2792–2802. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Pyrc, K.; Dijkman, R.; Deng, L.; Jebbink, M.F.; Ross, H.A.; Berkhout, B.; van der Hoek, L. Mosaic Structure of Human Coronavirus NL63, One Thousand Years of Evolution. J. Mol. Biol. 2006, 364, 964–973. [Google Scholar] [CrossRef]
- Denison, M.R.; Graham, R.L.; Donaldson, E.F.; Eckerle, L.D.; Baric, R.S. Coronaviruses. RNA Biol. 2011, 8, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Meli, M.; Kipar, A.; Müller, C.; Jenal, K.; Gönczi, E.; Borel, N.; Gunn-Moore, D.; Chalmers, S.; Lin, F.; Reinacher, M.; et al. High viral loads despite absence of clinical and pathological findings in cats experimentally infected with feline coronavirus (FCoV) type I and in naturally FCoV-infected cats. J. Feline Med. Surg. 2004, 6, 69–81. [Google Scholar] [CrossRef]
- Herrewegh, A.A.; de Groot, R.J.; Cepica, A.; Egberink, H.F.; Horzinek, M.C.; Rottier, P.J. Detection of feline coronavirus RNA in feces, tissues, and body fluids of naturally infected cats by reverse transcriptase PCR. J. Clin. Microbiol. 1995, 33, 684–689. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Allen, C.E.; Lyons, L.A. Pathogenesis of feline enteric coronavirus infection. J. Feline Med. Surg. 2008, 10, 529–541. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Boyle, J.F.; Floyd, K.; Fudge, A.; Barker, J. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. Am. J. Vet. Res. 1981, 42, 368–377. [Google Scholar] [PubMed]
- Riemer, F.; Kuehner, K.A.; Ritz, S.; Sauter-Louis, C.; Hartmann, K. Clinical and laboratory features of cats with feline infectious peritonitis—A retrospective study of 231 confirmed cases (2000–2010). J. Feline Med. Surg. 2016, 18, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Worthing, K.A.; Wigney, D.I.; Dhand, N.K.; Fawcett, A.; McDonagh, P.; Malik, R.; Norris, J.M. Risk factors for feline infectious peritonitis in Australian cats. J. Feline Med. Surg. 2012, 14, 405–412. [Google Scholar] [CrossRef]
- Pesteanu-Somogyi, L.D.; Radzai, C.; Pressler, B.M. Prevalence of feline infectious peritonitis in specific cat breeds. J. Feline Med. Surg. 2006, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.E.; Pedersen, N.C. The inheritance of susceptibility to feline infectious peritonitis in purebred catteries. Feline Pract. 1996, 24, 14–22. [Google Scholar]
- Heeney, J.L.; Evermann, J.F.; McKeirnan, A.J.; Marker-Kraus, L.; Roelke, M.E.; Bush, M.; Wildt, D.E.; Meltzer, D.G.; Colly, L.; Lukas, J. Prevalence and implications of feline coronavirus infections of captive and free-ranging cheetahs (Acinonyx jubatus). J. Virol. 1990, 64, 1964–1972. [Google Scholar] [CrossRef]
- Colby, E.D.; Low, R.J. Feline infectious peritonitis. Vet. Med. Small Anim. Clin. 1970, 65, 783–786. [Google Scholar]
- Stephenson, N.; Swift, P.; Moeller, R.B.; Worth, S.J.; Foley, J. Feline Infectious Peritonitis in a Mountain Lion (Puma concolor), California, USA. J. Wildl. Dis. 2013, 49, 408–412. [Google Scholar] [CrossRef]
- Tuch, K.; Witte, K.H.; Wüller, H. Feststellung der Felinen Infektiösen Peritonitis (FIP) bei Hauskatzen und Leoparden in Deutschland. Zentralblatt Für Veterinärmedizin Reihe B 1974, 21, 426–441. [Google Scholar] [CrossRef]
- Poelma, F.G.; Peters, J.C.; Mieog, W.H.C.; Zwart, P. Infektiöse Peritonitis bei Krakal (Felis Caracal) und Nordluchs (Felis Lynx Lynx). In Proceedings of the Erkrankungen der Zootiere, 13th International Symposium of Zoo Veterinarians, Helsinki, Finland, 2–6 June 1974; pp. 249–253. [Google Scholar]
- Juan-Salles, C.; Domingo, M.; Herraez, P.; Fernandez, A.; Segales, J.; Fernandez, J. Feline infectious peritonitis in servals (Felis serval). Vet. Rec. 1998, 143, 535–536. [Google Scholar] [CrossRef]
- Watt, N.J.; MacIntyre, N.J.; McOrist, S. An extended outbreak of infectious peritonitis in a closed colony of european wildcats (Felis silvestris). J. Comp. Pathol. 1993, 108, 73–79. [Google Scholar] [CrossRef]
