Pediatric Enteroviral Central Nervous System Infections in Bialystok, Poland: Epidemiology, Viral Types, and Drivers of Seasonal Variation

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
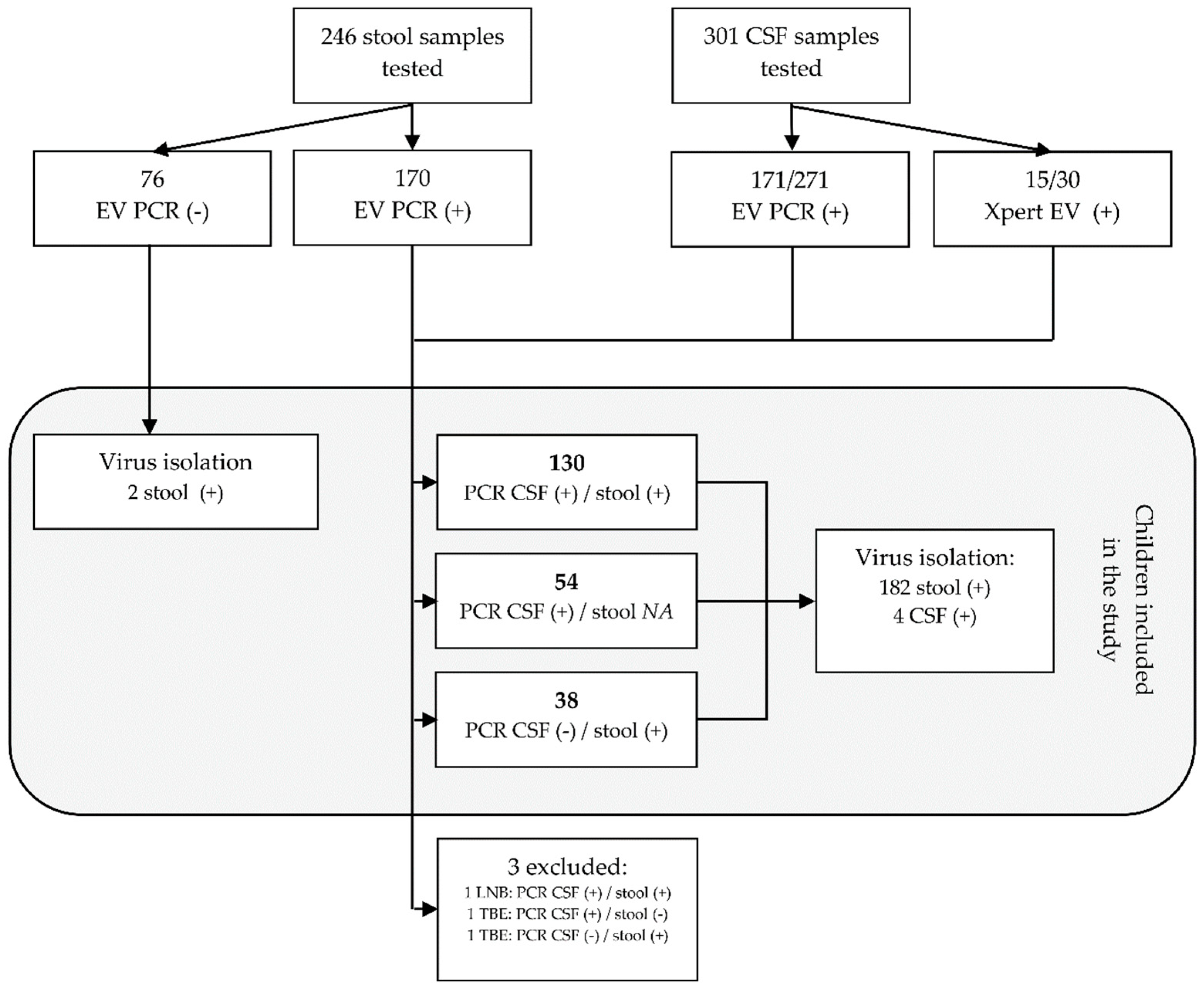
2.1. Study Design and Participants
2.2. EV Molecular Diagnosis
2.3. Virus Isolation in Cells
2.4. Enterovirus Identification
2.5. Sequence Analysis
2.6. Meteorological Data
2.7. Statistical Analysis
2.8. Ethical Considerations
3. Results
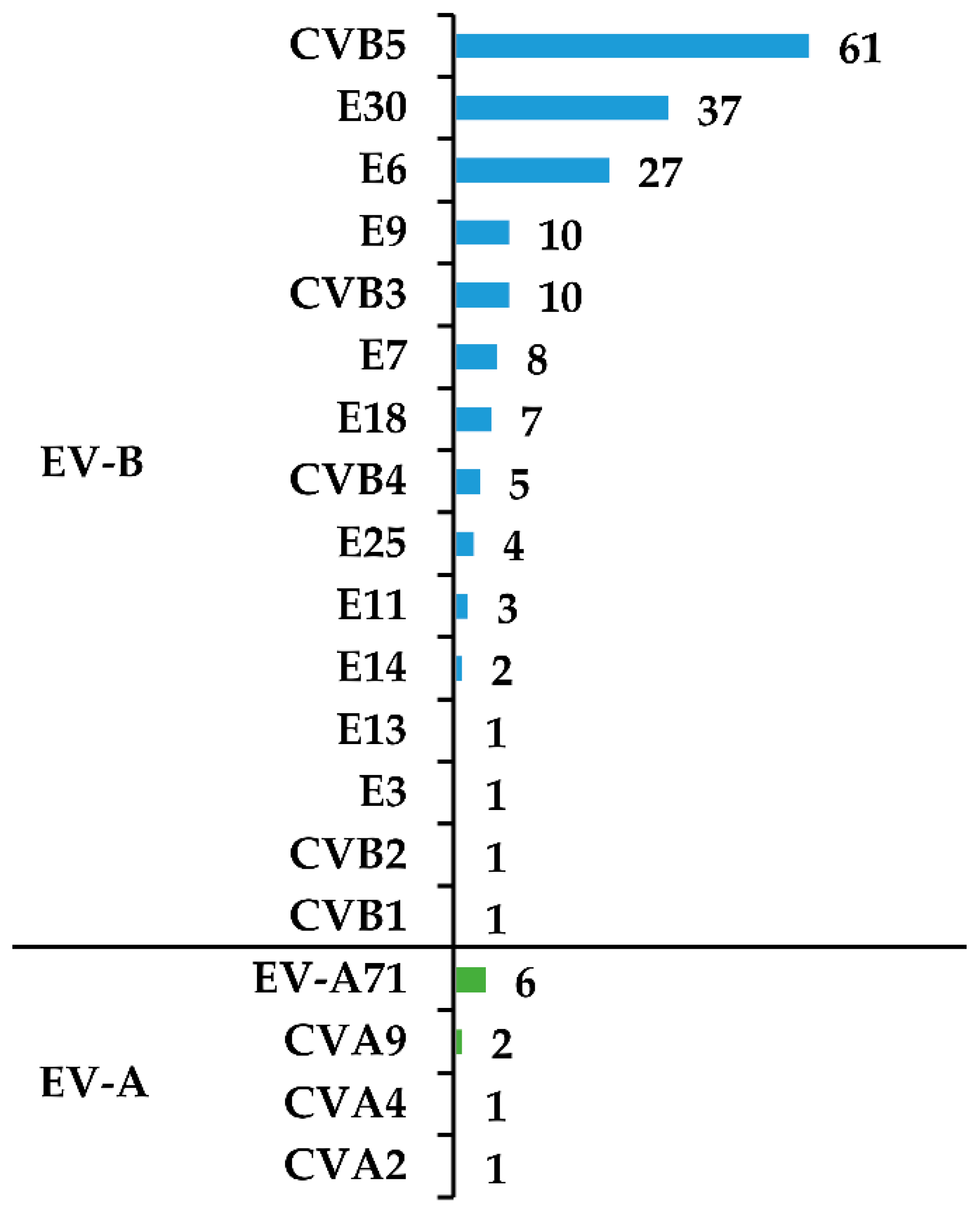
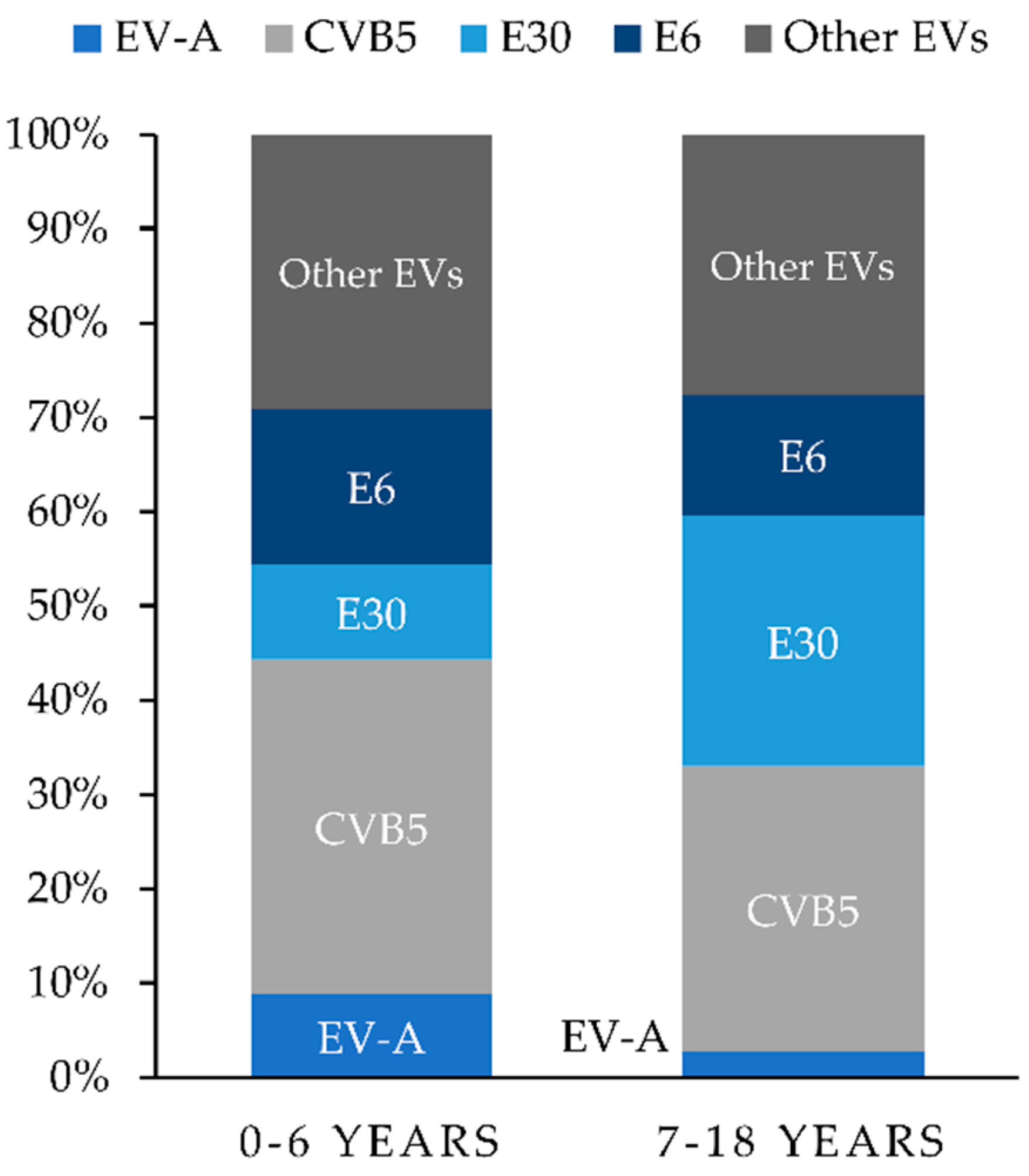
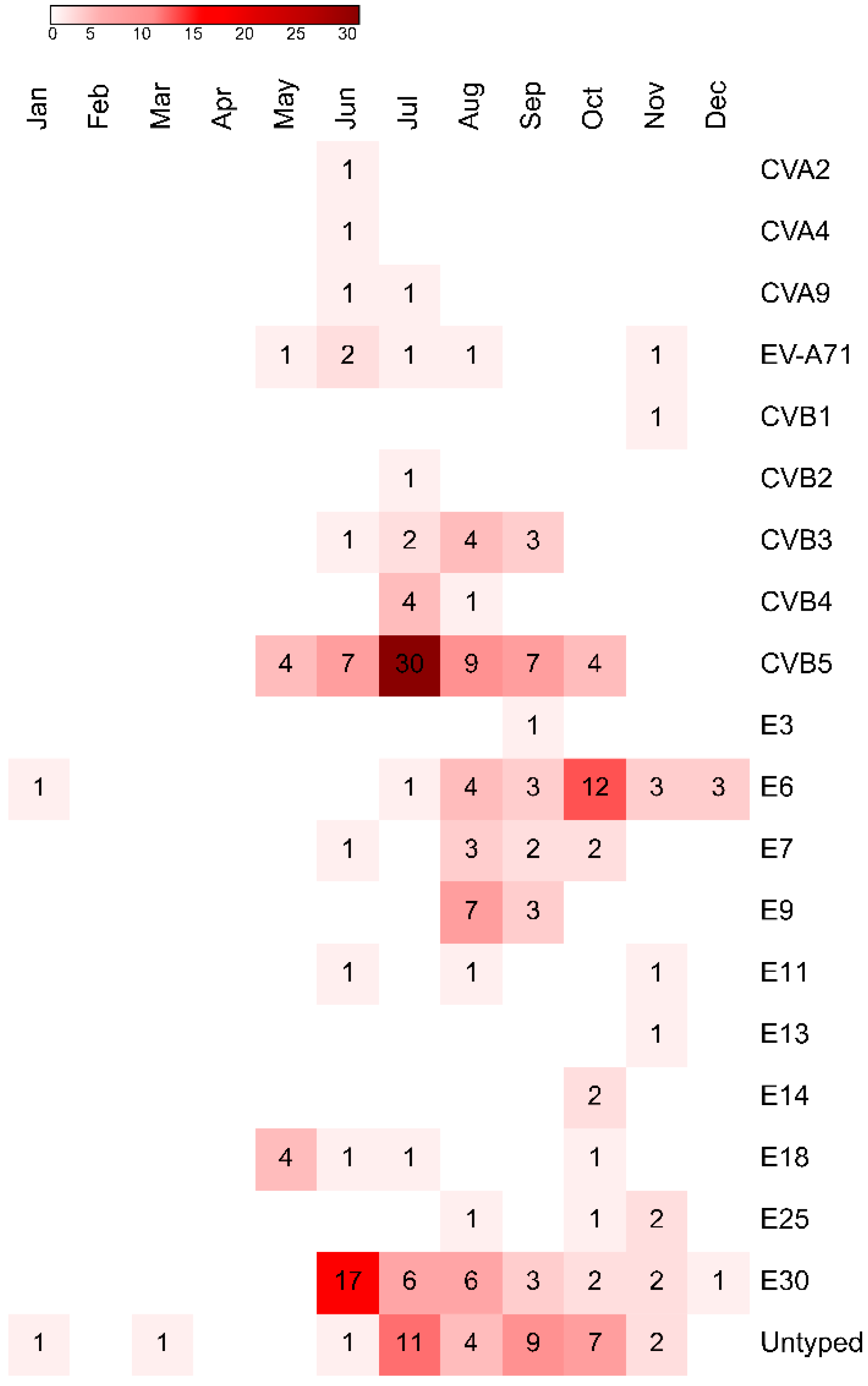
3.1. Virus Isolation and Typing
3.2. Clinical Features of the Study Population
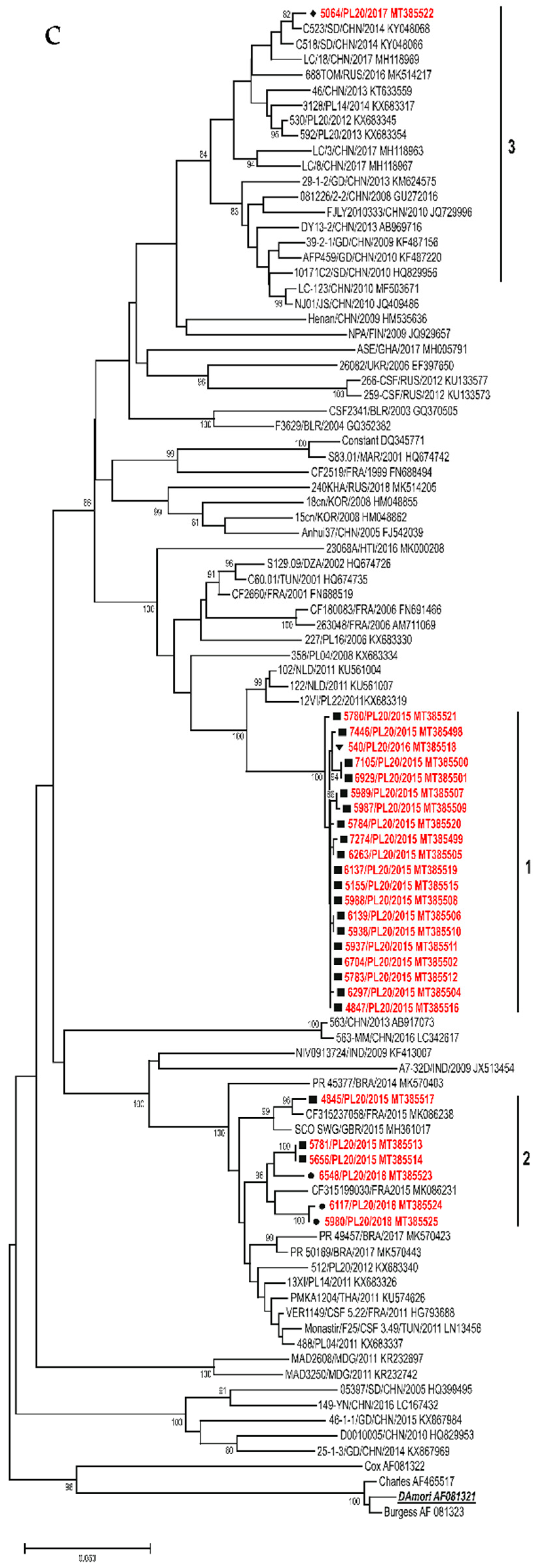
3.3. Sequence Analysis
3.3.1. E30
3.3.2. CVB5
3.3.3. E6
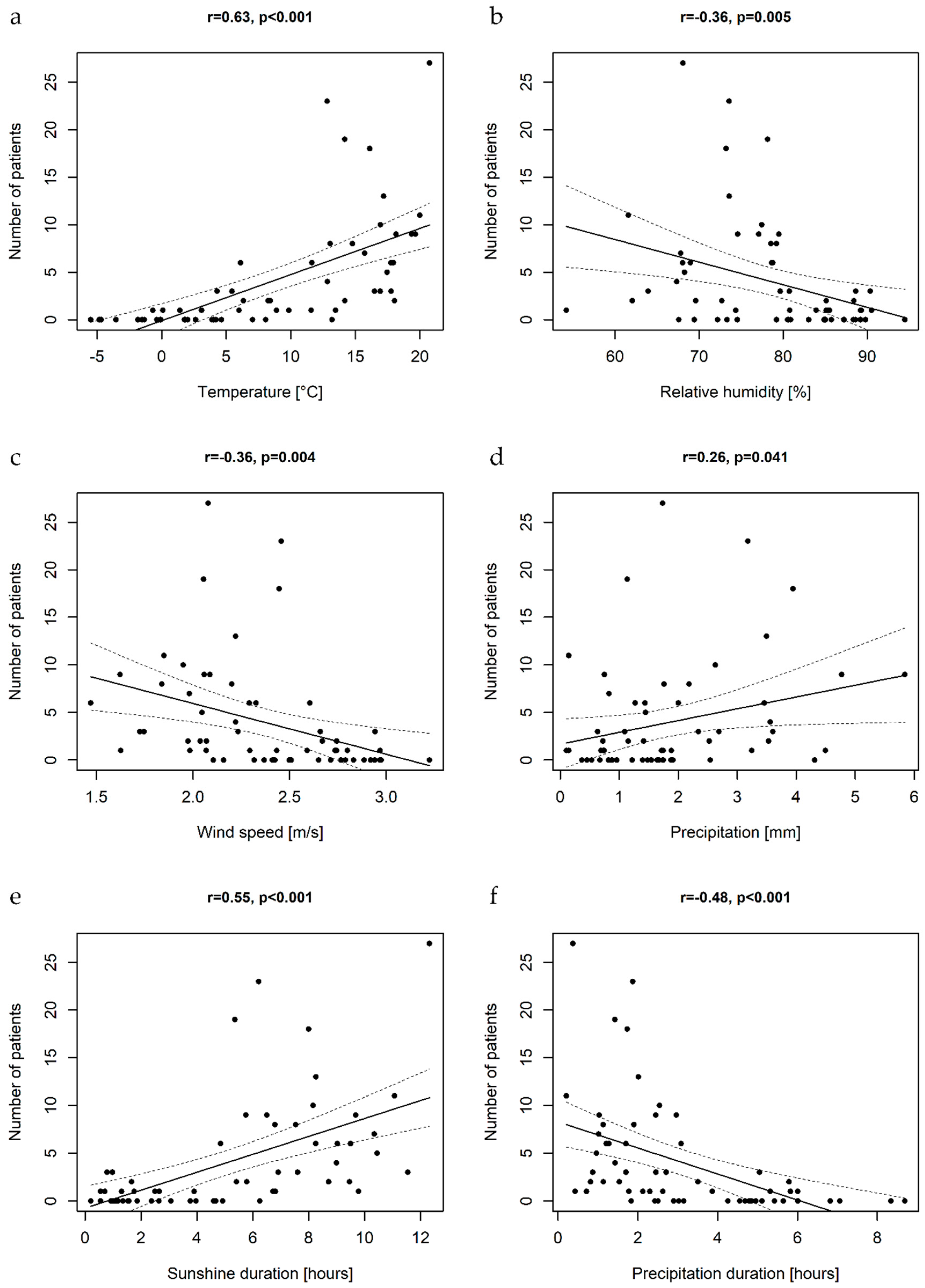
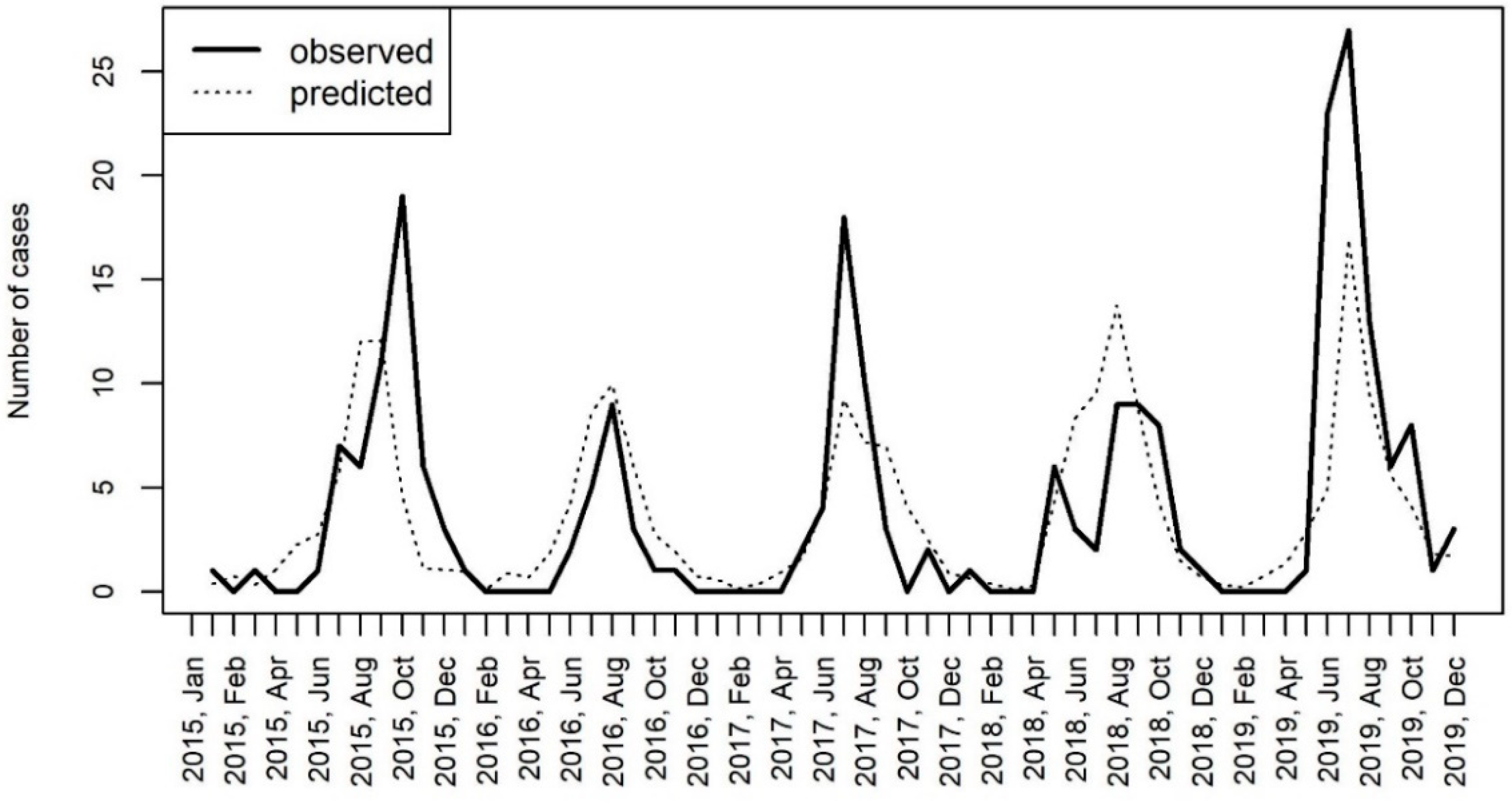
3.4. Epidemiology and Meteorological Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus taxonomy profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Khetsuriani, N.; Lamonte-Fowlkes, A.; Oberst, S.; Pallansch, M.A. Centers for disease control and prevention enterovirus surveillance—United States, 1970–2005. MMWR Surveill. Summ. 2006, 55, 1–20. [Google Scholar]
- Pons-Salort, M.; Oberste, M.S.; Pallansch, M.A.; Abedi, G.R.; Takahashi, S.; Grenfell, B.T.; Grassly, N.C. The seasonality of nonpolio enteroviruses in the United States: Patterns and drivers. Proc. Natl. Acad. Sci. USA 2018, 115, 3078–3083. [Google Scholar] [CrossRef]
- Dyer, O. Polio: WHO declares type 3 poliovirus eradicated after 31 year campaign. BMJ 2019, 367, l6201. [Google Scholar] [CrossRef] [PubMed]
