Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Non-Persistent Transmission
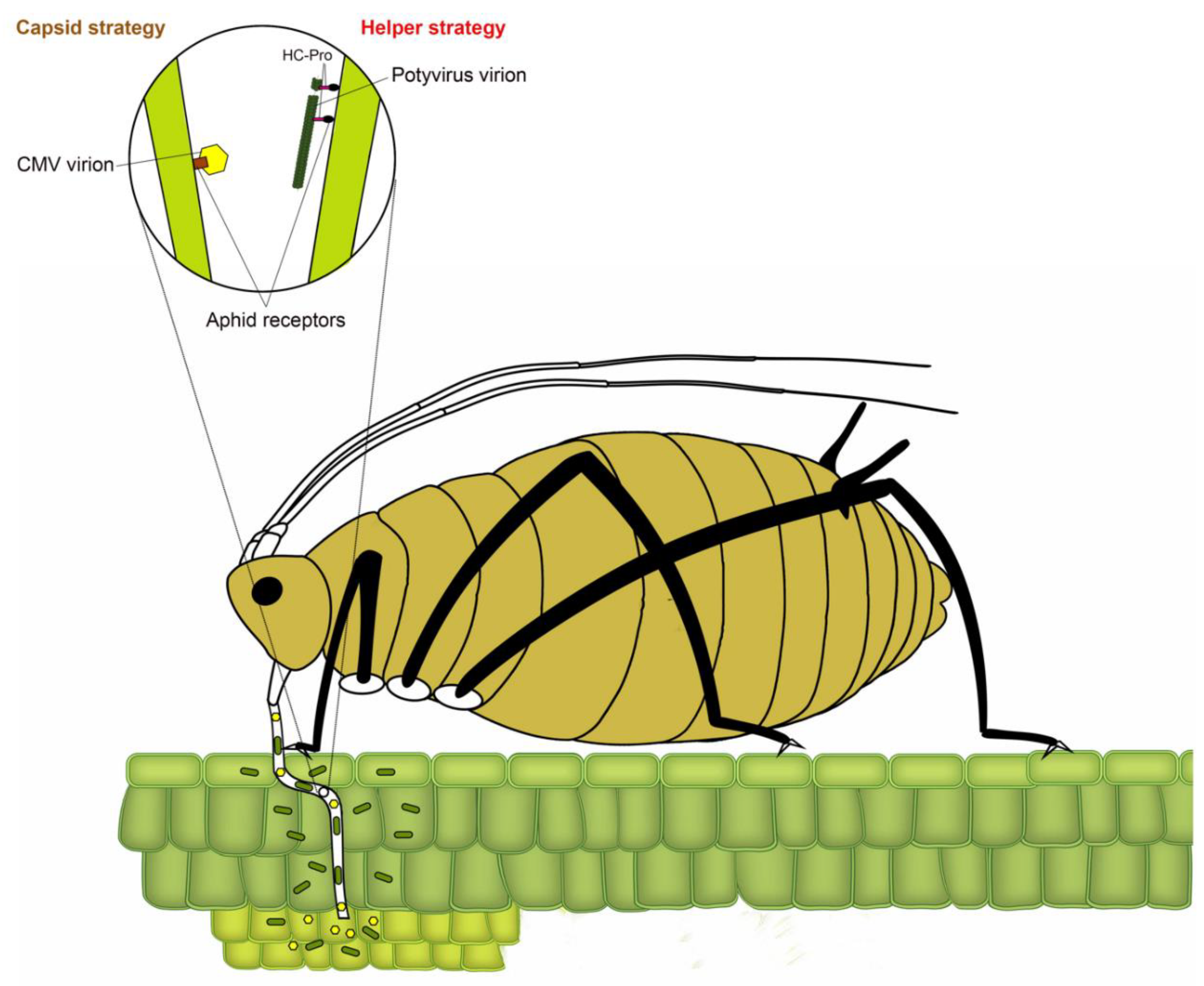
3. Binding: Viral Proteins, Sites, and Aphid Factors
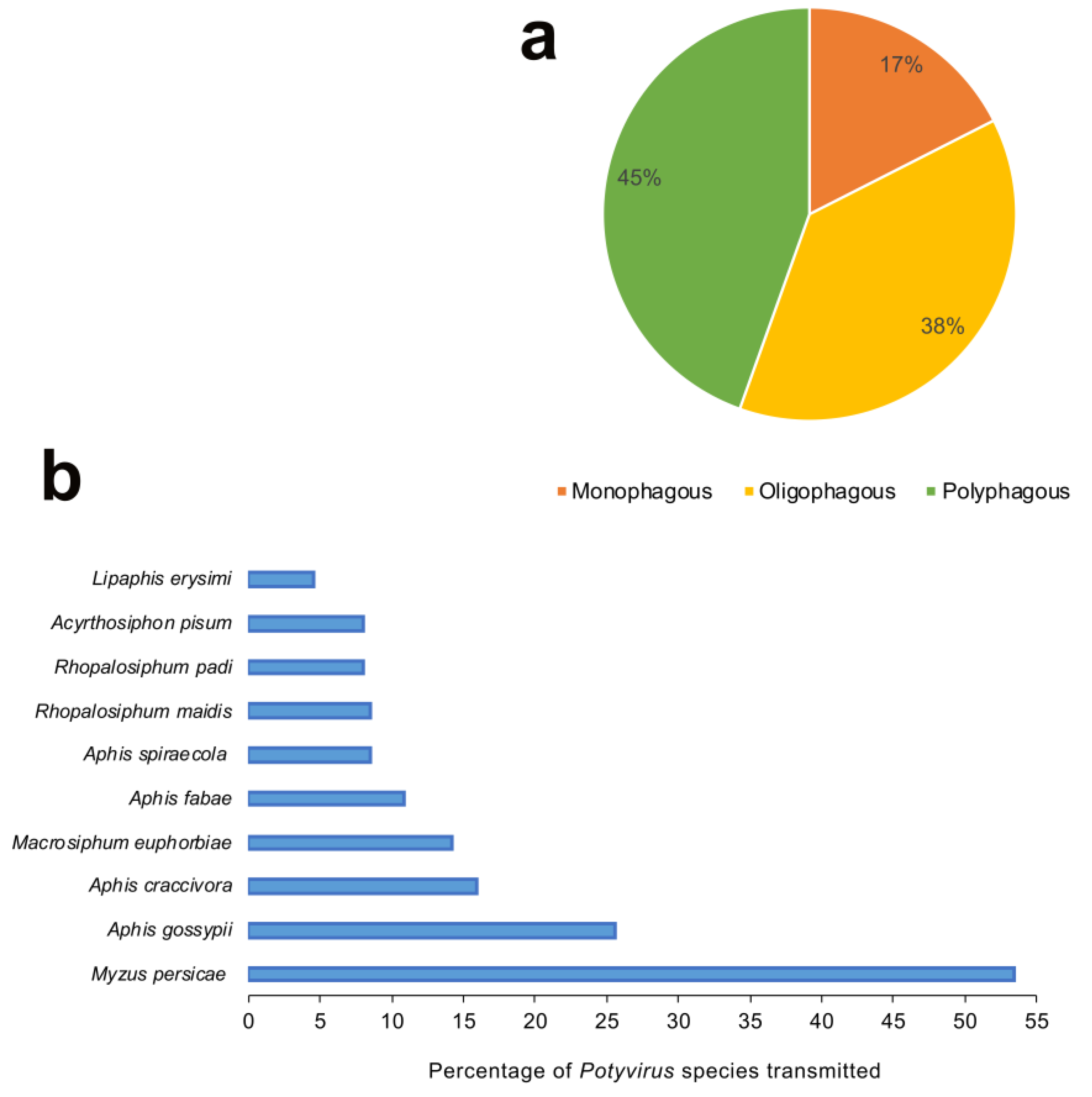
4. Aphid Biology: Behavior and Fitness
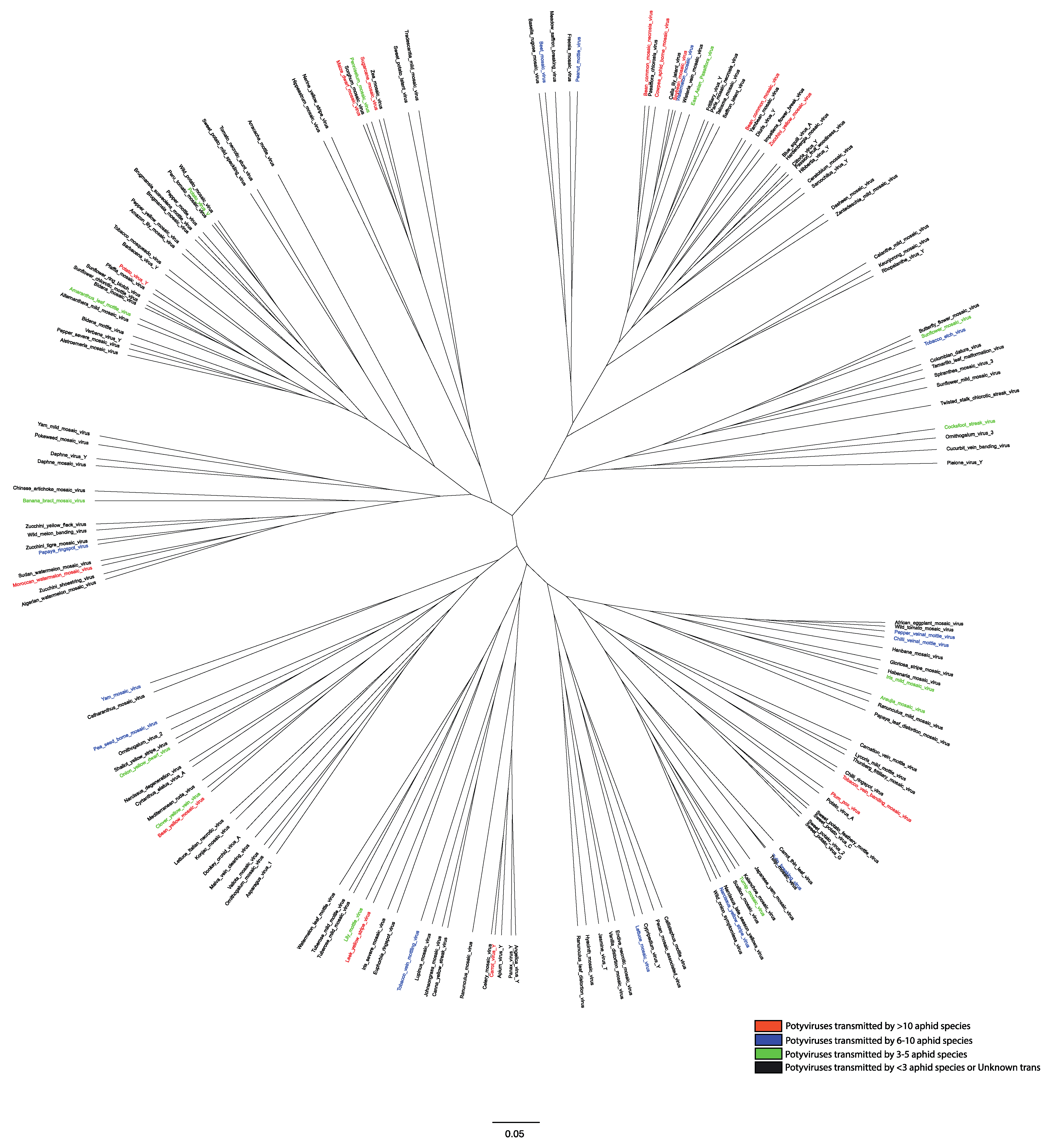
5. Virus Epidemics
6. Transmission Bottlenecks
7. Management
7.1. Current Measures
7.2. Future Directions
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harris, K.F.; Maramorosch, K. Aphids as Virus Vectors, 1st ed.; Academic Press: Cambridge, MA, USA, 1997; p. 576. ISBN 978-14-8327-388-4. [Google Scholar]
- Ng, J.C.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant. Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Lovisolo, O.; Hull, R.; Rosler, O. Coevolution of viruses with hosts and vectors and possible paleontology. Adv. Virus Res. 2003, 62, 325–379. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.S.; Day, M.F.; Eastop, V.F. A Conspectus of Aphids as Vectors of Plant. Viruses, 1st ed.; Commonwealth Institute Of Entomology: London, UK, 1962; p. 114. [Google Scholar]
- Gibbs, A.J.; Hajizadeh, M.; Ohshima, K.; Jones, R.A. The potyviruses: An evolutionary synthesis is emerging. Viruses 2020, 12, 132. [Google Scholar] [CrossRef]
- Wylie, S.J.; Adams, M.; Chalam, C.; Kreuze, J.; López-Moya, J.J.; Ohshima, K.; Praveen, S.; Rabenstein, F.; Stenger, D.; Wang, A.M.; et al. ICTV virus taxonomy profile: Potyviridae. J. Gen. Virol. 2017, 98, 352–354. [Google Scholar] [CrossRef]
- Scholthof, K.B.G.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant. Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef]
- Valli, A.; Garvia, J.A.; López-Moya, J.J. Potyviridae. In Encyclopedia of Life Sciences, 3rd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2015; pp. 1–10. ISBN 978-04-7001-590-2. [Google Scholar]
- Genus: Potyvirus; International Committee on Taxonomy of Viruses, 2020; Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/potyviridae/572/genus-potyvirus (accessed on 24 March 2020).