- Theobald, J. Felidae. In Zoo Wild Animal Medecine, 4th ed.; Fowler, M.E., Ed.; W.B. Saunders Co: Philadelphia, Pennsylvania, 1978; pp. 614–656. [Google Scholar]
- Pedersen, N.C. A review of feline infectious peritonitis virus infection: 1963–2008. J. Feline Med. Surg. 2009, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottier, P.J.M. Spike Protein Fusion Peptide and Feline Coronavirus Virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Licitra, B.N.; Millet, J.K.; Regan, A.D.; Hamilton, B.S.; Rinaldi, V.D.; Duhamel, G.E.; Whittaker, G.R. Mutation in spike protein cleavage site and pathogenesis of feline coronavirus. Emerg. Infect. Dis. 2013, 19, 1066–1073. [Google Scholar] [CrossRef]
- Bank-Wolf, B.R.; Stallkamp, I.; Wiese, S.; Moritz, A.; Tekes, G.; Thiel, H.-J. Mutations of 3c and spike protein genes correlate with the occurrence of feline infectious peritonitis. Vet. Microbiol. 2014, 173, 177–188. [Google Scholar] [CrossRef]
- Porter, E.; Tasker, S.; Day, M.J.; Harley, R.; Kipar, A.; Siddell, S.G.; Helps, C.R. Amino acid changes in the spike protein of feline coronavirus correlate with systemic spread of virus from the intestine and not with feline infectious peritonitis. Vet. Res. 2014, 45, 49. [Google Scholar] [CrossRef]
- Felten, S.; Hartmann, K. Diagnosis of Feline Infectious Peritonitis: A Review of the Current Literature. Viruses 2019, 11, 1068. [Google Scholar] [CrossRef]
- Pedersen, N.C. An update on feline infectious peritonitis: Diagnostics and therapeutics. Vet. J. Lond. Engl. 1997 2014, 201, 133–141. [Google Scholar] [CrossRef]
- Timmann, D.; Cizinauskas, S.; Tomek, A.; Doherr, M.; Vandevelde, M.; Jaggy, A. Retrospective analysis of seizures associated with feline infectious peritonitis in cats. J. Feline Med. Surg. 2008, 10, 9–15. [Google Scholar] [CrossRef]
- Diaz, J.V.; Poma, R. Diagnosis and clinical signs of feline infectious peritonitis in the central nervous system. Can. Vet. J. 2009, 50, 1091–1093. [Google Scholar]
- Kipar, A.; Meli, M.L. Feline infectious peritonitis: Still an enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef]
- Berg, A.-L.; Ekman, K.; Belák, S.; Berg, M. Cellular composition and interferon-γ expression of the local inflammatory response in feline infectious peritonitis (FIP). Vet. Microbiol. 2005, 111, 15–23. [Google Scholar] [CrossRef]
- Kipar, A.; May, H.; Menger, S.; Weber, M.; Leukert, W.; Reinacher, M. Morphologic features and development of granulomatous vasculitis in feline infectious peritonitis. Vet. Pathol. 2005, 42, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, Y.; Alcaraz, A.; Bossong, F.J.; Collisson, E.W.; Diniz, P.P.V.P. Feline Coronavirus in Multicat Environments. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1133–1169. [Google Scholar] [CrossRef] [PubMed]
- Jacobse-Geels, H.E.; Daha, M.R.; Horzinek, M.C. Antibody, immune complexes, and complement activity fluctuations in kittens with experimentally induced feline infectious peritonitis. Am. J. Vet. Res. 1982, 43, 666–670. [Google Scholar] [PubMed]
- Paltrinieri, S.; Parodi, M.C.; Cammarata, G.; Mambretti, M. Type IV Hypersensitivity in the Pathogenesis of FIPV-Induced Lesions. J. Vet. Med. Ser. B 1998, 45, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Eggleton, P. Hypersensitivity: Immune Complex Mediated (Type III). In ELS; John Wiley & Sons Ltd.: Chichester, UK, 2001; pp. 1–9. [Google Scholar] [CrossRef]
- Petersen, N.C.; Boyle, J.F. Immunologic phenomena in the effusive form of feline infectious peritonitis. Am. J. Vet. Res. 1980, 41, 868–876. [Google Scholar] [PubMed]
- Takano, T.; Ohyama, T.; Kokumoto, A.; Satoh, R.; Hohdatsu, T. Vascular endothelial growth factor (VEGF), produced by feline infectious peritonitis (FIP) virus-infected monocytes and macrophages, induces vascular permeability and effusion in cats with FIP. Virus Res. 2011, 158, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Warrington, R.; Watson, W.; Kim, H.L.; Antonetti, F.R. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. Off. J. Can. Soc. Allergy Clin. Immunol. 2011, 7, S1. [Google Scholar] [CrossRef]