- Lukashev, A.N.; Vakulenko, Y.A.; Turbabina, N.A.; Deviatkin, A.A.; Drexler, J.F. Molecular epidemiology and phylogenetics of human enteroviruses: Is there a forest behind the trees? Rev. Med. Virol. 2018, 28, e2002. [Google Scholar] [CrossRef] [PubMed]
- Suresh, S.; Rawlinson, W.D.; Andrews, P.I.; Stelzer-Braid, S. Global epidemiology of nonpolio enteroviruses causing severe neurological complications: A systematic review and meta-analysis. Rev. Med. Virol. 2020, 30, e2082. [Google Scholar] [CrossRef]
- Greninger, A.L.; Naccache, S.N.; Messacar, K.; Clayton, A.; Yu, G.; Somasekar, S.; Federman, S.; Stryke, D.; Anderson, C.; Yagi, S.; et al. A novel outbreak enterovirus D68 strain associated with acute flaccid myelitis cases in the USA (2012–2014): A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 671–682. [Google Scholar] [CrossRef]
- Toczylowski, K.; Sulik, A.; Kroten, A.; Potocka, P.; Oldak, E. Outbreak of Enteroviral Meningitis in Northeastern Poland During the Summer Season 2014. In Proceedings of the 33rd Annual Meeting of the European Society for Paediatric Infectious Diseases, Leipzig, Germany, 12–16 May 2015; Programme Book: Leipzig, Germany, 2015. [Google Scholar]
- Wieczorek, M.; Purzyńska, M.; Krzysztoszek, A.; Ciąćka, A.; Figas, A.; Szenborn, L. Genetic characterization of enterovirus A71 isolates from severe neurological cases in Poland. J. Med. Virol. 2018, 90, 372–376. [Google Scholar] [CrossRef]
- Toczylowski, K.; Bojkiewicz, E.; Barszcz, M.; Wozinska-Klepadlo, M.; Potocka, P.; Sulik, A. Etiology, clinical presentation and incidence of infectious meningitis and encephalitis in Polish children. J. Clin. Med. 2020, 9, 2324. [Google Scholar] [CrossRef]
