The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 1: Serology and Virology


Abstract
1. Introduction
- If the porcine foetus becomes infected in utero following intra-nasal exposure of the sow;
- The concentrations of Bungowannah virus RNA in serum, and shed in oropharyngeal secretions and faeces, and whether this is affected by the stage of pregnancy at which the sow is infected;
- If the pig foetus mounts a humoral immune response following in utero infection;
- If persistent infections with Bungowannah virus occur and whether there is a critical stage of gestation at which infection results in this outcome;
- The optimal tissue samples for the detection of Bungowannah virus;
- If PI pigs can readily transmit infection to naïve pigs.
2. Materials and Methods
2.1. Londitudinal Study Design
2.2. Inoculum
2.3. Sample Collection—Pregnant Animals
2.4. Sample Collection—Piglets
2.5. Transmission Study Design
2.6. Sample Collection—Transmission Study
2.7. RNA Extraction and Real-Time Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.8. Serology
2.9. Analysis of Results
3. Results
3.1. Longitudinal Study
3.1.1. Pregnant Pigs
3.1.2. Piglets—qRT-PCR
- Serum: the mean quantity of Bungowannah virus RNA detected in serum at birth exceeded 6.4 log10 copies/mL for all groups of pigs, with the highest levels recorded in those animals where no antibody was detected at birth and where the sow had been infected prior to foetal immunocompetence (D35 (Ab −ve) 7.8 log10 copies/mL; D55 (Ab −ve) 8.4 log10 copies/mL) (Figure 2). For those animals seropositive at birth, the infection gradually cleared over time, although low-level viraemias were still detected at 75 days of age in some animals. At around 28 days of age when maternal antibodies started to wane (Figure 3) the viral load in serum started to rise for the pigs born seronegative in the D35 and D55 groups (Figure 2). By day 120 viral RNA could no longer be detected in the serum of the four remaining D55 (Ab +ve) pigs (data not shown).
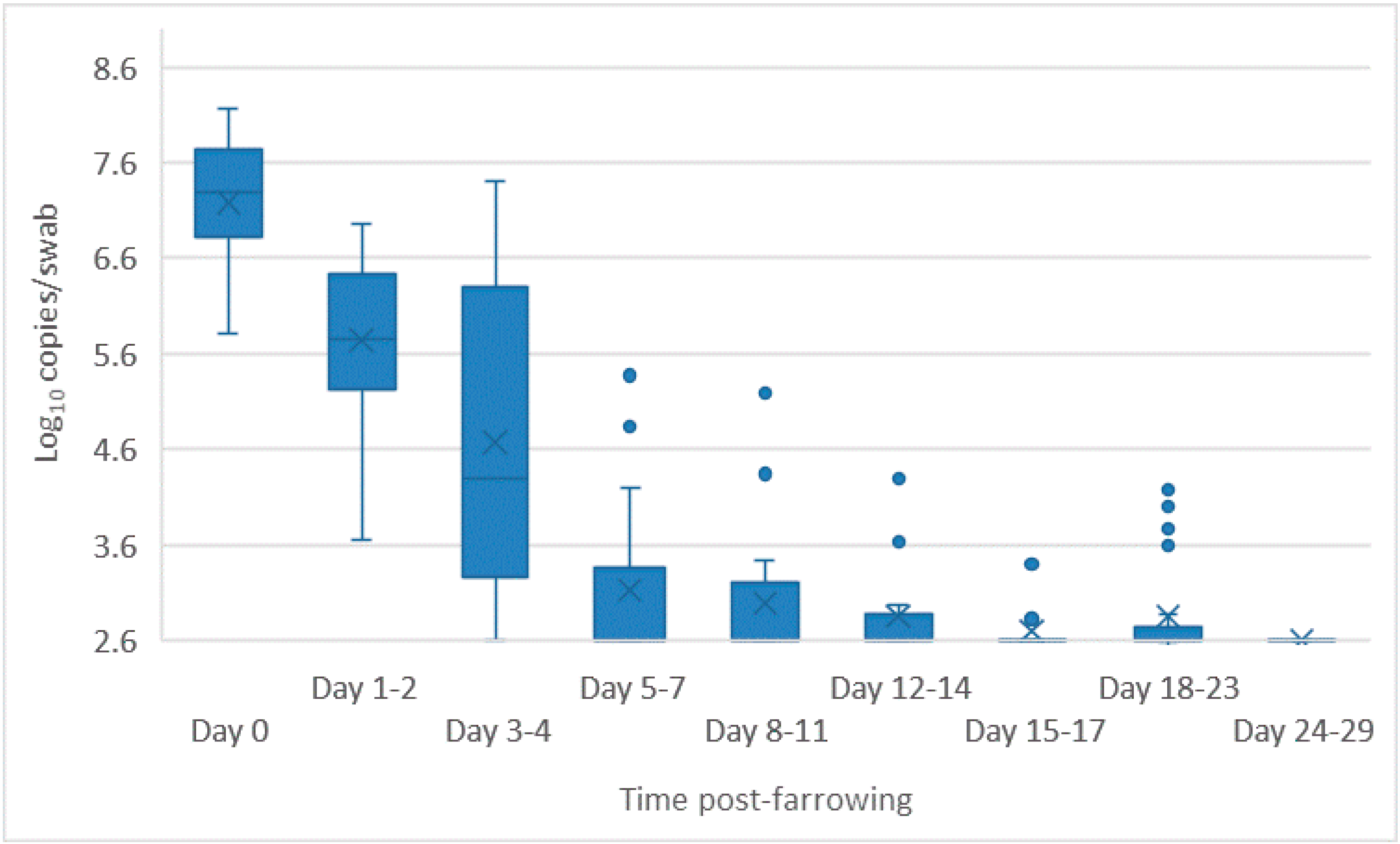
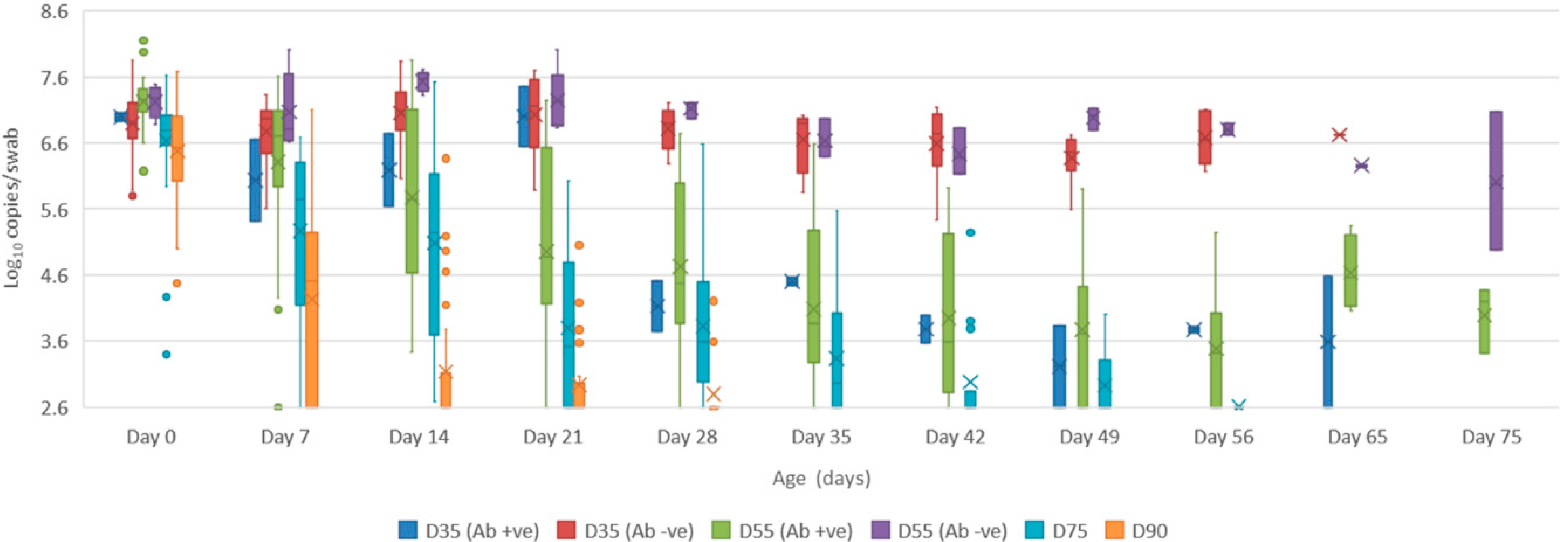
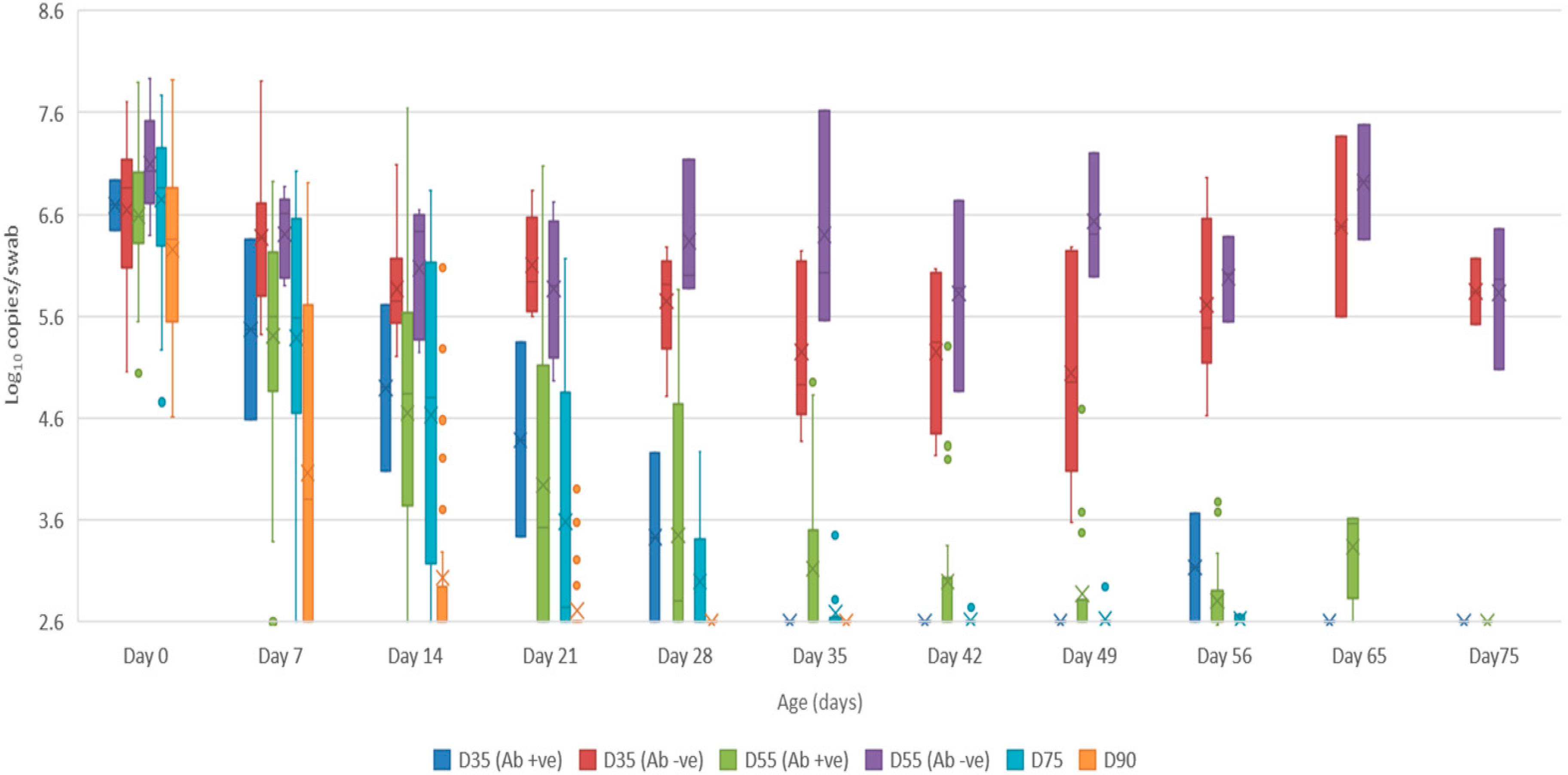
- Oropharyngeal swabs: the mean quantity of viral RNA detected on oropharyngeal swabs at birth ranged from 6.5 (D90) to 7.2 log10 copies/swab (D55 Ab −ve & Ab +ve) (Figure 4). Through to 75 days of age the amount of RNA detected on oropharyngeal swabs remained elevated for the D35 (Ab −ve) and D55 (Ab −ve) groups (Figure 4). In contrast, for all other groups, the amount of RNA decreased over time, reducing most rapidly for the D75 and D90 animals. By day 120 viral RNA could no longer be detected on oropharyngeal swabs of the three D55 (Ab +ve) pigs sampled (data not shown).
