Catching Common Cold Virus with a Net: Pyridostatin Forms Filaments in Tris Buffer That Trap Viruses—A Novel Antiviral Strategy?

 , ,
, , 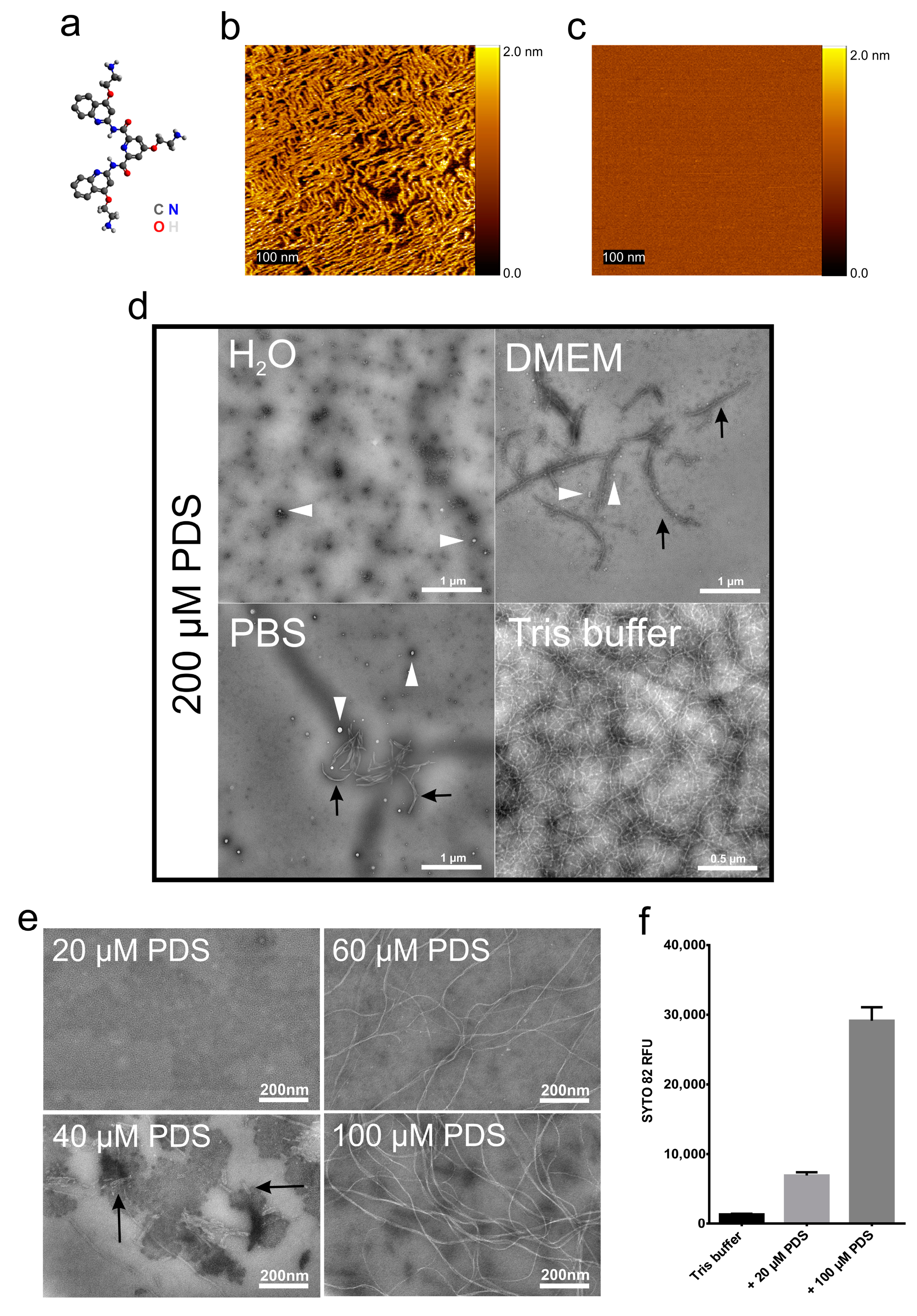{kind=link}
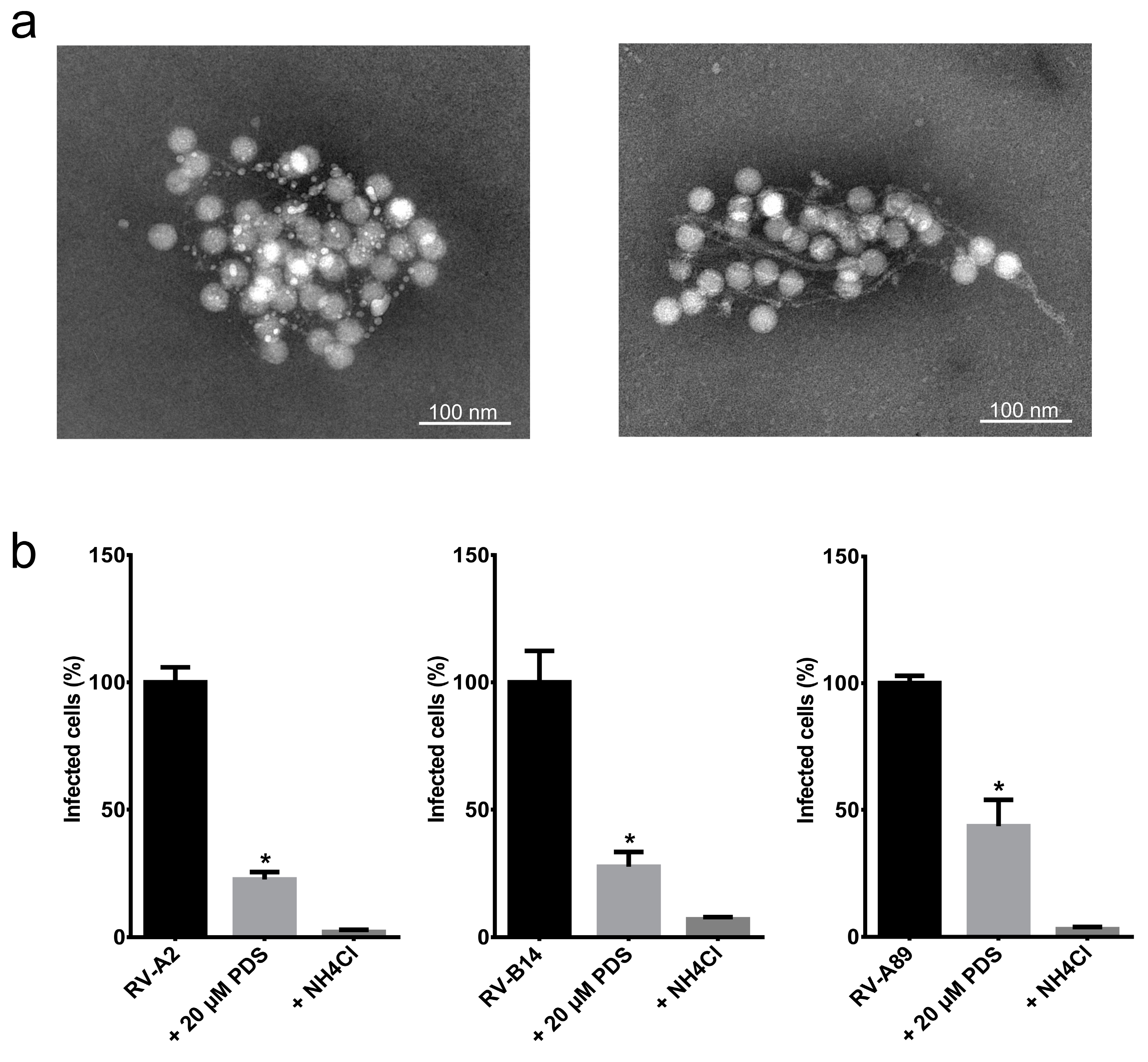{kind=link}
Abstract
Text
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldmann, O.; Medina, E. The expanding world of extracellular traps: Not only neutrophils but much more. Front. Immunol. 2012, 3, 420. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Schonrich, G.; Raftery, M.J. Neutrophil extracellular traps go viral. Front. Immunol. 2016, 7, 366. [Google Scholar] [CrossRef] [PubMed]
- Narayana Moorthy, A.; Narasaraju, T.; Rai, P.; Perumalsamy, R.; Tan, K.B.; Wang, S.; Engelward, B.; Chow, V.T. In vivo and in vitro studies on the roles of neutrophil extracellular traps during secondary pneumococcal pneumonia after primary pulmonary influenza infection. Front. Immunol. 2013, 4, 56. [Google Scholar] [CrossRef] [PubMed]
- Sumby, P.; Barbian, K.D.; Gardner, D.J.; Whitney, A.R.; Welty, D.M.; Long, R.D.; Bailey, J.R.; Parnell, M.J.; Hoe, N.P.; Adams, G.G.; et al. Extracellular deoxyribonuclease made by group a streptococcus assists pathogenesis by enhancing evasion of the innate immune response. Proc. Natl. Acad. Sci. USA 2005, 102, 1679–1684. [Google Scholar] [CrossRef] [PubMed]
- Lavezzo, E.; Berselli, M.; Frasson, I.; Perrone, R.; Palu, G.; Brazzale, A.R.; Richter, S.N.; Toppo, S. G-quadruplex forming sequences in the genome of all known human viruses: A comprehensive guide. PLoS Comput. Biol. 2018, 14, e1006675. [Google Scholar] [CrossRef] [PubMed]