- Stańczak, J.; Znajewska, A.; Szałtys, D. Population in Poland. Size and Structure by Territorial Division. As of June 30, 2017. In Statistical Information and Elaborations; Central Statistical Office: Warsaw, Poland, 2017. [Google Scholar]
- Albrecht, P.; Hryniewicz, W.; Kuch, A.; Przyjałkowski, W.; Skoczyńska, A.; Szenborn, L. Recommendations for the Management of Bacterial Infections of the Central Nervous System; National Medicines Institute: Warsaw, Poland, 2011; p. 38. [Google Scholar]
- Venkatesan, A.; Tunkel, A.R.; Bloch, K.C.; Lauring, A.S.; Sejvar, J.; Bitnun, A.; Stahl, J.-P.; Mailles, A.; Drebot, M.; Rupprecht, C.E.; et al. Case definitions, diagnostic algorithms, and priorities in encephalitis: Consensus statement of the international encephalitis consortium. Clin. Infect. Dis. 2013, 57, 1114–1128. [Google Scholar] [CrossRef]
- World Health Organization. Polio Laboratory Manual, 4th ed.; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Leitch, E.C.M.; Harvala, H.; Robertson, I.; Ubillos, I.; Templeton, K.; Simmonds, P. Direct identification of human enterovirus serotypes in cerebrospinal fluid by amplification and sequencing of the VP1 region. J. Clin. Virol. 2009, 44, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, M.; Krzysztoszek, A.; Figas, A. Molecular characterization of echovirus 30 isolates from Poland, 1995–2015. Virus Genes 2016, 52, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Henquell, C.; Mirand, A.; Richter, J.; Schuffenecker, I.; Böttiger, B.; Diedrich, S.; Terletskaia-Ladwig, E.; Christodoulou, C.; Peigue-Lafeuille, H.; Bailly, J.-L. Phylogenetic patterns of human coxsackievirus B5 arise from population dynamics between two genogroups and reveal evolutionary factors of molecular adaptation and transmission. J. Virol. 