Tolerance of Honey Bees to Varroa Mite in the Absence of Deformed Wing Virus

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Processing for Viral Analysis
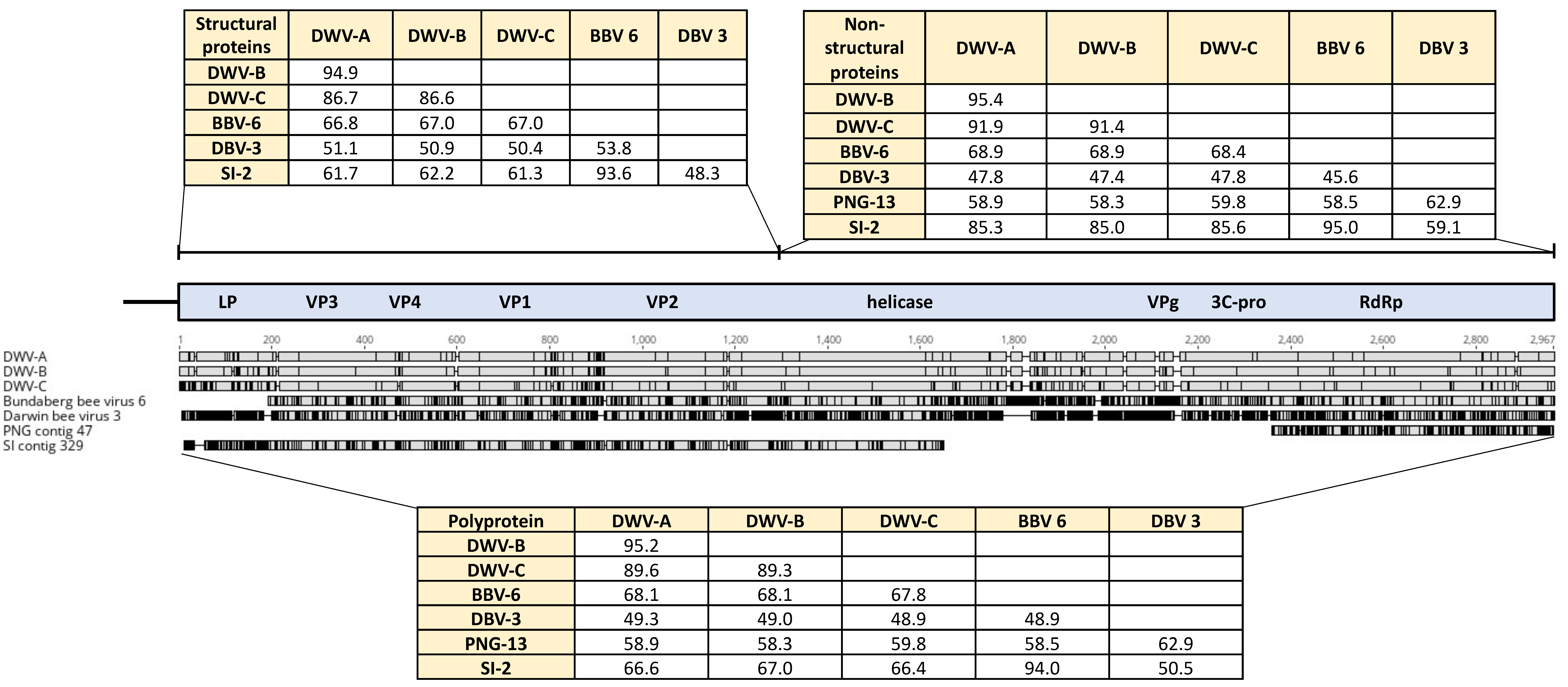
2.3. Viral sequence analysis
3. Results
3.1. HTS Virus Detection in V. jacobsoni Reproducing on A. mellifera
3.2. HTS Virus Detection in A. mellifera in PNG and Solomon Islands 2014
3.3. HTS Virus Detection in A. mellifera and A. cerana in PNG 2018
3.4. Diagnostic RT-PCR for DWV in A. mellifera and A. cerana
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Genersch, E.; Aubert, M. Emerging and re-emerging viruses of the honey bee (Apis mellifera L.). Vet. Res. 2010, 41, 54. [Google Scholar] [CrossRef] [PubMed]
- McMenamin, A.J.; Genersch, E. Honey bee colony losses and associated viruses. Curr. Opin. Insect Sci. 2015, 8, 121–129. [Google Scholar] [CrossRef]
- Grozinger, C.M.; Flenniken, M.L. Bee viruses: Ecology, pathogenicity, and impacts. Ann. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Brettell, L.E. Deformed wing virus in Honeybees and Other Insects. Ann. Rev. Virol. 2019, 6, 49–69. [Google Scholar] [CrossRef]
- Mondet, F.; de Miranda, J.R.; Kretzschmar, A.; Le Conte, Y.; Mercer, A.R. On the Front Line: Quantitative Virus Dynamics in Honeybee (Apis mellifera L.) Colonies along a New Expansion Front of the Parasite Varroa destructor. Plos Pathog. 2014, 10, 1–15. [Google Scholar] [CrossRef]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef]
- Todd, J.H.; De Miranda, J.R.; Ball, B.V. Incidence and molecular characterization of viruses found in dying New Zealand honey bee (Apis mellifera) colonies infested with Varroa destructor. Apidologie 2007, 38, 354–367. [Google Scholar] [CrossRef]
- Roberts, J.M.K.; Anderson, D.L.; Durr, P.A. Metagenomic analysis of Varroa-free Australian honey bees (Apis mellifera) shows a diverse Picornavirales virome. J. Gen. Virol. 2018, 99, 818–826. [Google Scholar] [CrossRef]
- Roberts, J.M.K.; Anderson, D.L.; Durr, P.A. Absence of deformed wing virus and Varroa destructor in Australia provides unique perspectives on honeybee viral landscapes and colony losses. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Beaurepaire, A.; Piot, N.; Doublet, V.; Antunez, K.; Campbell, E.; Chantawannakul, P.; Chejanovsky, N.; Gajda, A.; Heerman, M.; Panziera, D. Diversity and Global Distribution of Viruses of the Western Honey Bee, Apis mellifera. Insects 2020, 11, 239. [Google Scholar] [CrossRef]