- Rectal swabs: the mean level of viral RNA detected on rectal swabs at birth ranged from 6.3 (D90) to 7.1 log10 copies/swab (D55 Ab −ve) (Figure 5). As observed for oropharyngeal swabs, the amount of RNA detected on rectal swabs remained elevated for the D35 (Ab −ve) and D55 (Ab −ve) animals through to day 75. In contrast, for all other groups where the pigs were seropositive at birth, the amount of RNA decreased over time, reducing most rapidly for the D75 and D90 animals. By day 120 viral RNA could not be detected on rectal swabs from the four remaining D55 (Ab +ve) pigs (data not shown). Three animals from the D55 (Ab −ve) group were followed for >6 months (8-01, 8-05 and 10-01). Pig 10-01 appeared to seroconvert between 42 to 49 days old, with a marked reduction in viral load in serum and on rectal swabs from around 84 days of age, and oropharyngeal swabs on day 98 (Figure 6A). Virus was not detected in the serum of 10-01 at time of euthanasia on day 189, but low levels of viral RNA were still detected on oropharyngeal and rectal swabs. Pigs 8-01 and 8-05 seroconverted between 127 and 190 days of age (Figure 6B,C) after which the viraemia was cleared. The amount of virus detected on oropharyngeal and rectal swabs decreased more rapidly after seroconversion for 8-01 compared with 8-05 (Figure 6B,C).