2013, 87, 12249–12259. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, M.; Krzysztoszek, A.; Ciąćka, A.; Figas, A. Molecular characterization of environmental and clinical echovirus 6 isolates from Poland, 2006–2014: Genetic analysis of echovirus 6 from Poland. J. Med. Virol. 2017, 89, 936–940. [Google Scholar] [CrossRef]
- Rhoades, R.E.; Tabor-Godwin, J.M.; Tsueng, G.; Feuer, R. Enterovirus infections of the central nervous system. Virology 2011, 411, 288–305. [Google Scholar] [CrossRef]
- Trallero, G.; Avellon, A.; Otero, A.; De Miguel, T.; Pérez, C.; Rabella, N.; Rubio, G.; Echevarria, J.E.; Cabrerizo, M. Enteroviruses in Spain over the decade 1998–2007: Virological and epidemiological studies. J. Clin. Virol. 2010, 47, 170–176. [Google Scholar] [CrossRef]
- Christian, K.A.; Ijaz, K.; Dowell, S.F.; Chow, C.C.; Chitale, R.A.; Bresee, J.S.; Mintz, E.; Pallansch, M.A.; Wassilak, S.; McCray, E.; et al. What we are watching—Five top global infectious disease threats, 2012: A perspective from CDC’s Global Disease Detection Operations Center. Emerg. Health Threat. J. 2013, 6, 20632. [Google Scholar] [CrossRef][Green Version]
- González-Sanz, R.; Taravillo, I.; Reina, J.; Navascués, A.; Moreno-Docón, A.; Aranzamendi, M.; Romero, M.P.; del Cuerpo, M.; Pérez-González, C.; Pérez-Castro, S.; et al. Enterovirus D68-associated respiratory and neurological illness in Spain, 2014–2018. Emerg. Microbes Infect. 2019, 8, 1438–1444. [Google Scholar] [CrossRef]
- Holmes, C.W.; Koo, S.S.F.; Osman, H.; Wilson, S.; Xerry, J.; Gallimore, C.I.; Allen, D.J.; Tang, J.W. Predominance of enterovirus B and echovirus 30 as cause of viral meningitis in a UK population. J. Clin. Virol. 2016, 81, 90–93. [Google Scholar] [CrossRef]