- Roberts, J.M.K.; Anderson, D.L.; Tay, W.T. Multiple host shifts by the emerging honeybee parasite, Varroa jacobsoni. Mol. Ecol. 2015, 24, 2379–2391. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.L. Surveillance of parasites and diseases of honeybees in Papua New Guinea and Indonesia. CSIRO Rep. 2008, 1, 1–41. [Google Scholar]
- Mitchener, C. A further note on Apis mellifera in New Guinea. Bee World 1964, 45, 114. [Google Scholar]
- Mitchener, C. The establishment and spread of European honeybees in Australian New Guinea. Bee World 1963, 44, 81. [Google Scholar] [CrossRef]
- Clinch, P.G. Nosema apis and mites in honeybee colonies in Papua New Guinea. J. Apic. Res. 1979, 18, 298–301. [Google Scholar] [CrossRef]
- Anderson, D.L. Mites and pathogens of the European honey bee Apis mellifera and the Eastern hive bee Apis cerana in Papua New Guinea. Nz Mert Rep. 1989, 1, 76. [Google Scholar]
- Anderson, D.L.; Fuchs, S. Two genetically distinct populations of Varroa jacobsoni with contrasting reproductive abilities on Apis mellifera. J. Apic. Res. 1998, 37, 69–78. [Google Scholar] [CrossRef]
- Delfinado-Baker, M.; Aggarwal, K. Infestation of Tropilaelaps clareae and Varroa jacobsoni in Apis mellifera ligustuca colonies in Papua, New Guinea. Am. Bee J. 1987, 127, 443. [Google Scholar]
- Oudemans, A. Note VIII. On a new genus and species of parasitic acari. Notes Leyden Mus. 1904, 24, 216–222. [Google Scholar]
- Anderson, D.L.; Trueman, J.W.H. Varroa jacobsoni (Acari: Varroidae) is more than one species. Exp. Appl. Acarol. 2000, 24, 165–189. [Google Scholar] [CrossRef]
- Anderson, D.; Annand, N.; Lacey, M.; Ete, S. Control of Asian Honeybees in the Solomon Islands; Australian Centre for Internation Agicultural Research: Canberra, Australia, 2012; Volume 1, pp. 1–41. [Google Scholar]
- Forsgren, E.; de Miranda, J.R.; Isaksson, M.; Wei, S.; Fries, I. Deformed wing virus associated with Tropilaelaps mercedesae infesting European honey bees (Apis mellifera). Exp. Appl. Acarol. 2009, 47, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Dainat, B.; Ken, T.; Berthoud, H.; Neumann, P. The ectoparasitic mite Tropilaelaps mercedesae (Acari, Laelapidae) as a vector of honeybee viruses. Insectes Sociaux 2009, 56, 40–43. [Google Scholar] [CrossRef]
- Roberts, J.M.; Schouten, C.N.; Sengere, R.W.; Jave, J.; Lloyd, D. Effectiveness of control strategies for Varroa jacobsoni and Tropilaelaps mercedesae in Papua New Guinea. Exp. Appl. Acarol. 2020, 80, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Kevill, J.; Highfield, A.; Mordecai, G.; Martin, S.; Schroeder, D. ABC assay: Method development and application to quantify the role of three DWV master variants in overwinter colony losses of European honey bees. Viruses 2017, 9, 314. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.S.; Moran, N.A.; Evans, J.D. Early gut colonizers shape parasite susceptibility and microbiota composition in honey bee workers. Proc. Natl. Acad. Sci. USA 2016, 113, 9345–9350. [Google Scholar] [CrossRef]