- Dougherty, W.G.; Carrington, J.C. Expression and function of potyviral gene products. Annu. Rev. Phytopathol. 1988, 26, 123–143. [Google Scholar] [CrossRef]
- Moury, B.; Desbiez, C. Host range evolution of potyviruses: A global phylogenetic analysis. Viruses 2020, 12, 111. [Google Scholar] [CrossRef] [PubMed]
- Sylvester, E. Circulative and propagative virus transmission by aphids. Ann. Rev. Entomol. 1980, 25, 257–286. [Google Scholar] [CrossRef]
- Ng, J.C.; Falk, B.W. Virus-vector interactions mediating nonpersistent and semipersistent transmission of plant viruses. Annu. Rev. Phytopathol. 2006, 44, 183–212. [Google Scholar] [CrossRef]
- Pirone, T.P.; Harris, K.F. Nonpersistent transmission of plant viruses by aphids. Annu. Rev. Phytopathol. 1977, 15, 55–73. [Google Scholar] [CrossRef]
- Dietzgen, R.G.; Mann, K.S.; Johnson, K.N. Plant virus–insect vector interactions: Current and potential future research directions. Viruses 2016, 8, 303. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.; Rideout, D. Comparative transmission of potato virus Y by four aphid species that infest potato. Can. J. Zool. 1953, 31, 333–341. [Google Scholar] [CrossRef]
- Boiteau, G.; Singh, R.; Parry, R.; Pelletier, Y. The spread of PVY in New brunswick potato fields: Timing and vectors. Am. Potato J. 1988, 65, 639–649. [Google Scholar] [CrossRef]
- Harrington, R.; Katis, N.; Gibson, R. Field assessment of the relative importance of different aphid species in the transmission of potato virus Y. Potato Res. 1986, 29, 67–76. [Google Scholar] [CrossRef]
- Froissart, R.; Michalakis, Y.; Blanc, S. Helper component-transcomplementation in the vector transmission of plant virus. Phytopathology 2002, 92, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Antoniw, J.F.; Beaudoin, F. Overview and analysis of the polyprotein cleavage sites in the family Potyviridae. Mol. Plant. Pathol. 2005, 6, 471–487. [Google Scholar] [CrossRef]
- Valli, A.A.; Gallo, A.; Rodamilans, B.; López-Moya, J.J.; Garcia, J.A. The HCPro from the Potyviridae family: An enviable multitasking helper component that every virus would like to have. Mol. Plant. Pathol. 2018, 19, 744–763. [Google Scholar] [CrossRef]
- Pirone, T.P.; Thornbury, D.W. Role of virion and helper component in regulating aphid trasmission of tobacco etch virus. Phytopathology 1983, 73, 872–875. [Google Scholar] [CrossRef]
- Kasschau, K.D.; Cronin, S.; Carrington, J.C. Genome amplification and long-distance movement functions associated with the central domain of tobacco etch potyvirus helper component–proteinase. Virology 1997, 228, 251–262. [Google Scholar] [CrossRef]
- Anandalakshmi, R.; Pruss, G.J.; Ge, X.; Marathe, R.; Mallory, A.C.; Smith, T.H.; Vance, V.B. A viral suppressor of gene silencing in plants. Proc. Natl. Acad. Sci. USA 1998, 95, 13079–13084. [Google Scholar] [CrossRef] [PubMed]
- Brigneti, G.; Voinnet, O.; Li, W.X.; Ji, L.H.; Ding, S.W.; Baulcombe, D.C. Retracted: Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO J. 1998, 17, 6739–6746. [Google Scholar] [CrossRef]
- Kasschau, K.D.; Carrington, J.C. A counterdefensive strategy of plant viruses: Suppression of posttranscriptional gene silencing. Cell 1998, 95, 461–470. [Google Scholar] [CrossRef]
- Nigam, D.; LaTourrette, K.; Souza, P.F.; Garcia-Ruiz, H. Genome-wide variation in potyviruses. Front. Plant. Sci. 2019, 10, 1439. [Google Scholar] [CrossRef]
- Govier, D.; Kassanis, B. A virus-induced component of plant sap needed when aphids acquire potato virus Y from purified preparations. Virology 1974, 61, 420–426. [Google Scholar] [CrossRef]
- Blanc, S.; López-Moya, J.J.; Wang, R.; García-Lampasona, S.; Thornbury, D.W.; Pirone, T.P. A specific interaction between coat protein and helper component correlates with aphid transmission of a potyvirus. Virology 1997, 231, 141–147. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Gollop, N.; Chen, S.; Chejanovsky, N.; Raccah, B. In vitro association between the helper component–proteinase of zucchini yellow mosaic virus and cuticle proteins of Myzus persicae. J. Gen. Virol. 2007, 88, 1602–1610. [Google Scholar] [CrossRef] [PubMed]
- Atreya, C.D.; Pirone, T.P. Mutational analysis of the helper component-proteinase gene of a potyvirus: Effects of amino acid substitutions, deletions, and gene replacement on virulence and aphid transmissibility. Proc. Natl. Acad. Sci. USA 1993, 90, 11919–11923. [Google Scholar] [CrossRef]
- Blanc, S.; Ammar, E.; Garcia-Lampasona, S.; Dolja, V.; Llave, C.; Baker, J.; Pirone, T. Mutations in the potyvirus helper component protein: Effects on interactions with virions and aphid stylets. J. Gen. Virol. 1998, 79, 3119–3122. [Google Scholar] [CrossRef]
- Peng, Y.; Kadoury, D.; Gal-On, A.; Huet, H.; Wang, Y.; Raccah, B. Mutations in the HC-Pro gene of zucchini yellow mosaic potyvirus: Effects on aphid transmission and binding to purified virions. J. Gen. Virol. 1998, 79, 897–904. [Google Scholar] [CrossRef]
- Huet, H.; Gal-On, A.; Meir, E.; Lecoq, H.; Raccah, B. Mutations in the helper component protease gene of zucchini yellow mosaic virus affect its ability to mediate aphid transmissibility. J. Gen. Virol. 1994, 75, 1407–1414. [Google Scholar] [CrossRef]
- López-Moya, J.; Wang, R.; Pirone, T. Context of the coat protein DAG motif affects potyvirus transmissibility by aphids. J. Gen. Virol. 1999, 80, 3281–3288. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Wu, M.; Wong, S.M. Nucleotide sequence of a Singapore isolate of zucchini yellow mosaic virus coat protein gene revealed an altered DAG motif. Virus Genes 1993, 7, 381–387. [Google Scholar] [CrossRef]
- Wylie, S.; Kueh, J.; Welsh, B.; Smith, L.; Jones, M.; Jones, R. A non-aphid-transmissible isolate of bean yellow mosaic potyvirus has an altered NAG motif in its coat protein. Arch. Virol. 2002, 147, 1813–1820. [Google Scholar] [CrossRef]