- Milia, M.G.; Cerutti, F.; Gregori, G.; Burdino, E.; Allice, T.; Ruggiero, T.; Proia, M.; De Rosa, G.; Enrico, E.; Lipani, F.; et al. Recent outbreak of aseptic meningitis in Italy due to Echovirus 30 and phylogenetic relationship with other European circulating strains. J. Clin. Virol. 2013, 58, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Nougairede, A.; Bessaud, M.; Thiberville, S.-D.; Piorkowski, G.; Ninove, L.; Zandotti, C.; Charrel, R.N.; Guilhem, N.; de Lamballerie, X. Widespread circulation of a new echovirus 30 variant causing aseptic meningitis and non-specific viral illness, South-East France, 2013. J. Clin. Virol. 2014, 61, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.N.; Jiang, Q.W.; Jiang, R.J.; Chen, L.; Perlin, D.S. Echovirus 30, Jiangsu Province, China. Emerg. Infect. Dis. 2005, 11, 562–567. [Google Scholar] [CrossRef]
- Benschop, K.S.; Geeraedts, F.; Beuvink, B.; Spit, S.A.; Fanoy, E.B.; Claas, E.C.; Pas, S.D.; Schuurman, R.; Verweij, J.J.; Bruisten, S.M.; et al. Increase in ECHOvirus 6 infections associated with neurological symptoms in the Netherlands, June to August 2016. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Luchs, A.; Russo, D.H.; Cilli, A.; Costa, F.F.; Morillo, S.G.; Machado, B.C.; Pellini, A.C.G.; de Cássia Compagnoli Carmona, R.; Timenetsky, M.D.C.S.T. Echovirus 6 associated to aseptic meningitis outbreak, in São Joaquim da Barra, São Paulo, Brazil. Braz. J. Microbiol. 2008, 39, 28–31. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mao, N.; Zhao, L.; Zhu, Z.; Chen, X.; Zhou, S.; Zhang, Y.; Cui, A.; Ji, Y.; Xu, S.; Xu, W. An aseptic meningitis outbreak caused by echovirus 6 in Anhui province, China. J. Med. Virol. 2010, 82, 441–445. [Google Scholar] [CrossRef]