- Conceição-Neto, N.; Zeller, M.; Lefrère, H.; De Bruyn, P.; Beller, L.; Deboutte, W.; Yinda, C.K.; Lavigne, R.; Maes, P.; Van Ranst, M. Modular approach to customise sample preparation procedures for viral metagenomics: a reproducible protocol for virome analysis. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef]
- Levin, S.; Sela, N.; Erez, T.; Nestel, D.; Pettis, J.; Neumann, P.; Chejanovsky, N. New viruses from the ectoparasite mite Varroa destructor Infesting Apis mellifera and Apis cerana. Viruses 2019, 11, 94. [Google Scholar] [CrossRef]
- Roberts, J.M.K.; Anderson, D.L. A novel strain of sacbrood virus of interest to world apiculture. J. Invertebr. Pathol. 2014, 118, 71–74. [Google Scholar] [CrossRef]
- Galbraith, D.A.; Fuller, Z.L.; Ray, A.M.; Brockmann, A.; Frazier, M.; Gikungu, M.W.; Martinez, J.F.I.; Kapheim, K.M.; Kerby, J.T.; Kocher, S.D. Investigating the viral ecology of global bee communities with high-throughput metagenomics. Sci. Rep. 2018, 8, 8879. [Google Scholar] [CrossRef]
- Wilfert, L.; Long, G.; Leggett, H.C.; Schmid-Hempel, P.; Butlin, R.; Martin, S.J.M.; Boots, M. Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef]
- Norton, A.M.; Remnant, E.J.; Buchmann, G.; Beekman, M. Accumulation and Competition Amongst Deformed Wing Virus Genotypes in Naïve Australian Honeybees Provides Insight Into the Increasing Global Prevalence of Genotype B. Front. Microbiol. 2020, 11, 620. [Google Scholar] [CrossRef] [PubMed]
- Dubois, E.; Dardouri, M.; Schurr, F.; Cougoule, N.; Sircoulomb, F.; Thiery, R. Outcomes of honeybee pupae inoculated with deformed wing virus genotypes A and B. Apidologie 2020, 51, 18–34. [Google Scholar] [CrossRef]
- Yañez, O.; Chávez-Galarza, J.; Tellgren-Roth, C.; Pinto, M.A.; Neumann, P.; de Miranda, J.R. the honeybee (Apis mellifera) developmental state shapes the genetic composition of the deformed wing virus-A quasispecies during serial transmission. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Remnant, E.J.; Mather, N.; Gillard, T.L.; Yagound, B.; Beekman, M. Direct transmission by injection affects competition among RNA viruses in honeybees. Proc. R. Soc. B 2019, 286, 20182452. [Google Scholar] [CrossRef]
- Mondet, F.; Beaurepaire, A.; McAfee, A.; Locke, B.; Alaux, C.; Blanchard, S.; Danka, B.; Le Conte, Y. Honey Bee Survival Mechanisms Against the Parasite Varroa destructor: A Systematic Review of Phenotypic and Genomic Research Efforts. Int. J. Parasitol. 2020. in Press. [Google Scholar]