- Ziółkowska, N.; Paździor-Czapula, K.; Lewczuk, B.; Mikulska-Skupień, E.; Przybylska-Gornowicz, B.; Kwiecińska, K.; Ziółkowski, H. Feline Infectious Peritonitis: Immunohistochemical Features of Ocular Inflammation and the Distribution of Viral Antigens in Structures of the Eye. Vet. Pathol. 2017, 54, 933–944. [Google Scholar] [CrossRef]
- Foley, J.E.; Lapointe, J.; Koblik, P.; Poland, A.; Pedersen, N.C. Diagnostic Features of Clinical Neurologic Feline Infectious Peritonitis. J. Vet. Intern. Med. 1998, 12, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Crawford, A.H.; Stoll, A.L.; Sanchez-Masian, D.; Shea, A.; Michaels, J.; Fraser, A.R.; Beltran, E. Clinicopathologic Features and Magnetic Resonance Imaging Findings in 24 Cats With Histopathologically Confirmed Neurologic Feline Infectious Peritonitis. J. Vet. Intern. Med. 2017, 31, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Redford, T.; Al-Dissi, A.N. Feline infectious peritonitis in a cat presented because of papular skin lesions. Can. Vet. J. 2019, 60, 183–185. [Google Scholar]
- Declercq, J.; De Bosschere, H.; Schwarzkopf, I.; Declercq, L. Papular cutaneous lesions in a cat associated with feline infectious peritonitis. Vet. Dermatol. 2008, 19, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.J.; Silkstone, M.A.; Kipar, A.M. Cutaneous lesions associated with coronavirus-induced vasculitis in a cat with feline infectious peritonitis and concurrent feline immunodeficiency virus infection. J. Feline Med. Surg. 2005, 7, 233–236. [Google Scholar] [CrossRef]
- Andre, N.M.; Miller, A.; Whittaker, G.R. Feline Infectious Peritonitis Virus-Associated Rhinitis in a Cat. 2020. Available online: http://creativecommons.org/licenses/by-nc-nd/4.0/ (accessed on 28 July 2020).
- Foster, R.A.; Caswell, J.L.; Rinkardt, N. Chronic fibrinous and necrotic orchitis in a cat. Can. Vet. J. 1996, 37, 681–682. [Google Scholar] [PubMed]
- Sigurðardóttir, Ó.G.; Kolbjørnsen, Ø.; Lutz, H. Orchitis in a Cat Associated with Coronavirus Infection. J. Comp. Pathol. 2001, 124, 219–222. [Google Scholar] [CrossRef]
- Rota, A.; Paltrinieri, S.; Jussich, S.; Ubertalli, G.; Appino, S. Priapism in a castrated cat associated with feline infectious peritonitis. J. Feline Med. Surg. 2008, 10, 181–184. [Google Scholar] [CrossRef]
- Kitagawa, M.; Okada, M.; Sato, T.; Kanayama, K.; Sakai, T. A feline case of isolated fourth ventricle with syringomyelia suspected to be related with feline infectious peritonitis. J. Vet. Med. Sci. 2007, 69, 759–762. [Google Scholar] [CrossRef]
- Jaimes, J.A.; Millet, J.K.; Stout, A.E.; André, N.M.; Whittaker, G.R. A Tale of Two Viruses: The Distinct Spike Glycoproteins of Feline Coronaviruses. Viruses 2020, 12, 83. [Google Scholar] [CrossRef]
- Motokawa, K.; Hohdatsu, T.; Aizawa, C.; Koyama, H.; Hashimoto, H. Molecular cloning and sequence determination of the peplomer protein gene of feline infectious peritonitis virus type I. Arch. Virol. 1995, 140, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Hohdatsu, T.; Okada, S.; Ishizuka, Y.; Yamada, H.; Koyama, H. The prevalence of types I and II feline coronavirus infections in cats. J. Vet. Med. Sci. 1992, 54, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Addie, D.D.; Schaap, I.A.T.; Nicolson, L.; Jarrett, O. Persistence and transmission of natural type I feline coronavirus infection. J. Gen. Virol. 2003, 84, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Terada, Y.; Matsui, N.; Noguchi, K.; Kuwata, R.; Shimoda, H.; Soma, T.; Mochizuki, M.; Maeda, K. Emergence of Pathogenic Coronaviruses in Cats by Homologous Recombination between Feline and Canine Coronaviruses. PLoS ONE 2014, 9, e106534. [Google Scholar] [CrossRef] [PubMed]
- Herrewegh, A.A.; Smeenk, I.; Horzinek, M.C.; Rottier, P.J.; de Groot, R.J. Feline coronavirus type II strains 79-1683 and 79-1146 originate from a double recombination between feline coronavirus type I and canine coronavirus. J. Virol. 1998, 72, 4508–4514. [Google Scholar] [CrossRef]