- Siafakas, N.; Goudesidou, M.; Gaitana, K.; Gounaris, A.; Velegraki, A.; Pantelidi, K.; Zerva, L.; Petinaki, E. Successful control of an echovirus 6 meningitis outbreak in a neonatal intensive care unit in central Greece. Am. J. Infect. Control. 2013, 41, 1125–1128. [Google Scholar] [CrossRef]
- Chen, P.; Tao, Z.; Song, Y.; Liu, G.; Wang, H.; Liu, Y.; Song, L.; Li, Y.; Lin, X.; Cui, N.; et al. A coxsackievirus B5-associated aseptic meningitis outbreak in Shandong Province, China in 2009. J. Med. Virol. 2013, 85, 483–489. [Google Scholar] [CrossRef]
- Kumar, A.; Shukla, D.; Kumar, R.; Idris, M.Z.; Misra, U.K.; Dhole, T.N. An epidemic of encephalitis associated with human enterovirus B in Uttar Pradesh, India, 2008. J. Clin. Virol. 2011, 51, 142–145. [Google Scholar] [CrossRef]
- Chen, P.; Lin, X.; Liu, G.; Wang, S.; Song, L.; Tao, Z.; Xu, A. Analysis of enterovirus types in patients with symptoms of aseptic meningitis in 2014 in Shandong, China. Virology 2018, 516, 196–201. [Google Scholar] [CrossRef]
- Vignuzzi, M.; Stone, J.K.; Arnold, J.J.; Cameron, C.E.; Andino, R. Quasispecies diversity determines pathogenesis through cooperative interactions in a viral population. Nature 2006, 439, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Wilson, M.E. The role of the traveler in emerging infections and magnitude of travel. Med. Clin. N. Am. 2008, 92, 1409–1432. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Sport and Tourism. Characteristics of Domestic and Foreign Trips of Polish Residents in the First Half of 2018. Available online: https://www.msit.gov.pl/ (accessed on 30 July 2020).