- Flasinski, S.; Cassidy, B. Potyvirus aphid transmission requires helper component and homologous coat protein for maximal efficiency. Arch. Virol. 1998, 143, 2159–2172. [Google Scholar] [CrossRef] [PubMed]
- Jossey, S.; Hobbs, H.A.; Domier, L.L. Role of soybean mosaic virus–encoded proteins in seed and aphid transmission in soybean. Phytopathology 2013, 103, 941–948. [Google Scholar] [CrossRef]
- Baulcombe, D.C.; Lloyd, J.; Manoussopoulos, I.N.; Roberts, I.M.; Harrison, B.D. Signal for potyvirus-dependent aphid transmission of potato aucuba mosaic virus and the effect of its transfer to potato virus X. J. Gen. Virol. 1993, 74, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Sako, N.; Ogata, K. Different helper factors associated with aphid transmission of some potyviruses. Virology 1981, 112, 762–765. [Google Scholar] [CrossRef]
- Lecoq, H.; Pitrat, M. Specificity of the helper-component-mediated aphid transmission of three potyviruses infecting muskmelon. Phytopathology 1985, 75, 890–893. [Google Scholar] [CrossRef]
- Ruiz-Ferrer, V.; Boskovic, J.; Alfonso, C.; Rivas, G.; Llorca, O.; López-Abella, D.; López-Moya, J.J. Structural analysis of tobacco etch potyvirus HC-Pro oligomers involved in aphid transmission. J. Virol. 2005, 79, 3758–3765. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.F.; Smith, O.P.; Duffus, J.E. Virus-Insect-Plant Interactions, 1st ed.; Academic Press: Cambridge, MA, USA, 2001; Chapter 10; pp. 181–206. [Google Scholar]
- Stenger, D.C.; Hein, G.L.; Gildow, F.E.; Horken, K.M.; French, R. Plant virus HC-Pro is a determinant of eriophyid mite transmission. J. Virol. 2005, 79, 9054–9061. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Calvino, L.; Goytia, E.; Lopez-Abella, D.; Giner, A.; Urizarna, M.; Vilaplana, L.; López-Moya, J.J. The helper-component protease transmission factor of tobacco etch potyvirus binds specifically to an aphid ribosomal protein homologous to the laminin receptor precursor. J. Gen. Virol. 2010, 91, 2862–2873. [Google Scholar] [CrossRef]
- Webster, C.G.; Pichon, E.; Van Munster, M.; Monsion, B.; Deshoux, M.; Gargani, D.; Calevro, F.; Jiménez, J.; Moreno, A.; Krenz, B. Identification of plant virus receptor candidates in the stylets of their aphid vectors. J. Gen. Virol. 2018, 92, e00432-18. [Google Scholar] [CrossRef]
- Uzest, M.; Gargani, D.; Dombrovsky, A.; Cazevieille, C.; Cot, D.; Blanc, S. The “acrostyle”: A newly described anatomical structure in aphid stylets. Arthropod Struct. Dev. 2010, 39, 221–229. [Google Scholar] [CrossRef]
- Markham, P.; Pinner, M.; Raccah, B.; Hull, R. The acquisition of a caulimovirus by different aphid species: Comparison with a potyvirus. Ann. Appl. Biol. 1987, 111, 571–587. [Google Scholar] [CrossRef]
- Blanc, S.; Uzest, M.; Drucker, M. New research horizons in vector-transmission of plant viruses. Curr. Opin. Microbiol. 2011, 14, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef]
- Bragard, C.; Caciagli, P.; Lemaire, O.; López-Moya, J.J.; MacFarlane, S.; Peters, D.; Susi, P.; Torrance, L. Status and prospects of plant virus control through interference with vector transmission. Annu. Rev. Phytopathol. 2013, 51, 177–201. [Google Scholar] [CrossRef]
- Ingwell, L.L.; Eigenbrode, S.D.; Bosque-Pérez, N.A. Plant viruses alter insect behavior to enhance their spread. Sci. Rep. 2012, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- McLean, G.D.; Garrett, R.G.; Ruesink, W.G. Plant Virus Epidemics: Monitoring, Modelling and Predicting Outbreaks, 1st ed.; Academic Press: Sydney, Australia, 1986; Chapter 1; pp. 13–33. [Google Scholar]
- Mauck, K.E.; Chesnais, Q.; Shapiro, L.R. Evolutionary determinants of host and vector manipulation by plant viruses. Adv. Virus Res. 2018, 101, 189–250. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, P.; Martin, D.P.; Elena, S.F.; Shepherd, D.N.; Roumagnac, P.; Varsani, A. Evolution and ecology of plant viruses. Nat. Rev. Microbiol. 2019, 17, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Calvino, L.; López-Abella, D.; López-Moya, J.J.; Fereres, A. Comparison of potato Virus Y and plum pox virus transmission by two aphid species in relation to their probing behavior. Phytoparasitica 2006, 34, 315. [Google Scholar] [CrossRef]
- Adachi, S.; Honma, T.; Yasaka, R.; Ohshima, K.; Tokuda, M. Effects of infection by turnip mosaic virus on the population growth of generalist and specialist aphid vectors on turnip plants. PLoS ONE 2018, 13, e0200784. [Google Scholar] [CrossRef] [PubMed]
- Gadhave, K.R.; Dutta, B.; Coolong, T.; Srinivasan, R. A non-persistent aphid-transmitted potyvirus differentially alters the vector and non-vector biology through host plant quality manipulation. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Boquel, S.; Giordanengo, P.; Ameline, A. Divergent effects of PVY-infected potato plant on aphids. Eur. J. Plant. Pathol. 2011, 129, 507–510. [Google Scholar] [CrossRef]
- Van Hoof, H. Determination of the infection pressure of potato virus YN. Neth. J. Plant. Pathol. 1977, 83, 123–127. [Google Scholar] [CrossRef]
- Singh, R.; Boiteau, G. Reevaluation of the potato aphid, Macrosiphum euphorbiae (Thomas), as vector of potato virus Y. Am. Potato J. 1986, 63, 335–340. [Google Scholar] [CrossRef]
- Raccah, B.; Gal-On, A.; Eastop, V. The role of flying aphid vectors in the transmission of cucumber mosaic virus and potato virus Y to peppers in Israel. Ann. Appl. Biol. 1985, 106, 451–460. [Google Scholar] [CrossRef]
- Harrington, R.; Gibson, R. Transmission of potato virus Y by aphids trapped in potato crops in southern England. Potato Res. 1989, 32, 167–174. [Google Scholar] [CrossRef]
- Sigvald, R. Relationship between aphid occurrence and spread of potato virus Y (PVY) in field experiments in southern Sweden. J. Appl. Entomol. 1989, 108, 35–43. [Google Scholar] [CrossRef]