- Lorusso, A.; Decaro, N.; Schellen, P.; Rottier, P.J.M.; Buonavoglia, C.; Haijema, B.-J.; Groot, R.J. de Gain, Preservation, and Loss of a Group 1a Coronavirus Accessory Glycoprotein. J. Virol. 2008, 82, 10312–10317. [Google Scholar] [CrossRef]
- Tresnan, D.B.; Levis, R.; Holmes, K.V. Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I. J. Virol. 1996, 70, 8669–8674. [Google Scholar] [CrossRef]
- Di Matteo, P.; Arrigoni, G.L.; Alberici, L.; Corti, A.; Gallo-Stampino, C.; Traversari, C.; Doglioni, C.; Rizzardi, G.-P. Enhanced expression of CD13 in vessels of inflammatory and neoplastic tissues. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2011, 59, 47–59. [Google Scholar] [CrossRef]
- Hohdatsu, T.; Izumiya, Y.; Yokoyama, Y.; Kida, K.; Koyama, H. Differences in virus receptor for type I and type II feline infectious peritonitis virus. Arch. Virol. 1998, 143, 839–850. [Google Scholar] [CrossRef]
- Van Hamme, E.; Desmarets, L.; Dewerchin, H.L.; Nauwynck, H.J. Intriguing interplay between feline infectious peritonitis virus and its receptors during entry in primary feline monocytes. Virus Res. 2011, 160, 32–39. [Google Scholar] [CrossRef]
- Watanabe, R.; Eckstrand, C.; Liu, H.; Pedersen, N.C. Characterization of peritoneal cells from cats with experimentally-induced feline infectious peritonitis (FIP) using RNA-seq. Vet. Res. 2018, 49, 81. [Google Scholar] [CrossRef] [PubMed]
- Olsen, C.W.; Corapi, W.V.; Ngichabe, C.K.; Baines, J.D.; Scott, F.W. Monoclonal antibodies to the spike protein of feline infectious peritonitis virus mediate antibody-dependent enhancement of infection of feline macrophages. J. Virol. 1992, 66, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Hohdatsu, T.; Yamada, M.; Tominaga, R.; Makino, K.; Kida, K.; Koyama, H. Antibody-Dependent Enhancement of Feline Infectious Peritonitis Virus Infection in Feline Alveolar Macrophages and Human Monocyte Cell Line U937 by Serum of Cats Experimentally or Naturally Infected with Feline Coronavirus. J. Vet. Med. Sci. 1998, 60, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Shang, J.; Sun, S.; Tai, W.; Chen, J.; Geng, Q.; He, L.; Chen, Y.; Wu, J.; Shi, Z.; et al. Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry. J. Virol. 2020, 94, e02015–e02019. [Google Scholar] [CrossRef] [PubMed]
- Van Hamme, E.; Dewerchin, H.L.; Cornelissen, E.; Verhasselt, B.; Nauwynck, H.J. Clathrin- and caveolae-independent entry of feline infectious peritonitis virus in monocytes depends on dynamin. J. Gen. Virol. 2008, 89, 2147–2156. [Google Scholar] [CrossRef]
- Fox, J.G.; Marini, R.P. Biology and Diseases of the Ferret; John Wiley & Sons: Hoboken, NJ, USA, 2014; ISBN 978-1-118-78273-6. [Google Scholar]
- Williams, B.H.; Kiupel, M.; West, K.H.; Raymond, J.T.; Grant, C.K.; Glickman, L.T. Coronavirus-associated epizootic catarrhal enteritis in ferrets. J. Am. Vet. Med. Assoc. 2000, 217, 526–530. [Google Scholar] [CrossRef]
- Hoefer, H.L. Gastrointestinal Diseases of Ferrets. Ferrets Rabbit. Rodents 2020, 27–38. [Google Scholar] [CrossRef]
- Wise, A.G.; Kiupel, M.; Maes, R.K. Molecular characterization of a novel coronavirus associated with epizootic catarrhal enteritis (ECE) in ferrets. Virology 2006, 349, 164–174. [Google Scholar] [CrossRef]
- Garner, M.M.; Ramsell, K.; Morera, N.; Juan-Sallés, C.; Jiménez, J.; Ardiaca, M.; Montesinos, A.; Teifke, J.P.; Löhr, C.V.; Evermann, J.F.; et al. Clinicopathologic features of a systemic coronavirus-associated disease resembling feline infectious peritonitis in the domestic ferret (Mustela putorius). Vet. Pathol. 2008, 45, 236–246. [Google Scholar] [CrossRef]
- Li, T.-C.; Yoshizaki, S.; Kataoka, M.; Doan, Y.H.; Ami, Y.; Suzaki, Y.; Nakamura, T.; Takeda, N.; Wakita, T. Determination of Ferret Enteric Coronavirus Genome in Laboratory Ferrets. Emerg. Infect. Dis. 2017, 23, 1568–1570. [Google Scholar] [CrossRef]