- Martin, S.J.; Hawkins, G.P.; Brettell, L.E.; Reece, N.; Correia-Oliveira, M.E.; Allsopp, M.H. Varroa destructor reproduction and cell re-capping in mite-resistant Apis mellifera populations. Apidologie 2019, 1–13. [Google Scholar] [CrossRef]
- Thaduri, S.; Stephan, J.G.; de Miranda, J.R.; Locke, B. Disentangling host-parasite-pathogen interactions in a varroa-resistant honeybee population reveals virus tolerance as an independent, naturally adapted survival mechanism. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Locke, B.; Forsgren, E.; de Miranda, J.R. Increased tolerance and resistance to virus infections: A possible factor in the survival of Varroa destructor-resistant honey bees (Apis mellifera). PLoS ONE 2014, 9, e99998. [Google Scholar] [CrossRef]
- Martin, S. A population model for the ectoparasitic mite Varroa jacobsoni in honey bee (Apis mellifera) colonies. Ecol. Model. 1998, 109, 267–281. [Google Scholar] [CrossRef]
- Brettell, L.E.; Martin, S.J. Oldest Varroa tolerant honey bee population provides insight into the origins of the global decline of honey bees. Sci. Rep. 2017, 7, 45953. [Google Scholar] [CrossRef]
- Fung, E.; Hill, K.; Hogendoorn, K.; Glatz, R.V.; Napier, K.R.; Bellgard, M.I.; Barrero, R.A. De novo assembly of honey bee RNA viral genomes by tapping into the innate insect antiviral response pathway. J. Invertebr. Pathol. 2018, 152, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Brettell, L.E.; Riegler, M.; O’Brien, C.; Cook, J.M. Occurrence of honey bee-associated pathogens in Varroa-free pollinator communities. J. Invertebr. Pathol. 2020, 171, 107344. [Google Scholar] [CrossRef] [PubMed]
- Malfroy, S.F.; Roberts, J.M.K.; Perrone, S.; Maynard, G.; Chapman, N. A pest and disease survey of the isolated Norfolk Island honey bee (Apis mellifera) population. J. Apic. Res. 2016, 55, 202–211. [Google Scholar] [CrossRef]
- Roper, T.; Gonzalez, M. Technical Report 49: Disease Survey of Honey Bees in Fiji; Technical Report 49; URS: Adelaide, Australia, 2013. [Google Scholar]
- Taylor, B.; Roper, T. Technical Report 34: Disease Survey of Honey Bees in Vanuatu; URS: Adelaide, Australia, 2013. [Google Scholar]
- Reid, M. Technical Report 35: Disease Survey of Honey Bees in Samoa; URS: Adelaide, Australia, 2012. [Google Scholar]
- Remnant, E.J.; Buchmann, G.; Blacquiere, T.; Holmes, E.C.; Beekman, M.; Ashe, A. A diverse range of novel viruses in geographically distinct honey bee populations. J. Virol. 2017, 91, e00158-17. [Google Scholar] [CrossRef] [PubMed]