- Kopecka, H.; Brown, B.; Pallansch, M. Genotypic variation in coxsackievirus B5 isolates from three different outbreaks in the United States. Virus Res. 1995, 38, 125–136. [Google Scholar] [CrossRef]
- Majer, A.; McGreevy, A.; Booth, T.F. Molecular pathogenicity of enteroviruses causing neurological disease. Front. Microbiol 2020, 11, 540. [Google Scholar] [CrossRef]
- Chang, L.-Y.; Hsia, S.-H.; Wu, C.-T.; Huang, Y.-C.; Lin, K.-L.; Fang, T.-Y.; Lin, T.-Y. Outcome of enterovirus 71 infections with or without stage-based management: 1998 to 2002. Pediatr. Infect. Dis. J. 2004, 23, 327–332. [Google Scholar] [CrossRef]
- Hsia, S.-H.; Wu, C.-T.; Chang, J.-J.; Lin, T.-Y.; Chung, H.-T.; Lin, K.-L.; Hwang, M.-S.; Chou, M.-L.; Chang, L.-Y. Predictors of unfavorable outcomes in enterovirus 71-related cardiopulmonary failure in children. Pediatr. Infect. Dis. J. 2005, 24, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Toczylowski, K.; Wojtkowska, M.; Sulik, A. Enteroviral meningitis reduces CSF concentration of Aβ42, but does not affect markers of parenchymal damage. Eur. J. Clin. Microbiol. Infect. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-W.; Chang, K.-C.; Kao, C.-M.; Chang, S.-P.; Tung, Y.-Y.; Chen, S.-H. Lymphocyte and antibody responses reduce enterovirus 71 lethality in mice by decreasing tissue viral loads. J. Virol. 2009, 83, 6477–6483. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-M.; Lei, H.-Y.; Huang, M.-C.; Su, L.-Y.; Lin, H.-C.; Yu, C.-K.; Wang, J.-L.; Liu, C.-C. Modulation of cytokine production by intravenous immunoglobulin in patients with enterovirus 71-associated brainstem encephalitis. J. Clin. Virol. 2006, 37, 47–52. [Google Scholar] [CrossRef]
- Abzug, M.J.; Keyserling, H.L.; Lee, M.L.; Levin, M.J.; Rotbart, H.A. Neonatal enterovirus infection: Virology, serology, and effects of intravenous immune globulin. Clin. Infect. Dis. 1995, 20, 1201–1206. [Google Scholar] [CrossRef]
- McKinney, R.E.; Katz, S.L.; Wilfert, C.M. Chronic enteroviral meningoencephalitis in agammaglobulinemic patients. Rev. Infect. Dis. 1987, 9, 334–356. [Google Scholar] [CrossRef] [PubMed]
- Jaijakul, S.; Salazar, L.; Wootton, S.H.; Aguilera, E.; Hasbun, R. The clinical significance of neutrophilic pleocytosis in cerebrospinal fluid in patients with viral central nervous system infections. Int. J. Infect. Dis. 2017, 59, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Cabrerizo, M.; Echevarria, J.E.; González, I.; de Miguel, T.; Trallero, G. Molecular epidemiological study of HEV-B enteroviruses involved in the increase in meningitis cases occurred in Spain during 2006. J. Med. Virol. 2008, 80, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Molet, L.; Saloum, K.; Marque-Juillet, S.; Garbarg-Chenon, A.; Henquell, C.; Schuffenecker, I.; Peigue-Lafeuille, H.; Rozenberg, F.; Mirand, A. Enterovirus infections in hospitals of Ile de France region over 2013. J. Clin. Virol. 2016, 74, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Vollbach, S.; Müller, A.; Drexler, J.F.; Simon, A.; Drosten, C.; Eis-Hübinger, A.M.; Panning, M. Prevalence, type and concentration of human enterovirus and parechovirus in cerebrospinal fluid samples of pediatric patients over a 10-year period: A retrospective study. Virol. J. 2015, 12, 199. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bubba, L.; Broberg, E.K.; Jasir, A.; Simmonds, P.; Harvala, H. Enterovirus study collaborators Circulation of non-polio enteroviruses in 24 EU and EEA countries between 2015 and 2017: A retrospective surveillance study. Lancet Infect. Dis. 2020, 20, 350–361. [Google Scholar] [CrossRef]
- Harper, G.J. Airborne micro-organisms: Survival tests with four viruses. J. Hyg. 1961, 59, 479–486. [Google Scholar] [CrossRef]
- Vareil, M.; Wille, H.; Kassab, S.; Le-cornec, C.; Puges, M.; Desclaux, A.; Lafon, M.E.; Tumiotto, C.; Cazanave, C.; Neau, D. Clinical and biological features of enteroviral meningitis among adults and children and factors associated with severity and length of stay. J. Clin. Virol. 2018, 104, 56–60. [Google Scholar] [CrossRef]
- Chung, P.W.; Huang, Y.C.; Chang, L.Y.; Lin, T.Y.; Ning, H.C. Duration of enterovirus shedding in stool. J. Microbiol. Immunol. Infect. 2001, 34, 167–170. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EV-A | EV-B | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | EVs Typed n (%) | CVA2 | CVA4 | CVA9 | EV-A71 | CVB1 | CVB2 | CVB3 | CVB4 | CVB5 | E3 | E6 | E7 | E9 | E11 | E13 | E14 | E18 | E25 | E30 |
| 2015 | 28/54 (52%) | 1 | 2 | 22 | 2 | 1 | ||||||||||||||
| 2016 | 14/22 (64%) | 1 | 1 | 1 | 2 | 8 | 1 | |||||||||||||
| 2017 | 33/35 (94%) | 1 | 1 | 1 | 4 | 2 | 18 | 1 | 1 | 2 | 1 | 1 | ||||||||
| 2018 | 38/38 (100%) | 2 | 4 | 3 | 6 | 1 | 2 | 5 | 3 | 12 | ||||||||||
| 2019 | 75/75 (100%) | 1 | 1 | 1 | 2 | 5 | 1 | 39 | 1 | 24 | ||||||||||
| Total | 188/224 (84%) | 1 | 1 | 2 | 6 | 1 | 1 | 10 | 5 | 61 | 1 | 27 | 8 | 10 | 3 | 1 | 2 | 7 | 4 | 37 |
| Enteroviruses—All | Coxsackie B5 | Echovirus 30 | Echovirus 6 | Other EV Types | |
|---|---|---|---|---|---|
| Sex | |||||
| Female; n (%) | 77 (100%) | 21 (27%) | 14 (18%) | 12 (16%) | 30 (39%) |