- Dominguez, E.; Novellas, R.; Moya, A.; Espada, Y.; Martorell, J. Abdominal radiographic and ultrasonographic findings in ferrets (Mustela putorius furo) with systemic coronavirus infection. Vet. Rec. 2011, 169, 231. [Google Scholar] [CrossRef] [PubMed]
- Autieri, C.R.; Miller, C.L.; Scott, K.E.; Kilgore, A.; Papscoe, V.A.; Garner, M.M.; Haupt, J.L.; Bakthavatchalu, V.; Muthupalani, S.; Fox, J.G. Systemic Coronaviral Disease in 5 Ferrets. Comp. Med. 2015, 65, 508–516. [Google Scholar] [PubMed]
- Doria-Torra, G.; Vidaña, B.; Ramis, A.; Amarilla, S.P.; Martínez, J. Coronavirus Infection in Ferrets: Antigen Distribution and Inflammatory Response. Vet. Pathol. 2016, 53, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Duijvestijn, M.; Mughini-Gras, L.; Schuurman, N.; Schijf, W.; Wagenaar, J.A.; Egberink, H. Enteropathogen infections in canine puppies: (Co-)occurrence, clinical relevance and risk factors. Vet. Microbiol. 2016, 195, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Dowgier, G.; Lorusso, E.; Decaro, N.; Desario, C.; Mari, V.; Lucente, M.S.; Lanave, G.; Buonavoglia, C.; Elia, G. A molecular survey for selected viral enteropathogens revealed a limited role of Canine circovirus in the development of canine acute gastroenteritis. Vet. Microbiol. 2017, 204, 54–58. [Google Scholar] [CrossRef]
- Stavisky, J.; Pinchbeck, G.L.; German, A.J.; Dawson, S.; Gaskell, R.M.; Ryvar, R.; Radford, A.D. Prevalence of canine enteric coronavirus in a cross-sectional survey of dogs presenting at veterinary practices. Vet. Microbiol. 2010, 140, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Binn, L.N.; Lazar, E.C.; Keenan, K.P.; Huxsoll, D.L.; Marchwicki, R.H.; Strano, A.J. Recovery and characterization of a coronavirus from military dogs with diarrhea. Proc. Annu. Meet. US Anim. Health Assoc. 1974, 359–366. [Google Scholar]
- Decaro, N.; Buonavoglia, C. An update on canine coronaviruses: Viral evolution and pathobiology. Vet. Microbiol. 2008, 132, 221–234. [Google Scholar] [CrossRef]
- Decaro, N.; Martella, V.; Desario, C.; Bellacicco, A.L.; Camero, M.; Manna, L.; d’Aloja, D.; Buonavoglia, C. First detection of canine parvovirus type 2c in pups with haemorrhagic enteritis in Spain. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 468–472. [Google Scholar] [CrossRef]
- Pratelli, A.; Martella, V.; Elia, G.; Tempesta, M.; Guarda, F.; Capucchio, M.T.; Carmichael, L.E.; Buonavoglia, C. Severe enteric disease in an animal shelter associated with dual infections by canine adenovirus type 1 and canine coronavirus. J. Vet. Med. B Infect. Dis. Vet. Public Health 2001, 48, 385–392. [Google Scholar] [CrossRef]
- Pratelli, A.; Tempesta, M.; Roperto, F.P.; Sagazio, P.; Carmichael, L.; Buonavoglia, C. Fatal Coronavirus Infection in Puppies following Canine Parvovirus 2b Infection. J. Vet. Diagn. Investig. 1999, 11, 550–553. [Google Scholar] [CrossRef]
- Evermann, J.F.; Abbott, J.R.; Han, S. Canine coronavirus-associated puppy mortality without evidence of concurrent canine parvovirus infection. J. Vet. Diagn. Investig. Off. Publ. Am. Assoc. Vet. Lab. Diagn. Inc. 2005, 17, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Buonavoglia, C.; Decaro, N.; Martella, V.; Elia, G.; Campolo, M.; Desario, C.; Castagnaro, M.; Tempesta, M. Canine coronavirus highly pathogenic for dogs. Emerg. Infect. Dis. 2006, 12, 492–494. [Google Scholar] [CrossRef] [PubMed]
- Alfano, F.; Fusco, G.; Mari, V.; Occhiogrosso, L.; Miletti, G.; Brunetti, R.; Galiero, G.; Desario, C.; Cirilli, M.; Decaro, N. Circulation of pantropic canine coronavirus in autochthonous and imported dogs, Italy. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Alfano, F.; Dowgier, G.; Valentino, M.P.; Galiero, G.; Tinelli, A.; Nicola, D.; Fusco, G. Identification of Pantropic Canine Coronavirus in a Wolf ( Canis lupus italicus) in Italy. J. Wildl. Dis. 2019, 55, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Lu, C. Two genotypes of Canine coronavirus simultaneously detected in the fecal samples of healthy foxes and raccoon dogs. Wei Sheng Wu Xue Bao 2005, 45, 305–308. [Google Scholar] [PubMed]