- De Magis, A.; Manzo, S.G.; Russo, M.; Marinello, J.; Morigi, R.; Sordet, O.; Capranico, G. DNA damage and genome instability by g-quadruplex ligands are mediated by r loops in human cancer cells. Proc. Natl. Acad. Sci. USA 2019, 116, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Kremser, L.; Okun, V.M.; Nicodemou, A.; Blaas, D.; Kenndler, E. Binding of fluorescent dye to genomic rna inside intact human rhinovirus after viral capsid penetration investigated by capillary electrophoresis. Anal. Chem. 2004, 76, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Rankl, C.; Kienberger, F.; Wildling, L.; Wruss, J.; Gruber, H.J.; Blaas, D.; Hinterdorfer, P. Multiple receptors involved in human rhinovirus attachment to live cells. Proc. Natl. Acad. Sci. USA 2008, 105, 17778–17783. [Google Scholar] [CrossRef] [PubMed]
- Kozma, E.; Kele, P. Fluorogenic probes for super-resolution microscopy. Org. Biomol. Chem. 2019, 17, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Cattoni, D.I.; Nollmann, M. The fluorescence properties and binding mechanism of sytox green, a bright, low photo-damage DNA intercalating agent. Eur. Biophys. J. 2015, 44, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.A.; Honig, B. A free energy analysis of nucleic acid base stacking in aqueous solution. Biophys. J. 1995, 69, 1528–1535. [Google Scholar] [CrossRef]
- Real-Hohn, A.; Provance, D.W., Jr.; Goncalves, R.B.; Denani, C.B.; de Oliveira, A.C.; Salerno, V.P.; Oliveira Gomes, A.M. Impairing the function of mlck, myosin va or myosin vb disrupts rhinovirus b14 replication. Sci. Rep. 2017, 7, 17153. [Google Scholar] [CrossRef] [PubMed]