- Cervantes, F.A.; Alvarez, J.M. Role of hairy nightshade in the transmission of different potato virus Y strains on Solanum tuberosum (L.). Plant. Health Prog. 2010, 11, 38. [Google Scholar] [CrossRef]
- Verbeek, M.; Piron, P.; Dullemans, A.; Cuperus, C.; Van Der Vlugt, R. Determination of aphid transmission efficiencies for N, NTN and Wilga strains of potato virus Y. Ann. Appl. Biol. 2010, 156, 39–49. [Google Scholar] [CrossRef]
- Srinivasan, R.; Hall, D.G.; Cervantes, F.A.; Alvarez, J.M.; Whitworth, J.L. Strain specificity and simultaneous transmission of closely related strains of a potyvirus by Myzus persicae. J. Econ. Entomol. 2012, 105, 783–791. [Google Scholar] [CrossRef]
- Powell, G.; Tosh, C.R.; Hardie, J. Host plant selection by aphids: Behavioral, evolutionary, and applied perspectives. Annu. Rev. Entomol. 2006, 51, 309–330. [Google Scholar] [CrossRef]
- Fereres, A.; Moreno, A. Behavioural aspects influencing plant virus transmission by homopteran insects. Virus Res. 2009, 141, 158–168. [Google Scholar] [CrossRef]
- Stafford, C.A.; Walker, G.P.; Ullman, D.E. Hitching a ride: Vector feeding and virus transmission. Commun. Integr. Biol. 2012, 5, 43–49. [Google Scholar] [CrossRef]
- Halbert, S.E.; Irwin, M.E.; Goodman, R.M. Alate aphid (Homoptera: Aphididae) species and their relative importance as field vectors of soybean mosaic virus. Ann. Appl. Biol. 1981, 97, 1–9. [Google Scholar] [CrossRef]
- Kalleshwaraswamy, C.; Kumar, N.K. Transmission efficiency of papaya ringspot virus by three aphid species. Phytopathology 2008, 98, 541–546. [Google Scholar] [CrossRef]
- Yuan, C.; Ullman, D.E. Comparison of efficiency and propensity as measures of vector importance in zucchini yellow mosaic potyvirus transmission by Aphis gossypii and A. craccivora. Phytopathology 1996, 86, 698–703. [Google Scholar] [CrossRef]
- Perez, P.; Collar, J.; Avilla, C.; Duque, M.; Fereres, A. Estimation of vector propensity of potato virus Y in open-field pepper crops of central Spain. J. Econ. Entomol. 1995, 88, 986–991. [Google Scholar] [CrossRef]
- Nanayakkara, U.; Nie, X.; Giguère, M.; Zhang, J.; Boquel, S.; Pelletier, Y. Aphid feeding behavior in relation to potato virus Y (PVY) acquisition. J. Econ. Entomol. 2012, 105, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Atiri, G.; Enobakhare, D.; Thottappilly, G. The importance of colonizing and non-colonizing aphid vectors in the spread of cowpea aphid-borne mosaic virus in cowpea. Crop Prot. 1986, 5, 406–410. [Google Scholar] [CrossRef]
- Martin, B.; Collar, J.L.; Tjallingii, W.F.; Fereres, A. Intracellular ingestion and salivation by aphids may cause the acquisition and inoculation of non-persistently transmitted plant viruses. J. Gen. Virol. 1997, 78, 2701–2705. [Google Scholar] [CrossRef]
- Will, T.; Vilcinskas, A. The structural sheath protein of aphids is required for phloem feeding. Insect Biochem. Mol. Biol. 2015, 57, 34–40. [Google Scholar] [CrossRef]
- Silva-Sanzana, C.; Celiz-Balboa, J.; Garzo, E.; Marcus, S.E.; Parra-Rojas, J.P.; Rojas, B.; Olmedo, P.; Rubilar, M.A.; Rios, I.; Chorbadjian, R.A. Pectin methylesterases modulate plant homogalacturonan status in defenses against the aphid Myzus persicae. Plant Cell 2019, 31, 1913–1929. [Google Scholar] [CrossRef]
- Restrepo, M.A.; Freed, D.D.; Carrington, J.C. Nuclear transport of plant potyviral proteins. Plant. Cell 1990, 2, 987–998. [Google Scholar] [CrossRef]
- Moreno, A.; Tjallingii, W.F.; Fernandez-Mata, G.; Fereres, A. Differences in the mechanism of inoculation between a semi-persistent and a non-persistent aphid-transmitted plant virus. J. Gen. Virol. 2012, 93, 662–667. [Google Scholar] [CrossRef]
- Powell, G.; Pirone, T.; Hardie, J. Aphid stylet activities during potyvirus acquisition from plants and an in vitro system that correlate with subsequent transmission. Eur. J. Plant. Pathol. 1995, 101, 411–420. [Google Scholar] [CrossRef]
- Powell, G. Intracellular salivation is the aphid activity associated with inoculation of non-persistently transmitted viruses. J. Gen. Virol. 2005, 86, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Garzo, E.; Duque, M.; Fereres, A. Transmission efficiency of different non-persistent viruses infecting melon by four aphid species. Span. J. Agric. Res. 2004, 2, 369–376. [Google Scholar] [CrossRef]
- Nault, L. Arthropod transmission of plant viruses: A new synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. USA 2010, 107, 3600–3605. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Ding, H.; Shiel, P.; Berger, P.H. Volatiles from potato plants infected with potato leafroll virus attract and arrest the virus vector, Myzus persicae (Homoptera: Aphididae). Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Wilfert Eckel, R.V.; Lampert, E. Relative attractiveness of tobacco etch virus-infected and healthy flue-cured tobacco plants to aphids (Homoptera: Aphididae). J. Econ. Entomol. 1996, 89, 1017–1027. [Google Scholar] [CrossRef]
- Fereres, A.; Kampmeier, G.; Irwin, M. Aphid attraction and preference for soybean and pepper plants infected with Potyviridae. Ann. Entomol. Soc. Am. 1999, 92, 542–548. [Google Scholar] [CrossRef]
- Salvaudon, L.; De Moraes, C.M.; Mescher, M.C. Outcomes of co-infection by two potyviruses: Implications for the evolution of manipulative strategies. Proc. R. Soc. Lond. B Biol. Sci. 2013, 280, 20122959. [Google Scholar] [CrossRef]
- Markkula, M.; Laurema, S. Changes in the concentration of free amino acids in plants induced by virus diseases and the reproduction of aphids. Ann. Agric. Fenn. 1964, 3, 265–271. [Google Scholar]
- Castle, S.; Berger, P. Rates of growth and increase of Myzus persicae on virus-infected potatoes according to type of virus-vector relationship. Entomol. Exp. Appl. 1993, 69, 51–60. [Google Scholar] [CrossRef]