- Mummified foetuses: the mean quantity of viral RNA detected on swabs of the internal organs of mummified foetuses was 5.6 log10 copies/swab for D35 (n = 13; range 4.9–6.9 log10 copies/swab), 5.6 log10 copies/swab for D75 (n = 2; range 5.5–5.7 log10 copies/swab) and 3.0 log10 copies/swab for D90 (n = 1).
- Tissues: at birth and in the first 10 days of life, virus was readily detected in all tissues collected from infected pigs in groups D35, D55 and D75 (Table 1). For group D90, virus was not always detected in tissues sampled at birth but was most likely to be detected in tonsillar swabs and from lymph nodes and heart. For all groups, in the first 10 days of life, the greatest quantity of viral RNA was detected from the tonsils (range of means 5.8 to 7.2 log10 copies/swab) and the brain (range of means 4.8 to 7.0 log10 copies/swab). Subsequently, the amount of Bungowannah viral RNA detected in tissues decreased over time and the proportion of tissues in which viral RNA was detected decreased in those groups where the pigs were seropositive at birth. This decrease occurred most rapidly for the D75 and D90 animals (Table 1). There were only two D35 (Ab +ve) animals identified and by the end of the study (day 75) Bungowannah viral RNA was either no longer detected in tissues or was at low levels, with the highest levels in lymph nodes followed by tonsil (Table 1). In contrast, for D35 (Ab −ve) virus was detected at all time-points for all animals and remained elevated in all tissues with highest quantities detected from tonsil, and in lymph nodes and brain. For the D55 (Ab −ve) group the amount of RNA detected in tissues was generally higher for the first two time-points compared with D55 (Ab +ve) although the number of animals sampled was low. An extensive range of tissues was collected from the presumptively PI pigs (8-01 and 8-05) that were followed until 11 months of age (Table 2). The quantity of Bungowannah virus RNA detected in epididymal semen (9.8 log10 copies/mL) is the highest amount of viral RNA detected in any sample collected throughout the course of the study.
- Urine: urine was collected directly from the bladder of six D35 (Ab −ve) animals at necropsy at between 57–77 days of age in with a mean 6.2 log10 copies/mL (range 5.1–6.9) of viral RNA detected. Virus was also detected in the urine of four D55 (Ab +ve) pigs between 80–90 days of age (mean 4.4 log10 copies/mL) but by day 117 viral RNA was no longer detectable in three. In contrast, Bungowannah virus RNA was only detected in the urine of 2/12 animals from D90 in low quantities when euthanased between 19 and 22 days of age (2.7 log10 copies/mL).
3.1.3. Piglets—Serology
- Animals are detected with antibody prior to suckling (or are presumably in the process of seroconverting – D90). The percentage of seropositive (titres ≥10) pigs for each challenge group seropositive was 4% (D35), 82% (D55), 98% (D75) and 50% (D90). The highest antibody titres were detected in those litters infected earliest in gestation: D35 (mean = 1280; n = 2), D55 (mean = 654; n = 32), D75 (mean = 164; n = 45) and D90 (mean = 27; n = 50). After ingestion of maternal antibodies, the antibody titre rises and stays elevated for the remainder of the study (D35 (Ab +ve); D55 (Ab +ve); D75; D90).
- Animals are seronegative at birth; antibody levels increase with ingestion of maternal antibody and then gradually wane with the animal becoming seronegative again at around 40–60 days of age (D35 (Ab −ve)).
- Animals are seronegative at birth (D55 (Ab −ve); n = 7). These animals have the same serological profile as the D35 group after the ingestion of colostrum. At a variable time after losing maternal antibody these animals seroconvert (10-01 at between 42–49 days of age; 8-01 and 8-05 between 127 and 190 days of age; Figure 6). Note that the seroconversion of 10-01 has resulted in the large box plot for D55 (Ab −ve) from day 42 onwards, as the other two animals in the group (8-01 and 8-05) were seronegative at these time-points.
- Uninfected (seronegative) pigs born to an infected dam; antibody levels increase with ingestion of maternal antibody and then gradually wane with the animal becoming seronegative again at around 60–70 days of age.