- Ganjian, H.; Zietz, C.; Mechtcheriakova, D.; Blaas, D.; Fuchs, R. Icam-1 binding rhinoviruses enter hela cells via multiple pathways and travel to distinct intracellular compartments for uncoating. Viruses 2017, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Abou Alaiwa, M.H.; Launspach, J.L.; Sheets, K.A.; Rivera, J.A.; Gansemer, N.D.; Taft, P.J.; Thorne, P.S.; Welsh, M.J.; Stoltz, D.A.; Zabner, J. Repurposing tromethamine as inhaled therapy to treat cf airway disease. JCI Insight 2016, 1, e87535. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Real-Hohn, A.; Zhu, R.; Ganjian, H.; Ibrahim, N.; Hinterdorfer, P.; Kowalski, H.; Blaas, D. Catching Common Cold Virus with a Net: Pyridostatin Forms Filaments in Tris Buffer That Trap Viruses—A Novel Antiviral Strategy? Viruses 2020, 12, 723. https://doi.org/10.3390/v12070723
Real-Hohn A, Zhu R, Ganjian H, Ibrahim N, Hinterdorfer P, Kowalski H, Blaas D. Catching Common Cold Virus with a Net: Pyridostatin Forms Filaments in Tris Buffer That Trap Viruses—A Novel Antiviral Strategy? Viruses. 2020; 12(7):723. https://doi.org/10.3390/v12070723
Chicago/Turabian StyleReal-Hohn, Antonio, Rong Zhu, Haleh Ganjian, Nahla Ibrahim, Peter Hinterdorfer, Heinrich Kowalski, and Dieter Blaas. 2020. "Catching Common Cold Virus with a Net: Pyridostatin Forms Filaments in Tris Buffer That Trap Viruses—A Novel Antiviral Strategy?" Viruses 12, no. 7: 723. https://doi.org/10.3390/v12070723
APA StyleReal-Hohn, A., Zhu, R., Ganjian, H., Ibrahim, N., Hinterdorfer, P., Kowalski, H., & Blaas, D. (2020). Catching Common Cold Virus with a Net: Pyridostatin Forms Filaments in Tris Buffer That Trap Viruses—A Novel Antiviral Strategy? Viruses, 12(7), 723. https://doi.org/10.3390/v12070723