- Westwood, J.H.; Lewsey, M.G.; Murphy, A.M.; Tungadi, T.; Bates, A.; Gilligan, C.A.; Carr, J.P. Interference with jasmonic acid-regulated gene expression is a general property of viral suppressors of RNA silencing but only partly explains virus-induced changes in plant–aphid interactions. J. Gen. Virol. 2014, 95, 733. [Google Scholar] [CrossRef] [PubMed]
- Blua, M.J.; Perring, T.M.; Madore, M.A. Plant virus-induced changes in aphid population development and temporal fluctuations in plant nutrients. J. Chem. Ecol. 1994, 20, 691–707. [Google Scholar] [CrossRef] [PubMed]
- Casteel, C.L.; Yang, C.; Nanduri, A.C.; De Jong, H.N.; Whitham, S.A.; Jander, G. The NIa-Pro protein of turnip mosaic virus improves growth and reproduction of the aphid vector, Myzus persicae (green peach aphid). Plant. J. 2014, 77, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Casteel, C.L.; De Alwis, M.; Bak, A.; Dong, H.; Whitham, S.A.; Jander, G. Disruption of ethylene responses by turnip mosaic virus mediates suppression of plant defense against the green peach aphid vector. Plant. Physiol. 2015, 169, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Bak, A.; Patton, M.F.; Perilla-Henao, L.M.; Aegerter, B.J.; Casteel, C.L. Ethylene signaling mediates potyvirus spread by aphid vectors. Oecologia 2019, 190, 139–148. [Google Scholar] [CrossRef]
- Kersch-Becker, M.F.; Thaler, J.S. Virus strains differentially induce plant susceptibility to aphid vectors and chewing herbivores. Oecologia 2014, 174, 883–892. [Google Scholar] [CrossRef]
- Bak, A.; Cheung, A.L.; Yang, C.; Whitham, S.A.; Casteel, C.L. A viral protease relocalizes in the presence of the vector to promote vector performance. Nat. Commun. 2017, 8, 14493. [Google Scholar] [CrossRef]
- Carr, J.P.; Donnelly, R.; Tungadi, T.; Murphy, A.M.; Jiang, S.; Bravo-Cazar, A.; Yoon, J.Y.; Cunniffe, N.J.; Glover, B.J.; Gilligan, C.A. Viral manipulation of plant stress responses and host interactions with insects. Adv. Virus Res. 2018, 102, 177–197. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Bosque-Pérez, N.A.; Davis, T.S. Insect-borne plant pathogens and their vectors: Ecology, evolution, and complex interactions. Ann. Rev. Entomol. 2018, 63, 169–191. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Biochemical and physiological mechanisms underlying effects of cucumber mosaic virus on host-plant traits that mediate transmission by aphid vectors. Plant. Cell Environ. 2014, 37, 1427–1439. [Google Scholar] [CrossRef]
- Carmo-Sousa, M.; Moreno, A.; Garzo, E.; Fereres, A. A non-persistently transmitted-virus induces a pull–push strategy in its aphid vector to optimize transmission and spread. Virus Res. 2014, 186, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, L.; Heathcote, G.; McDermott, N.; Taylor, C. The effect of date of planting and of harvesting potatoes on virus infection and on yield. Ann. Appl. Biol. 1957, 45, 603–622. [Google Scholar] [CrossRef]
- Kennedy, J. Aphid migration and the spread of plant viruses. Nature 1950, 165, 1024–1025. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A. The evolution of aphid life cycles. Annu. Rev. Entomol. 1992, 37, 321–348. [Google Scholar] [CrossRef]
- Pleydell, D.R.; Soubeyrand, S.; Dallot, S.; Labonne, G.; Chadœuf, J.; Jacquot, E.; Thébaud, G. Estimation of the dispersal distances of an aphid-borne virus in a patchy landscape. PLoS Comput. Biol. 2018, 14, e1006085. [Google Scholar] [CrossRef]
- Kimura, K.; Usugi, T.; Hoshi, H.; Kato, A.; Ono, T.; Koyano, S.; Kagiwada, S.; Nishio, T.; Tsuda, S. Surveys of viruliferous alate aphid of plum pox virus in Prunus mume orchards in Japan. Plant. Dis. 2016, 100, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Madden, L.; Raccah, B.; Pirone, T. Modeling plant disease increase as a function of vector numbers: Nonpersistent viruses. Res. Popul. Ecol. 1990, 32, 47–65. [Google Scholar] [CrossRef]
- Madden, L.; Jeger, M.; Van den Bosch, F. A theoretical assessment of the effects of vector-virus transmission mechanism on plant virus disease epidemics. Phytopathology 2000, 90, 576–594. [Google Scholar] [CrossRef]
- Bergstrom, C.T.; McElhany, P.; Real, L.A. Transmission bottlenecks as determinants of virulence in rapidly evolving pathogens. Proc. Natl. Acad. Sci. USA 1999, 96, 5095–5100. [Google Scholar] [CrossRef]
- Pirone, T.P.; Perry, K.L. Aphids: Non-persistent transmission. Adv. Bot. Res. 2002, 36, 1–19. [Google Scholar] [CrossRef]
- Fereres, A. The role of aphid salivation in the transmission of plant viruses. Phytoparasitica 2007, 35, 3–7. [Google Scholar] [CrossRef]
- Pirone, T.P.; Thornbury, D.W. Quantity of virus required for aphid transmission of a potyvirus. Phytopathology 1988, 78, 104–107. [Google Scholar] [CrossRef]
- Moury, B.; Fabre, F.; Senoussi, R. Estimation of the number of virus particles transmitted by an insect vector. Proc. Natl. Acad. Sci. USA 2007, 104, 17891–17896. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Fereres, A.; Cambra, M. Quantitative estimation of plum pox virus targets acquired and transmitted by a single Myzus persicae. Arch. Virol. 2009, 154, 1391–1399. [Google Scholar] [CrossRef]
- Gibson, R.; Rice, A.; Sawicki, R. Effects of the pyrethroid deltamethrin on the acquisition and inoculation of viruses by Myzus persicae. Ann. Appl. Biol. 1982, 100, 49–54. [Google Scholar] [CrossRef]
- Cambra, M.; Vidal, E. Sharka, a vector-borne disease caused by plum pox virus: Vector species, transmission mechanism, epidemiology and mitigation strategies to reduce its natural spread. Acta Hortic. 2017, 1163, 57–68. [Google Scholar] [CrossRef]
- Dupuis, B. The movement of potato virus Y (PVY) in the vascular system of potato plants. Eur. J. Plant. Pathol. 2017, 147, 365–373. [Google Scholar] [CrossRef]