| Male; n (%) | 147 (100%) | 40/61 (27%) | 23 (16%) | 15 (10%) | 69 (47%) |
| Male-to-female ratio | 1.91 | 1.90 | 1.64 | 1.25 | 2.3 |
| Age (years) | 8.1 (5.4–12.7) | 8.6 (5.7–12.8) | 12.6 (8.3–15.1) 4 | 7.2 (5.4–13.7) | 7.5 (5.0–11.0) |
| Age groups | |||||
| <1; n (%) | 3 (100%) | 0 (0%) | 0 (0%) | 0 (0%) | 3 (100%) |
| 1–3; n (%) | 18 (100%) | 6 (33%) | 0 (0%) | 1 (6%) | 11 (61%) |
| 4–6; n (%) | 74 (100%) | 22 (30%) | 8 (11%) | 12 (16%) | 32 (43%) |
| 7–13; n (%) | 89 (100%) | 19 (21%) | 16 (18%) | 9 (10%) | 45 (51%) |
| 14–17; n (%) | 40 (100%) | 14 (35%) | 13 (33%) | 5 (12%) | 8 (20%) |
| Clinical presentation | |||||
| Meningitis; n (%) | 219 (100%) | 60 (27%) | 37 (17%) | 27 (12%) | 95 (44%) |
| Encephalitis; n (%) | 5 (2%) | 1 (20%) | 0 (0%) | 0 (0%) | 4 (80%) |
| Symptoms onset to CSF collection (days) | 2 (1–4) | 2 (1–4) 2 | 1 (1–2) 1,3,4 | 2 (1–6) 2 | 2 (1–4) 2 |
| Length of hospital stay (days) | 7 (7–9) | 8 (7–10) 2 | 7 (6–7) 1,4 | 7 (6–10) | 7 (7–9) 2 |
| Only stool PCR positive; n (%) | 40/224 (18%) | 21/61 (34%) 2 | 0/37 (0%) 1 | 6/27 (22%) | 13/99 (13%) |
| Signs and symptoms | |||||
| Headaches; n (%) | 221/224 (99%) | 59/61 (97%) | 37/37 (100%) | 27/27 (100%) | 98/99 (99%) |
| Headaches (days) | 2.5 (2–4) | 3 (2–5) | 2 (2–3) | 2 (1–3) | 2 (2–4) |
| Fever; n (%) | 203/224 (91%) | 61/61 (100%) | 29/37 (85%) | 23/27 (85%) | 90/99 (91%) |
| Fever (days) | 2 (1–3) | 3 (1–5) 2,3,4 | 1 (1–2) 1 | 1 (1–2) 1 | 2 (1–2) 1 |
| Vomiting; n (%) | 170/224 (76%) | 47/61 (77%) | 28/37 (76%) | 19/27 (70%) | 76/99 (77%) |
| Vomiting (days) | 1 (1–1) | 1 (1–2) | 1 (1–2) | 1 (0–1) | 1 (1–1) |
| Photophobia; n (%) | 42/224 (19%) | 5/61 (8%) | 9/37 (24%) | 7/27 (26%) | 21/99 (21%) |
| Neck stiffness; n (%) | 164/224 (73%) | 37/61 (61%) | 24/37 (65%) | 21/27 (78%) | 82/99 (83%) |
| Tremor; n (%) | 3/224 (1%) | 1/61 (2%) | 0/37 (0%) | 0/27 (0%) | 2/99 (2%) |
| Seizures; n (%) | 2/224 (<1%) | 1/61 (2%) | 0/37 (0%) | 0/27 (0%) | 1/99 (1%) |
| Altered level of consciousness; n (%) | 8/224 (4%) | 1/61 (2%) | 0/37 (0%) | 2/27 (7%) | 5/99 (5%) |
| CRP (mg/L) | 3.0 (1.0–11.0) | 1.2 (0.5–2.9) 2,3,4 | 7.7 (2.2–16.0) 1 | 6.0 (1.5–12.5) 1 | 5.2 (2.5–12.7) 1 |
| CRP > 10 mg/L; n (%) | 57/223 (26%) | 5/61 (8%) 2 | 15/36 (42%) 1 | 7/27 (26%) | 30/99 (30%) |
| ALT (IU/mL) | 11 (9–14) | 10 (9–13) 4 | 11 (9–14) | 12 (10–14) | 12 (10–14.5) 1 |
| AST (IU/mL) | 22 (17–26) | 20 (16–24) 4 | 20 (14–25) 4 | 25 (19–28) | 23.5 (19–27) 1,2 |
| WBC (×109 cells/L) | 8.8 (6.9–11.1) | 8.2 (6.9–11.0) | 8.7 (6.3–10.9) | 10.3 (7.9–13.3) | 8.7 (7.0–11.5) |
| Blood lymphocytes (%) | 28 (18–37) | 33 (25–43) 3 | 26 (18–31) | 16 (12–21) 1,4 | 27 (20–40) 4 |
| Blood neutrophils (%) | 61 (51–73) | 55 (45–63) 3 | 62 (56–71) | 78 (69–82) 1,4 | 60 (47–73) 3 |
| Blood monocytes (%) | 9 (7–12) | 9 (8–11) | 9 (7–12) | 8 (6–9) | 10 (7–12) |
| CSF protein (g/L) | 0.33 (0.25–0.44) | 0.39 (0.30–0.55) 4 | 0.31 (0.24–0.38) | 0.31 (0.28–0.40) | 0.29 (0.24–0.43) 1 |
| CSF cells (/µL) | 137 (47–364) | 249 (136–618) 2,3,4 | 73 (37–190) 1 | 124 (48–170) 1 | 86 (26–290) 1 |
| CSF lymphocytes (%) | 60 (32–80) | 73 (48–83) 2 | 46 (30–68) 1 | 55 (21–68) | 55 (30–80) |
| CSF neutrophils (%) | 26 (9–57) | 16 (6–43) 3 | 26 (14–64) | 35 (22–76) 1 | 29 (24–43) |
| CSF monocytes (%) | 7 (2–13) | 7 (3–13) | 9 (5–15) | 4 (1–10) | 7 (2–12) |
| CSF neutrophils >50%; n (%) | 62/203 (31%) | 11/58 (29%) | 12/34 (35%) | 10/24 (42%) | 29/87 (33%) |
| Univariable Analysis | Multivariable Analysis | |||
|---|---|---|---|---|
| RR (95% CI) | p | RR (95% CI) | p | |
| Temperature (°C) | 1.17 (1.15–1.2) | <0.001 | 1.17 (1.14–1.2) | <0.001 |
| Sunshine duration (hours) | 1.26 (1.21–1.31) | <0.001 | ||
| Wind speed (m/s] | 0.25 (0.18–0.34) | <0.001 | ||
| Relative humidity (%) | 0.95 (0.94–0.96) | <0.001 | ||
| Precipitation (mm) | 1.45 (1.32–1.6) | <0.001 | 1.11 (1.00–1.23) | 0.043 |
| Precipitation duration (hours) | 0.58 (0.52–0.65) | <0.001 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toczylowski, K.; Wieczorek, M.; Bojkiewicz, E.; Wietlicka-Piszcz, M.; Gad, B.; Sulik, A. Pediatric Enteroviral Central Nervous System Infections in Bialystok, Poland: Epidemiology, Viral Types, and Drivers of Seasonal Variation. Viruses 2020, 12, 893. https://doi.org/10.3390/v12080893
Toczylowski K, Wieczorek M, Bojkiewicz E, Wietlicka-Piszcz M, Gad B, Sulik A. Pediatric Enteroviral Central Nervous System Infections in Bialystok, Poland: Epidemiology, Viral Types, and Drivers of Seasonal Variation. Viruses. 2020; 12(8):893. https://doi.org/10.3390/v12080893
Chicago/Turabian StyleToczylowski, Kacper, Magdalena Wieczorek, Ewa Bojkiewicz, Magdalena Wietlicka-Piszcz, Beata Gad, and Artur Sulik. 2020. "Pediatric Enteroviral Central Nervous System Infections in Bialystok, Poland: Epidemiology, Viral Types, and Drivers of Seasonal Variation" Viruses 12, no. 8: 893. https://doi.org/10.3390/v12080893
APA StyleToczylowski, K., Wieczorek, M., Bojkiewicz, E., Wietlicka-Piszcz, M., Gad, B., & Sulik, A. (2020). Pediatric Enteroviral Central Nervous System Infections in Bialystok, Poland: Epidemiology, Viral Types, and Drivers of Seasonal Variation. Viruses, 12(8), 893. https://doi.org/10.3390/v12080893