3.2. Transmission Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McOrist, S.; Thornton, E.; Peake, A.; Walker, R.; Robson, S.; Finlaison, D.; Kirkland, P.; Reece, R.; Ross, A.; Walker, K.; et al. An infectious myocarditis syndrome affecting late-term and neonatal piglets. Aust. Vet. J. 2004, 82, 509–511. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Frost, M.J.; Finlaison, D.S.; King, K.R.; Ridpath, J.F.; Gu, X. Identification of a novel virus in pigs-Bungowannah virus: A possible new species of pestivirus. Virus Res. 2007, 129, 26–34. [Google Scholar] [CrossRef]
- Hause, B.M.; Collin, E.A.; Peddireddi, L.; Yuan, F.; Chen, Z.; Hesse, R.A.; Gauger, P.C.; Clement, T.; Fang, Y.; Anderson, G. Discovery of a novel putative atypical porcine pestivirus in pigs in the USA. J. Gen. Virol. 2015, 96, 2994–2998. [Google Scholar] [CrossRef]
- Lamp, B.; Schwarz, L.; Hogler, S.; Riedel, C.; Sinn, L.; Rebel-Bauder, B.; Weissenbock, H.; Ladinig, A.; Rumenapf, T. Novel pestivirus species in pigs, Austria, 2015. Emerg. Infect. Dis. 2017, 23, 1176–1179. [Google Scholar] [CrossRef]
- Moennig, V.; Liess, B. Pathogenesis of intrauterine infections with bovine viral diarrhea virus. Vet. Clin. North Am. Food Anim. Pract. 1995, 11, 477–487. [Google Scholar] [CrossRef]
- Roeder, P.L.; Jeffrey, M.; Cranwell, M.P. Pestivirus fetopathogenicity in cattle: Changing sequelae with fetal maturation. Vet. Rec. 1986, 118, 44–48. [Google Scholar] [CrossRef]
- Baker, J.C. The Clinical Manifestations of Bovine Viral Diarrhea Infection. Vet. Clin. N. Am.-Food A. 1995, 11, 425–445. [Google Scholar] [CrossRef]
- McGowan, M.R.; Kirkland, P.D. Early reproductive loss due to bovine pestivirus infection. Br. Vet. J. 1995, 151, 263–270. [Google Scholar] [CrossRef]
- Garcia-Perez, A.L.; Minguijon, E.; Estevez, L.; Barandika, J.F.; Aduriz, G.; Juste, R.A.; Hurtado, A. Clinical and laboratorial findings in pregnant ewes and their progeny infected with Border disease virus (BDV-4 genotype). Res. Vet. Sci. 2009, 86, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.L.; Arruda, P.H.; Magstadt, D.R.; Schwartz, K.J.; Dohlman, T.; Schleining, J.A.; Patterson, A.R.; Visek, C.A.; Victoria, J.G. Identification of a divergent lineage porcine pestivirus in nursing piglets with congenital tremors and reproduction of disease following experimental inoculation. PLoS ONE 2016, 11, e0150104. [Google Scholar] [CrossRef]
- Nettleton, P.F. Border disease. In Infectious Diseases of Livestock, 2nd ed.; Coetzer, J.A.W., Tustin, R.C., Eds.; Oxford University Press Southern Africa: Cape Town, South Africa, 2004; Volume 2, pp. 970–974. [Google Scholar]
- Trautwein, G.; Hewicker, M.; Liess, B.; Orban, S.; Grunert, E. Studies on transplacental transmissibility of a bovine virus diarrhoea (BVD) vaccine virus in cattle III. Occurrence of central nervous system malformations in calves born from vaccinated cows. J. Vet. Med. B. 1986, 33, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Van Oirschot, J.T. Experimental production of congenital persistent swine fever infections: I. Clinical, pathological and virological observations. Vet. Microbiol. 1979, 4, 117–132. [Google Scholar] [CrossRef]
- Meyer, H.; Liess, B.; Frey, H.R.; Hermanns, W.; Trautwein, G. Experimental transplacental transmission of hog cholera virus in pigs. IV. Virological and serological studies in newborn piglets. Zentralbl Veterinarmed B 1981, 28, 659–668. [Google Scholar] [CrossRef]