- Boiteau, G.; King, R.; Levesque, D. Lethal and sublethal effects of aldicarh on two potato aphids (Homoptera: Aphididae): Myzus persicae (sulzer) and Macrosiphum euphorbiae (thomas). J. Econ. Entomol. 1985, 78, 41–44. [Google Scholar] [CrossRef]
- Lowery, D.; Boiteau, G. Effects of five insecticides on the probing, walking, and settling behavior of the green peach aphid and the buckthorn aphid (Homoptera: Aphididae) on potato. J. Econ. Entomol. 1988, 81, 208–214. [Google Scholar] [CrossRef]
- Perring, T.M.; Gruenhagen, N.M.; Farrar, C.A. Management of plant viral diseases through chemical control of insect vectors. Annu. Rev. Entomol. 1999, 44, 457–481. [Google Scholar] [CrossRef] [PubMed]
- Boquel, S.; Zhang, J.; Goyer, C.; Giguère, M.A.; Clark, C.; Pelletier, Y. Effect of insecticide-treated potato plants on aphid behavior and potato virus Y acquisition. Pest. Manag. Sci. 2015, 71, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Hooks, C.R.; Fereres, A. Protecting crops from non-persistently aphid-transmitted viruses: A review on the use of barrier plants as a management tool. Virus Res. 2006, 120, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, J.J.; Summers, C.G. Reflective mulches for management of aphids and aphid-borne virus diseases in late-season cantaloupe (Cucumis melo L. var. cantalupensis). Crop Prot. 2002, 21, 891–898. [Google Scholar] [CrossRef]
- Murphy, J.F.; Eubanks, M.D.; Masiri, J. Reflective plastic mulch but not a resistance-inducing treatment reduced watermelon mosaic virus incidence and yield losses in squash. Int. J. Veg. Sci. 2008, 15, 3–12. [Google Scholar] [CrossRef]
- Wróbel, S. Effect of a mineral oil on Myzus persicae capability to spread of PVY and PVM to successive potato plants. J. Plant. Protect. Res. 2007, 47, 383–390. [Google Scholar]
- Boquel, S.; Giguère, M.A.; Clark, C.; Nanayakkara, U.; Zhang, J.; Pelletier, Y. Effect of mineral oil on potato virus Y acquisition by Rhopalosiphum padi. Entomol. Exp. Appl. 2013, 148, 48–55. [Google Scholar] [CrossRef]
- Pinese, B.; Lisle, A.; Ramsey, M.; Halfpapp, K.; DeFaveri, S. Control of aphid-borne papaya ringspot potyvirus in zucchini marrow (Cucurbita pepo) with reflective mulches and mineral oil-insecticide sprays. Int. J. Pest. Manag. 1994, 40, 81–87. [Google Scholar] [CrossRef]
- Boiteau, G.; Singh, M.; Lavoie, J. Crop border and mineral oil sprays used in combination as physical control methods of the aphid-transmitted potato virus Y in potato. Pest. Manag. Sci. 2009, 65, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Boquel, S.; Giguère, M.A.; Pelletier, Y. Effect of mineral oils on host plant selection and probing behavior of Rhopalosiphum padi. Entomol. Exp. Appl. 2016, 160, 241–250. [Google Scholar] [CrossRef]
- Fraser, R. The genetics of resistance to plant viruses. Annu. Rev. Phytopathol. 1990, 28, 179–200. [Google Scholar] [CrossRef]
- Diaz-Pendon, J.A.; Truniger, V.; Nieto, C.; Garcia-Mas, J.; Bendahmane, A.; Aranda, M.A. Advances in understanding recessive resistance to plant viruses. Mol. Plant. Pathol. 2004, 5, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Neriya, Y.; Yamaji, Y.; Namba, S. Recessive resistance to plant viruses: Potential resistance genes beyond translation initiation factors. Front. Microbiol. 2016, 7, 1695. [Google Scholar] [CrossRef] [PubMed]
- Nicaise, V.; German-Retana, S.; Sanjuán, R.; Dubrana, M.P.; Mazier, M.; Maisonneuve, B.; Candresse, T.; Caranta, C.; LeGall, O. The eukaryotic translation initiation factor 4E controls lettuce susceptibility to the potyvirus Lettuce mosaic virus. Plant. Physiol. 2003, 132, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Ruffel, S.; Gallois, J.; Lesage, M.; Caranta, C. The recessive potyvirus resistance gene pot-1 is the tomato orthologue of the pepper pvr2-eIF4E gene. Mol. Genet. Genom. 2005, 274, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Ruffel, S.; Gallois, J.L.; Moury, B.; Robaglia, C.; Palloix, A.; Caranta, C. Simultaneous mutations in translation initiation factors eIF4E and eIF (iso) 4E are required to prevent pepper veinal mottle virus infection of pepper. J. Gen. Virol. 2006, 87, 2089–2098. [Google Scholar] [CrossRef]
- Malnoë, P.; Farinelli, L.; Collet, G.F.; Reust, W. Small-scale field tests with transgenic potato, cv. Bintje, to test resistance to primary and secondary infections with potato virus Y. Plant. Mol. Biol. 1994, 25, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Pehu, T.; Mäki-Valkama, T.; Valkonen, J.; Koivu, K.; Lehto, K.; Pehu, E. Potato plants transformed with a potato virus Y P1 gene sequence are resistant to PVY. Am. Potato J. 1995, 72, 523–532. [Google Scholar] [CrossRef]
- Gargouri-Bouzid, R.; Jaoua, L.; Rouis, S.; Saïdi, M.N.; Bouaziz, D.; Ellouz, R. PVY-resistant transgenic potato plants expressing an anti-NIa protein scFv antibody. Mol. Biotechnol. 2006, 33, 133–140. [Google Scholar] [CrossRef]
- Bouaziz, D.; Ayadi, M.; Bidani, A.; Rouis, S.; Nouri-Ellouz, O.; Jellouli, R.; Drira, N.; Gargouri-Bouzid, R. A stable cytosolic expression of VH antibody fragment directed against PVY NIa protein in transgenic potato plant confers partial protection against the virus. Plant. Sci. 2009, 176, 489–496. [Google Scholar] [CrossRef]
- Farinelli, L.; Malnoë, P.; Collet, G.F. Heterologous encapsidation of potato virus Y strain O (PVY) with the transgenic coat protein of PVY strain N (PVY N) in Solanum tuberosum cv. Bintje. Nat. Biotechnol. 1992, 10, 1020–1025. [Google Scholar] [CrossRef]
- Kaniewski, W.; Lawson, C.; Sammons, B.; Haley, L.; Hart, J.; Delannay, X.; Tumer, N.E. Field resistance of transgenic russeet burbank potato to effects of infection by potato virus X and potato virus Y. Nat. Biotechnol. 1990, 8, 750–754. [Google Scholar] [CrossRef]