- Zarnke, R.L.; Evermann, J.; Ver Hoef, J.M.; McNay, M.E.; Boertje, R.D.; Gardner, C.L.; Adams, L.G.; Dale, B.W.; Burch, J. Serologic survey for canine coronavirus in wolves from Alaska. J. Wildl. Dis. 2001, 37, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Molnar, B.; Duchamp, C.; Möstl, K.; Diehl, P.-A.; Betschart, B. Comparative survey of canine parvovirus, canine distemper virus and canine enteric coronavirus infection in free-ranging wolves of central Italy and south-eastern France. Eur. J. Wildl. Res. 2014, 60, 613–624. [Google Scholar] [CrossRef]
- Wang, X.; Li, C.; Guo, D.; Wang, X.; Wei, S.; Geng, Y.; Wang, E.; Wang, Z.; Zhao, X.; Su, M.; et al. Co-Circulation of Canine Coronavirus I and IIa/b with High Prevalence and Genetic Diversity in Heilongjiang Province, Northeast China. PLoS ONE 2016, 11, e0146975. [Google Scholar] [CrossRef]
- Decaro, N.; Mari, V.; Campolo, M.; Lorusso, A.; Camero, M.; Elia, G.; Martella, V.; Cordioli, P.; Enjuanes, L.; Buonavoglia, C. Recombinant canine coronaviruses related to transmissible gastroenteritis virus of Swine are circulating in dogs. J. Virol. 2009, 83, 1532–1537. [Google Scholar] [CrossRef]
- Licitra, B.; Duhamel, G.; Whittaker, G. Canine Enteric Coronaviruses: Emerging Viral Pathogens with Distinct Recombinant Spike Proteins. Viruses 2014, 6, 3363–3376. [Google Scholar] [CrossRef]
- Zappulli, V.; Caliari, D.; Cavicchioli, L.; Tinelli, A.; Castagnaro, M. Systemic fatal type II coronavirus infection in a dog: Pathological findings and immunohistochemistry. Res. Vet. Sci. 2008, 84, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Erles, K.; Brownlie, J. Canine respiratory coronavirus: An emerging pathogen in the canine infectious respiratory disease complex. Vet. Clin. North Am. Small Anim. Pract. 2008, 38, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Priestnall, S.L.; Brownlie, J.; Dubovi, E.J.; Erles, K. Serological prevalence of canine respiratory coronavirus. Vet. Microbiol. 2006, 115, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Priestnall, S.L.; Pratelli, A.; Brownlie, J.; Erles, K. Serological Prevalence of Canine Respiratory Coronavirus in Southern Italy and Epidemiological Relationship with Canine Enteric Coronavirus. J. Vet. Diagn. Investig. 2007, 19, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Yachi, A.; Mochizuki, M. Survey of Dogs in Japan for Group 2 Canine Coronavirus Infection. J. Clin. Microbiol. 2006, 44, 2615–2618. [Google Scholar] [CrossRef] [PubMed]
- Knesl, O.; Allan, F.J.; Shields, S. The seroprevalence of canine respiratory coronavirus and canine influenza virus in dogs in New Zealand. N. Z. Vet. J. 2009, 57, 295–298. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Erles, K.; Toomey, C.; Brooks, H.W.; Brownlie, J. Detection of a group 2 coronavirus in dogs with canine infectious respiratory disease. Virology 2003, 310, 216–223. [Google Scholar] [CrossRef]
- Decaro, N.; Mari, V.; Larocca, V.; Losurdo, M.; Lanave, G.; Lucente, M.S.; Corrente, M.; Catella, C.; Bo, S.; Elia, G.; et al. Molecular surveillance of traditional and emerging pathogens associated with canine infectious respiratory disease. Vet. Microbiol. 2016, 192, 21–25. [Google Scholar] [CrossRef]
- Lavan, R.; Knesl, O. Prevalence of canine infectious respiratory pathogens in asymptomatic dogs presented at US animal shelters. J. Small Anim. Pract. 2015, 56, 572–576. [Google Scholar] [CrossRef]