- Chapman, N.C.; Sheng, J.; Lim, J.; Malfroy, S.F.; Harpur, B.A.; Zayed, A.; Allsopp, M.H.; Rinderer, T.E.; Roberts, J.M.; Remnant, E.J. Genetic origins of honey bees (Apis mellifera) on Kangaroo Island and Norfolk Island (Australia) and the Kingdom of Tonga. Apidologie 2019, 50, 28–39. [Google Scholar] [CrossRef]
- Martin, S.J. The role of Varroa and viral pathogens in the collapse of honeybee colonies: a modelling approach. J. Appl. Ecol. 2001, 38, 1082–1093. [Google Scholar] [CrossRef]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef]
- Wang, S.; Chen, G.; Lin, Z.; Wu, Y.; Hu, F.; Zheng, H. Occurrence of multiple honeybee viruses in the ectoparasitic mites Varroa spp. in Apis cerana colonies. J. Invertebr. Pathol. 2019, 166, 107225. [Google Scholar] [CrossRef]
- Levin, S.; Galbraith, D.; Sela, N.; Erez, T.; Grozinger, C.; Chejanovsky, N. Presence of Apis rhabdovirus-1 in populations of pollinators and their parasites from two continents. Front. Microbiol. 2017, 8, 2482. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Harvey, E.; Rose, K.; Eden, J.-S.; Lo, N.; Abeyasuriya, T.; Shi, M.; Doggett, S.L.; Holmes, E.C. Extensive diversity of RNA viruses in Australian ticks. J. Virol. 2019, 93, e01358-18. [Google Scholar] [CrossRef] [PubMed]
- Herrero, S.; Millán-Leiva, A.; Coll, S.; González-Martínez, R.M.; Parenti, S.; Gonzalez-Cabrera, J. Identification of new viral variants specific to the honey bee mite Varroa destructor. Exp. Appl. Acarol. 2019, 79, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.; Sela, N.; Chejanovsky, N. Two novel viruses associated with the Apis mellifera pathogenic mite Varroa destructor. Sci. Rep. 2016, 6, 37710. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Viruses Detected | Vd Homolog | Vj-B1 2008 | Vj-C1 2008 | PNG Am 2014 | Sol. Is. Am 2014 | PNG Am 2018 | PNG Ac 2018 |
|---|---|---|---|---|---|---|---|
| VJRV-1 (MT482464) | ARV-1 | 24,476 | 29,575 | 0 | 0 | 31 | 0 |
| VJRV-2 (MT482465) | ARV-2 | 34,735 | 77,497 | 0 | 0 | 0 | 0 |
| VJV-2 PNG1 (MT482466) | VDV-2 | 199,968 | 130,682 | 0 | 0 | 0 | 0 |
| VJV-2 PNG2 (MT482467) | VDV-2 | 511,272 | 407,942 | 0 | 0 | 0 | 0 |
| VJV-4 (MT482468) | VDV-4 | 229,548 | 354,301 | 0 | 0 | 0 | 0 |
| SBV | na | 0 | 0 | 705,914 | 886,074 | 36,903 | 379,433 |
| BQCV | na | 0 | 0 | 32,943 | 8680 | 7028 | 55 |
| IAPV | na | 0 | 0 | 15 | 21 | 5 | 0 |
| LSV-1 | na | 0 | 0 | 2620 | 0 | 66,617 | 0 |
| LSV-2 | na | 0 | 0 | 8628 | 43,407 | 45,055 | 0 |
| LSV-3 | na | 0 | 0 | 7473 | 0 | 91,150 | 82 |
| LSV-8 | na | 0 | 0 | 176,038 | 0 | 272,652 | 215 |
| Novel Virus | Length | Taxonomy | Closest Relative | % Amino Acid Identity | GenBank Accession |