- Lawson, C.; Kaniewski, W.; Haley, L.; Rozman, R.; Newell, C.; Sanders, P.; Tumer, N.E. Engineering resistance to mixed virus infection in a commercial potato cultivar: Resistance to potato virus X and potato virus Y in transgenic Russet Burbank. Nat. Biotechnol. 1990, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, D.; Nielsen, S.V.; Albrechtsen, M.; Borkhardt, B. General resistance against potato virus Y introduced into a commercial potato cultivar by genetic transformation with PVY N coat protein gene. Potato Res. 1996, 39, 271–282. [Google Scholar] [CrossRef]
- Smith, G.; Ford, R.; Bryant, J.; Gambley, R.; McGhie, T.; Harding, R.; Dale, J. Expression, purification, and use as an antigen of recombinant sugarcane mosaic virus coat protein. Arch. Virol. 1995, 140, 1817–1831. [Google Scholar] [CrossRef] [PubMed]
- Missiou, A.; Kalantidis, K.; Boutla, A.; Tzortzakaki, S.; Tabler, M.; Tsagris, M. Generation of transgenic potato plants highly resistant to potato virus Y (PVY) through RNA silencing. Mol. Breed. 2004, 14, 185–197. [Google Scholar] [CrossRef]
- Cruz, A.; Aragão, F. RNA i-based enhanced resistance to cowpea severe mosaic virus and cowpea aphid-borne mosaic virus in transgenic cowpea. Plant. Pathol. 2014, 63, 831–837. [Google Scholar] [CrossRef]
- Ling, K.; Namba, S.; Gonsalves, C.; Slightom, J.L.; Gonsalves, D. Protection against detrimental effects of potyvirus infection in transgenic tobacco plants expressing the papaya ringspot virus coat protein gene. Nat. Biotechnol. 1991, 9, 752–758. [Google Scholar] [CrossRef]
- Hamim, I.; Borth, W.B.; Marquez, J.; Green, J.C.; Melzer, M.J.; Hu, J.S. Transgene-mediated resistance to papaya ringspot virus: Challenges and solutions. Phytoparasitica 2018, 46, 1–18. [Google Scholar] [CrossRef]
- Polák, J.; Kundu, J.K.; Krška, B.; Beoni, E.; Komínek, P.; Pívalova, J.; Jarošová, J. Transgenic plum Prunus domestica L., clone C5 (cv. HoneySweet) for protection against sharka disease. J. Integr. Agric. 2017, 16, 516–522. [Google Scholar] [CrossRef]
- Harrison, B.D. Virus variation in relation to resistance-breaking in plants. Euphytica 2002, 124, 181–192. [Google Scholar] [CrossRef]
- Nicaise, V. Crop immunity against viruses: Outcomes and future challenges. Front. Plant. Sci. 2014, 5, 660. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.P.; Lewsey, M.G.; Palukaitis, P. Signaling in induced resistance. Adv. Virus Res. 2010, 76, 57–121. [Google Scholar] [CrossRef]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Montes, C.; Castro, A.; Barba, P.; Rubio, J.; Sánchez, E.; Carvajal, D.; Aguirre, C.; Tapia, E.; Dell, P.; Decroocq, V. Differential RNAi responses of Nicotiana benthamiana individuals transformed with a hairpin-inducing construct during plum pox virus challenge. Virus Genes 2014, 49, 325–338. [Google Scholar] [CrossRef]
- Yang, X.; Niu, L.; Zhang, W.; Yang, J.; Xing, G.; He, H.; Guo, D.; Du, Q.; Qian, X.; Yao, Y. RNAi-mediated SMV P3 cistron silencing confers significantly enhanced resistance to multiple potyvirus strains and isolates in transgenic soybean. Plant Cell Rep. 2018, 37, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Gao, S.; Lin, Q.; Wang, H.; Que, Y.; Xu, L. Transgenic sugarcane resistant to sorghum mosaic virus based on coat protein gene silencing by RNA interference. BioMed Res. Int. 2015, 2015, 861907. [Google Scholar] [CrossRef] [PubMed]
- Worrall, E.A.; Bravo-Cazar, A.; Nilon, A.T.; Fletcher, S.J.; Robinson, K.E.; Carr, J.P.; Mitter, N. Exogenous application of RNAi-inducing double-stranded RNA inhibits aphid-mediated transmission of a plant virus. Front. Plant. Sci. 2019, 10, 265. [Google Scholar] [CrossRef]
- Simón-Mateo, C.; García, J.A. Antiviral strategies in plants based on RNA silencing. Biochim. Biophys. Acta 2011, 1809, 722–731. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.; Kellner, M.J.; Regev, A. RNA targeting with CRISPR–Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Zhan, X.; Zhang, F.; Zhong, Z.; Chen, R.; Wang, Y.; Chang, L.; Bock, R.; Nie, B.; Zhang, J. Generation of virus-resistant potato plants by RNA genome targeting. Plant. Biotechnol. J. 2019, 17, 1814–1822. [Google Scholar] [CrossRef] [PubMed]
- Pyott, D.E.; Sheehan, E.; Molnar, A. Engineering of CRISPR/Cas9-mediated potyvirus resistance in transgene-free Arabidopsis plants. Mol. Plant Pathol. 2016, 17, 1276–1288. [Google Scholar] [CrossRef] [PubMed]
- Bastet, A.; Zafirov, D.; Giovinazzo, N.; Guyon-Debast, A.; Nogué, F.; Robaglia, C.; Gallois, J.L. Mimicking natural polymorphism in eIF 4E by CRISPR-Cas9 base editing is associated with resistance to potyviruses. Plant. Biotechnol. J. 2019, 17, 1736–1750. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J. Simultaneous CRISPR/Cas9-mediated editing of cassava eIF 4E isoforms nCBP-1 and nCBP-2 reduces cassava brown streak disease symptom severity and incidence. Plant. Biotechnol. J. 2019, 17, 421–434. [Google Scholar] [CrossRef]
- Le Trionnaire, G.; Tanguy, S.; Hudaverdian, S.; Gleonnec, F.; Richard, G.; Cayrol, B.; Monsion, B.; Pichon, E.; Deshoux, M.; Webster, C. An integrated protocol for targeted mutagenesis with CRISPR-Cas9 system in the pea aphid. Insect Biochem. Mol. Biol. 2019, 110, 34–44. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus. Viruses 2020, 12, 773. https://doi.org/10.3390/v12070773
Gadhave KR, Gautam S, Rasmussen DA, Srinivasan R. Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus. Viruses. 2020; 12(7):773. https://doi.org/10.3390/v12070773
Chicago/Turabian StyleGadhave, Kiran R., Saurabh Gautam, David A. Rasmussen, and Rajagopalbabu Srinivasan. 2020. "Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus" Viruses 12, no. 7: 773. https://doi.org/10.3390/v12070773
APA StyleGadhave, K. R., Gautam, S., Rasmussen, D. A., & Srinivasan, R. (2020). Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus. Viruses, 12(7), 773. https://doi.org/10.3390/v12070773