- Van Oirschot, J.T.; Terpstra, C. A congenital persistent swine fever infection. I. Clinical and virological observations. Vet. Microbiol. 1977, 2, 121–132. [Google Scholar] [CrossRef]
- Van Oirschot, J.T. A congenital persistent swine fever infection. II. Immune response to swine fever virus and unrelated antigens. Vet. Microbiol. 1977, 2, 133–142. [Google Scholar] [CrossRef]
- Nettleton, P.F.; Gilmour, J.S.; Herring, J.A.; Sinclair, J.A. The production and survival of lambs persistently infected with border disease virus. Comp. Immunol. Microbiol. Infect. Dis. 1992, 15, 179–188. [Google Scholar] [CrossRef]
- Barlow, R.M.; Vantsis, J.T.; Gardiner, A.C.; Rennie, J.C.; Herring, J.A.; Scott, F.M.M. Mechanisms of natural transmission of Border disease. J. Comp. Pathol. 1980, 90, 57–65. [Google Scholar] [CrossRef]
- Plateau, E.; Vannier, P.; Tillon, J.P. Atypical hog cholera infection: Viral isolation and clinical study of in utero transmission. Am. J. Vet. Res. 1980, 41, 2012–2015. [Google Scholar]
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical signs and epidemiology of classical swine fever: A review of new knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef]
- Frey, H.R.; Liess, B.; Richter-Reichhelm, H.B.; von Benten, K.; Trautwein, G. Experimental transplacental transmission of hog cholera virus in pigs. I. Virological and serological studies. Zentralbl Veterinarmed B 1980, 27, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Van Oirschot, J.T. Congenital infections with nonarbo togaviruses. Vet. Microbiol. 1983, 8, 321–361. [Google Scholar] [CrossRef]
- Finlaison, D.S.; King, K.R.; Gabor, M.; Kirkland, P.D. An experimental study of Bungowannah virus infection in weaner aged pigs. Vet. Microbiol. 2012, 160, 245–250. [Google Scholar] [CrossRef]
- Finlaison, D.S.; Kirkland, P.D. The outcome of porcine foetal infection with Bungowannah virus is dependent on the stage of gestation at which infection occurs. Part 2: Clinical signs and gross pathology. Viruses 2020. submitted for publication. [Google Scholar]
- Finlaison, D.S.; Cook, R.W.; Srivastava, M.; Frost, M.J.; King, K.R.; Kirkland, P.D. Experimental infections of the porcine foetus with Bungowannah virus, a novel pestivirus. Vet Microbiol 2010, 144, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, C.; Han, D.U.; Chae, C. Simultaneous detection of porcine circovirus type 2 and porcine parvovirus in pigs with PMWS by multiplex PCR. Vet. Rec. 2001, 149, 304–305. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Finlaison, D.S.; King, K.R.; Frost, M.J.; Kirkland, P.D. Field and laboratory evidence that Bungowannah virus, a recently recognised pestivirus, is the causative agent of the porcine myocarditis syndrome (PMC). Vet. Microbiol. 2009, 136, 259–265. [Google Scholar] [CrossRef]
- Cowart, W.O.; Morehouse, L. Effects of attenuated hog cholera virus in pregnant swine at various stages of gestation. J. Am. Vet. Med. Assoc. 1967, 151, 1788–1794. [Google Scholar]
- Paton, D.J.; Done, S.H. Congenital infection of pigs with ruminant-type pestiviruses. J. Comp. Pathol. 1994, 111, 151–163. [Google Scholar] [CrossRef]