|---|---|---|---|---|---|
| PNG 2014 | |||||
| PNG bee virus 1 | 10,129 | Caliciviridae | Hubei picorna-like virus 68 | 29 | MT482483 |
| PNG bee virus 2 | 7385 | Dicistroviridae | Melipona quadrifasciata virus 1a | 38 | MT482484 |
| PNG bee virus 3 | 4258 | Dicistroviridae | Plautia stali intestine virus | 61 | MT482485 |
| PNG bee virus 4 | 4236 | unclassified | Hubei picorna-like virus 51 | 43 | MT482486 |
| PNG bee virus 5 | 4084 | unclassified | Bundaberg bee virus 8 | 28 | MT482487 |
| PNG bee virus 6 | 4077 | unclassified | Hubei arthropod virus 1 | 37 | MT482488 |
| PNG bee virus 7 | 4017 | unclassified | Darwin bee virus 6 | 44 | MT482489 |
| PNG bee virus 8 | 3854 | Dicistroviridae | Solenopsis invicta virus 13 | 51 | MT482490 |
| PNG bee virus 9 | 3738 | Caliciviridae | Thika virus | 45 | MT482491 |
| PNG bee virus 10 | 3483 | Iflaviridae | La Jolla virus | 45 | MT482492 |
| PNG bee virus 11 | 3082 | Dicistroviridae | Darwin bee virus 6 | 70 | MT482493 |
| PNG bee virus 12 | 3065 | Caliciviridae | Hubei picorna-like virus 67 | 24 | MT482494 |
| PNG bee virus 13 * | 2063 | Iflaviridae | Darwin bee virus 3 | 62 | MT482495 |
| Solomon Islands 2014 | |||||
| SI bee virus 1 | 5882 | Iflaviridae | Bradson virus | 55 | MT482497 |
| SI bee virus 2 * | 4799 | Iflaviridae | Bundaberg bee virus 6 | 94 | MT482498 |
| SI bee virus 3 | 4280 | unclassified | Riptortus pedestris virus-1 | 74 | MT482499 |
| PNG 2018 samples | |||||
| PNG bee virus 9 | 4567 | unclassified | Thika virus | 45 | MT482491 |
| PNG bee virus 14 (A. mellifera) | 3137 | Nodaviridae | Mosinovirus–RNA1 | 42 | MT482496 |
| PNG bee virus 14 (A. cerana) | 2115 | Nodaviridae | Mosinovirus–RNA1 | 42 | MT482496 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts, J.M.K.; Simbiken, N.; Dale, C.; Armstrong, J.; Anderson, D.L. Tolerance of Honey Bees to Varroa Mite in the Absence of Deformed Wing Virus. Viruses 2020, 12, 575. https://doi.org/10.3390/v12050575
Roberts JMK, Simbiken N, Dale C, Armstrong J, Anderson DL. Tolerance of Honey Bees to Varroa Mite in the Absence of Deformed Wing Virus. Viruses. 2020; 12(5):575. https://doi.org/10.3390/v12050575
Chicago/Turabian StyleRoberts, John M. K., Nelson Simbiken, Chris Dale, Joel Armstrong, and Denis L. Anderson. 2020. "Tolerance of Honey Bees to Varroa Mite in the Absence of Deformed Wing Virus" Viruses 12, no. 5: 575. https://doi.org/10.3390/v12050575
APA StyleRoberts, J. M. K., Simbiken, N., Dale, C., Armstrong, J., & Anderson, D. L. (2020). Tolerance of Honey Bees to Varroa Mite in the Absence of Deformed Wing Virus. Viruses, 12(5), 575. https://doi.org/10.3390/v12050575