- Priestnall, S.L.; Mitchell, J.A.; Brooks, H.W.; Brownlie, J.; Erles, K. Quantification of mRNA encoding cytokines and chemokines and assessment of ciliary function in canine tracheal epithelium during infection with canine respiratory coronavirus (CRCoV). Vet. Immunol. Immunopathol. 2009, 127, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Arsevska, E.; Priestnall, S.L.; Singleton, D.A.; Jones, P.H.; Smyth, S.; Brant, B.; Dawson, S.; Sánchez-Vizcaíno, F.; Noble, P.J.M.; Radford, A.D. Small animal disease surveillance: Respiratory disease 2017. Vet. Rec. 2018, 182, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Wensman, J.J.; Larsson, S.; van Damme, R.; Theelke, A.-K.; Hayer, J.; Malmberg, M. Evolutionary genetics of canine respiratory coronavirus and recent introduction into Swedish dogs. Infect. Genet. Evol. 2020, 82, 104290. [Google Scholar] [CrossRef] [PubMed]
- Priestnall, S.L. Canine Respiratory Coronavirus: A Naturally Occurring Model of COVID-19? Vet. Pathol. 2020, 57, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Szczepanski, A.; Owczarek, K.; Milewska, A.; Baster, Z.; Rajfur, Z.; Mitchell, J.A.; Pyrc, K. Canine respiratory coronavirus employs caveolin-1-mediated pathway for internalization to HRT-18G cells. Vet. Res. 2018, 49, 55. [Google Scholar] [CrossRef] [PubMed]
- Szczepanski, A.; Owczarek, K.; Bzowska, M.; Gula, K.; Drebot, I.; Ochman, M.; Maksym, B.; Rajfur, Z.; Mitchell, J.A.; Pyrc, K. Canine Respiratory Coronavirus, Bovine Coronavirus, and Human Coronavirus OC43: Receptors and Attachment Factors. Viruses 2019, 11, 328. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Brooks, H.; Shiu, K.-B.; Brownlie, J.; Erles, K. Development of a quantitative real-time PCR for the detection of canine respiratory coronavirus. J. Virol. Methods 2009, 155, 136–142. [Google Scholar] [CrossRef]
- Davis, E.; Rush, B.R.; Cox, J.; DeBey, B.; Kapil, S. Neonatal Enterocolitis Associated with Coronavirus Infection in a Foal: A Case Report. J. Vet. Diagn. Investig. 2000, 12, 153–156. [Google Scholar] [CrossRef]
- Pusterla, N.; Vin, R.; Leutenegger, C.M.; Mittel, L.D.; Divers, T.J. Enteric coronavirus infection in adult horses. Vet. J. 2018, 231, 13–18. [Google Scholar] [CrossRef]
- Fielding, C.L.; Higgins, J.K.; Higgins, J.C.; McIntosh, S.; Scott, E.; Giannitti, F.; Mete, A.; Pusterla, N. Disease associated with equine coronavirus infection and high case fatality rate. J. Vet. Intern. Med. 2015, 29, 307–310. [Google Scholar] [CrossRef]
- Pusterla, N.; Mapes, S.; Wademan, C.; White, A.; Ball, R.; Sapp, K.; Burns, P.; Ormond, C.; Butterworth, K.; Bartol, J.; et al. Emerging outbreaks associated with equine coronavirus in adult horses. Vet. Microbiol. 2013, 162, 228–231. [Google Scholar] [CrossRef]
- Nemoto, M.; Oue, Y.; Morita, Y.; Kanno, T.; Kinoshita, Y.; Niwa, H.; Ueno, T.; Katayama, Y.; Bannai, H.; Tsujimura, K.; et al. Experimental inoculation of equine coronavirus into Japanese draft horses. Arch. Virol. 2014, 159, 3329–3334. [Google Scholar] [CrossRef]
- Schaefer, E.; Harms, C.; Viner, M.; Barnum, S.; Pusterla, N. Investigation of an experimental infection model of equine coronavirus in adult horses. J. Vet. Intern. Med. 2018, 32, 2099–2104. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Vin, R.; Leutenegger, C.; Mittel, L.D.; Divers, T.J. Equine coronavirus: An emerging enteric virus of adult horses. Equine Vet. Educ. 2016, 28, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Cebra, C.K.; Mattson, D.E.; Baker, R.J.; Sonn, R.J.; Dearing, P.L. Potential pathogens in feces from unweaned llamas and alpacas with diarrhea. J. Am. Vet. Med. Assoc. 2003, 223, 1806–1808. [Google Scholar] [CrossRef] [PubMed]
- Crossley, B.M.; Barr, B.C.; Magdesian, K.G.; Ing, M.; Mora, D.; Jensen, D.; Loretti, A.P.; McConnell, T.; Mock, R. Identification of a Novel Coronavirus Possibly Associated with Acute Respiratory Syndrome in Alpacas (Vicugna Pacos) in California, 2007. J. Vet. Diagn. Investig. 2010, 22, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.K.; Rodriguez, J.S.; Tibary, A. Disorders and Diseases of Pregnancy. Llama Alpaca Care 2014, 256–273. [Google Scholar] [CrossRef]
- Zhang, J.; Guy, J.S.; Snijder, E.J.; Denniston, D.A.; Timoney, P.J.; Balasuriya, U.B.R. Genomic characterization of equine coronavirus. Virology 2007, 369, 92–104. [Google Scholar] [CrossRef]