- Terpstra, C.; Wensvoort, G. A congenital persistent infection of bovine virus diarrhoea virus in pigs: Clinical, virological and immunological observations. Vet. Q. 1997, 19, 97–101. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Richards, S.G.; Rothwell, J.T.; Stanley, D.F. Replication of bovine viral diarrhoea virus in the bovine reproductive tract and excretion of virus in semen during acute and chronic infections. Vet. Rec. 1991, 128, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, P.D.; McGowan, M.R.; Mackintosh, S.G.; Moyle, A. Insemination of cattle with semen from a bull transiently infected with pestivirus. Vet. Rec. 1997, 140, 124–127. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D35 (Ab +ve) | D35 (Ab −ve) | D55 (Ab +ve) | D55 (Ab −ve) | D75 | D90 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tissue | Age | No. +ve (%) a | Mean b | Range b | No. +ve (%) | Mean | Range | No. +ve (%) | Mean | Range | No. +ve (%) | Mean | Range | No. +ve (%) | Mean | Range | No. +ve (%) | Mean | Range |
| Tonsil | 0–10 | 28 (100%) | 7.1 | 6.2–8.3 | 10 (100%) | 7.2 | 6.3–7.8 | 2 (100%) | 7.2 | 7.0–7.5 | 30 (100%) | 6.7 | 5.0–7.8 | 24 (96%) | 5.8 | ND–7.7 | |||
| 11–30 | 4 (100%) | 6.7 | 5.2–7.6 | 5 (71%) | 4.4 | ND–7.1 | 2 (100%) | 6.5 | 6.4–6.6 | 3 (75%) | 4.0 | ND–6.4 | 1 (5%) | 2.6 | ND–2.8 | ||||
| 31–60 | 5 (100%) | 6.9 | 6.4–7.4 | 8 (44%) | 2.9 | ND–4.3 | 1 (8%) | 2.6 | ND–2.8 | ||||||||||
| >60 | 1 (50%) | 3.3 | ND–4.0 | 3 (100%) | 7.0 | 6.5–7.4 | 10 (48%) | 3.2 | ND–6.8 | 0 (0%) | <2.6 | NA | |||||||
| Lymph | 0–10 | 31 (100%) | 5.3 | 4.4–6.3 | 10 (100%) | 5.4 | 4.5–6.4 | 2 (100%) | 4.5 | 3.9–5.2 | 34 (100%) | 5.3 | 3.9–6.5 | 26 (100%) | 4.7 | 3.2–6.4 | |||
| Node | 11–30 | 4 (100%) | 4.5 | 3.4–5.7 | 7 (100%) | 5.1 | 2.8–6.0 | 2 (100%) | 5.5 | 5.0–6.1 | 4 (100%) | 4.8 | 3.7–5.4 | 12 (55%) | 3.6 | ND–5.6 | |||
| 31–60 | 5 (100%) | 5.3 | 4.5–6.6 | 12 (71%) | 3.9 | ND–6.3 | 10 (77%) | 3.9 | ND–5.4 | ||||||||||
| >60 | 2 (100%) | 4.2 | 4.0–4.5 | 3 (100%) | 6.0 | 5.7–6.2 | 17 (77%) | 4.5 | ND–6.5 | 1 (100%) | 6.5 | NA | |||||||
| Spleen | 0–10 | 30 (100%) | 5.9 | 4.5–7.3 | 10 (100%) | 5.2 | 4.7–5.8 | 2 (100%) | 4.9 | 4.7–5.0 | 34 (100%) | 5.4 | 3.6–6.9 | 18 (69%) | 3.6 | ND–5.2 | |||
| 11–30 | 4 (100%) | 4.5 | 3.4–5.9 | 7 (100%) | 3.8 | 2.9–4.7 | 2 (100%) | 4.3 | 3.8–4.8 | 4 (100%) | 3.5 | 2.8–4.6 | 1 (5%) | 2.6 | ND–2.8 | ||||
| 31–60 | 5 (100%) | 4.6 | 3.8–5.8 | 9 (50%) | 3.1 | ND–4.4 | 0 (0%) | <2.6 | NA | ||||||||||
| >60 | 1 (50%) | 2.8 | ND–3.1 | 3 (100%) | 5.8 | 5.3–6.4 | 14 (67%) | 3.3 | ND–4.9 | 1 (100%) | 3.9 | NA | |||||||
| Thymus | 0–10 | 29 (100%) | 5.6 | 4.1–6.4 | 10 (100%) | 5.2 | 4.3–5.8 | 2 (100%) | 4.9 | 4.8–5.1 | 32 (100%) | 5.4 | 3.3–6.8 | 19 (79%) | 4.1 | ND–5.7 | |||
| 11–30 | 4 (100%) | 4.3 | 3.0–5.1 | 7 (100%) | 3.8 | 2.9–4.9 | 2 (100%) | 4.5 | 4.5–4.6 | 1 (25%) | 2.9 | ND–3.7 | 2 (10%) | 2.6 | ND–3.2 | ||||
| 31–60 | 5 (100%) | 4.6 | 4.0–5.4 | 7 (39%) | 2.8 | ND–3.9 | 1 (8%) | 2.6 | ND–3.0 | ||||||||||
| >60 | 0 (0%) | <2.6 | ND | 3 (100%) | 5.2 | 4.7–5.5 | 9 (41%) | 3.0 | ND–4.5 | 1 (100%) | 3.0 | NA | |||||||
| Lung | 0–10 | 31 (100%) | 6.4 | 5.1–7.6 | 10 (100%) | 6.0 | 5.2–6.8 | 2 (100%) | 6.2 | 6.2–6.2 | 33 (100%) | 6.5 | 4.4–8.2 | 21 (81%) | 4.2 | ND–6.2 | |||
| 11–30 | 4 (100%) | 4.8 | 4.2–5.6 | 7 (100%) | 3.8 | 2.7–5.5 | 2 (100%) | 4.7 | 4.5–4.9 | 4 (100%) | 3.9 | 3.6–4.2 | 1 (5%) | 2.6 | ND–2.9 | ||||
| 31–60 | 5 (100%) | 4.8 | 4.3–5.5 | 11 (61%) | 3.4 | ND–6.8 | 2 (15%) | 2.7 | ND–3.1 | ||||||||||
| >60 | 0 (0%) | <2.6 | ND | 3 (100%) | 5.0 | 4.4–6.1 | 12 (57%) | 3.2 | ND–5.5 | 1 (100%) | 4.1 | NA | |||||||
| Heart | 0–10 | 31 (100%) | 6.1 | 4.3–7.2 | 10 (100%) | 6.0 | 4.9–7.0 | 2 (100%) | 5.6 | 5.5–5.8 | 34 (100%) | 6.0 | 4.2–7.9 | 26 (100%) | 5.4 | 3.9–6.9 | |||
| 11–30 | 4 (100%) | 4.3 | 3.7–5.2 | 7 (100%) | 3.7 | 2.6–4.8 | 2 (100%) | 4.2 | 4.2–4.2 | 4 (100%) | 3.4 | 2.9–4.2 | 8 (36%) | 2.9 | ND–4.1 | ||||
| 31–60 | 5 (100%) | 4.2 | 3.4–5.2 | 4 (22%) | 2.8 | ND–4.3 | 4 (31%) | 2.7 | ND–3.1 | ||||||||||
| >60 | 0 (0%) | <2.6 | ND | 3 (100%) | 4.3 | 3.8–5.1 | 7 (32%) | 2.9 | ND–4.9 | 0 (0%) | <2.6 | NA | |||||||
| Intestine | 0–10 | 31 (100%) | 5.4 | 3.4–7.4 | 10 (100%) | 4.3 | 3.3–5.7 | 2 (100%) | 4.6 | 4.4–4.8 | 34 (100%) | 5.0 | 2.6–6.8 | 17 (68%) | 3.7 | ND–5.5 | |||
| 11–30 | 4 (80%) | 4.4 | ND–5.4 | 3 (43%) | 3.0 | ND–4.5 | 1 (50%) | 3.8 | 2.6–5.1 | 3 (75%) | 3.5 | ND–4.3 | 2 (10%) | 2.7 | ND–3.8 | ||||
| 31–60 | 5 (100%) | 6.2 | 4.2–7.1 | 3 (17%) | 2.7 | ND–3.7 | 4 (31%) | 2.8 | ND–3.3 | ||||||||||
| >60 | 0 (0%) | <2.6 | ND | 3 (100%) | 5.7 | 5.2–6.1 | 10 (45%) | 3.1 | ND–6.2 | 1 (100%) | 4.5 | NA | |||||||
| Brain | 0–10 | 32 (100%) | 7.0 | 6.1–7.8 | 10 (100%) | 6.5 | 5.4–7.7 | 2 (100%) | 6.8 | 6.5–7.2 | 30 (100%) | 6.3 | 5.2–7.4 | 23 (92%) | 4.8 | ND–6.2 | |||
| 11–30 | 4 (100%) | 6.1 | 5.8–6.5 | 7 (100%) | 5.3 | 4.4–6.1 | 2 (100%) | 5.9 | 5.8–6.0 | 4 (100%) | 4.3 | 3.8–5.0 | 4 (18%) | 2.9 | ND–4.8 | ||||
| 31–60 | 5 (100%) | 6.0 | 5.5–6.5 | 14 (78%) | 3.5 | ND–4.5 | 8 (62%) | 3.1 | ND–3.9 | ||||||||||
| >60 | 1 (50%) | 3.1 | ND–3.7 | 3 (100%) | 6.4 | 5.8–7.2 | 17 (77%) | 4.0 | ND–5.9 | 1 (100%) | 3.3 | NA | |||||||
| Animal ID | Sample | Viral Load a | Animal ID | Sample | Viral Load a |
|---|---|---|---|---|---|
| 08-01 | Tonsil | 4.74 | 08-05 | Tonsil | 7.46 |
| Lymph node | 4.82 | Lymph node | 7.31 | ||
| Spleen | 4.36 | Spleen | 4.93 | ||
| Thymus | ND | Thymus | ND | ||
| Brain | ND | Brain | 4.18 | ||
| Heart | 4.25 | Heart | 3.78 | ||
| Lung | ND | Lung | ND | ||
| Intestine | 4.78 | Intestine | 6.34 | ||
| Urine | 6.9 | Urine | 5.8 | ||
| Kidney | ND | Kidney | 3.7 | ||
| Epididymal semen | 9.8 | Ovarian follicular fluid | ND | ||
| Seminal fluid | 3.4 | Cervix | ND | ||
| Bulbourethral gland | 5.8 | Ovary | 2.8 | ||
| Epididymis (head) | 6.2 | Uterus | 4.3 | ||
| Epididymis (tail) | 8.0 | Vagina | 5.2 | ||
| Prostate | 4.1 | ||||
| Seminal vesicle | ND | ||||
| Testis | 6.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finlaison, D.S.; Kirkland, P.D. The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 1: Serology and Virology. Viruses 2020, 12, 691. https://doi.org/10.3390/v12060691
Finlaison DS, Kirkland PD. The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 1: Serology and Virology. Viruses. 2020; 12(6):691. https://doi.org/10.3390/v12060691
Chicago/Turabian StyleFinlaison, Deborah S., and Peter D. Kirkland. 2020. "The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 1: Serology and Virology" Viruses 12, no. 6: 691. https://doi.org/10.3390/v12060691
APA StyleFinlaison, D. S., & Kirkland, P. D. (2020). The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 1: Serology and Virology. Viruses, 12(6), 691. https://doi.org/10.3390/v12060691