- Nemoto, M.; Oue, Y.; Murakami, S.; Kanno, T.; Bannai, H.; Tsujimura, K.; Yamanaka, T.; Kondo, T. Complete genome analysis of equine coronavirus isolated in Japan. Arch. Virol. 2015, 160, 2903–2906. [Google Scholar] [CrossRef]
- Genova, S.G.; Streeter, R.N.; Simpson, K.M.; Kapil, S. Detection of an Antigenic Group 2 Coronavirus in an Adult Alpaca with Enteritis. Clin. Vaccine Immunol. CVI 2008, 15, 1629–1632. [Google Scholar] [CrossRef]
- Jin, L.; Cebra, C.K.; Baker, R.J.; Mattson, D.E.; Cohen, S.A.; Alvarado, D.E.; Rohrmann, G.F. Analysis of the genome sequence of an alpaca coronavirus. Virology 2007, 365, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and Sources of Endemic Human Coronaviruses. Adv. Virus Res. 2018, 100, 163–188. [Google Scholar] [CrossRef]
- Crossley, B.; Mock, R.; Callison, S.; Hietala, S. Identification and Characterization of a Novel Alpaca Respiratory Coronavirus Most Closely Related to the Human Coronavirus 229E. Viruses 2012, 4, 3689–3700. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M.C.; Cavanagh, D. The Molecular Biology of Coronaviruses. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Shatkin, A.J., Eds.; Academic Press: Cambridge, MA, USA, 1997; Volume 48, pp. 1–100. [Google Scholar]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Primary Target Organ(s) | Primary Host Cell | Disease/Symptoms | Transmission | Cellular Receptor |
|---|---|---|---|---|---|
| Genus Alphacoronavirus | |||||
| Feline enteric coronavirus (FECV) | GI tract | Enterocyte | Asymptomatic to mild gastroenteritis and diarrhea | Direct contact; fecal-oral, maternal shedding | Serotype I: unknown Serotype II: APN |
| Feline infectious peritonitis virus (FIPV) | Omentum, serosal/pleural surfaces, liver, kidneys, lymph nodes, eyes, brain | Monocyte, macrophage | Peritonitis, thoracic and abdominal effusions, CNS and ocular signs. | Rare to no horizontal transmission | Serotype I: unknown Serotype II: APN |
| Ferret enteric coronavirus (FRECV) | GI tract | Enterocyte | Epizootic catarrhal enteritis | Fecal-oral | Unknown |
| Ferret systemic coronavirus (FRSCV) | Spleen, mesenteric lymph nodes, intestines, kidneys, liver, lungs, brain | Unknown | Weight loss, anorexia, diarrhea, abdominal granulomas/masses, CNS signs | Unknown | Unknown |
| Canine enteric coronavirus (CECoV) | GI tract | Enterocyte | Mild gastroenteritis and diarrhea; rarely, severe enteritis and systemic signs | Fecal-oral | Serotype I: unknown Serotype II: APN |
| Alpaca respiratory coronavirus | Respiratory tract | Respiratory epithelia (presumed) | Mild to severe respiratory disease | Aerosol (presumed) | Unknown |
| Genus Betacoronavirus | |||||
| Canine respiratory coronavirus (CRCoV) | Respiratory tract | Respiratory epithelia | Mild upper respiratory disease | Aerosol | Unknown |
| Equine coronavirus (ECoV) | GI tract | Enterocyte | Fever, anorexia, lethargy; less frequently, diarrhea, colic, neurologic signs | Fecal-oral | Unknown |
| Alpaca enteric coronavirus | GI tract | Enterocyte (presumed) | Enteritis, severe diarrhea | Fecal-oral (presumed) | Unknown |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haake, C.; Cook, S.; Pusterla, N.; Murphy, B. Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features. Viruses 2020, 12, 1023. https://doi.org/10.3390/v12091023
Haake C, Cook S, Pusterla N, Murphy B. Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features. Viruses. 2020; 12(9):1023. https://doi.org/10.3390/v12091023
Chicago/Turabian StyleHaake, Christine, Sarah Cook, Nicola Pusterla, and Brian Murphy. 2020. "Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features" Viruses 12, no. 9: 1023. https://doi.org/10.3390/v12091023
APA StyleHaake, C., Cook, S., Pusterla, N., & Murphy, B. (2020). Coronavirus Infections in Companion Animals: Virology, Epidemiology, Clinical and Pathologic Features. Viruses, 12(9), 1023. https://doi.org/10.3390/v12091023