Genome Analysis and Replication Studies of the African Green Monkey Simian Foamy Virus Serotype 3 Strain FV2014

Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Preparation
2.2. PCR Assays and Copy Number Standard
2.3. Infectivity Analysis
2.4. Preparation of Viral Nucleic Acid, Sequencing, and Bioinformatics Analysis
2.4.1. High-Throughput Sequencing
2.4.2. Sanger Sequencing
2.5. Sequence Comparison and Phylogenetic Analysis
3. Results
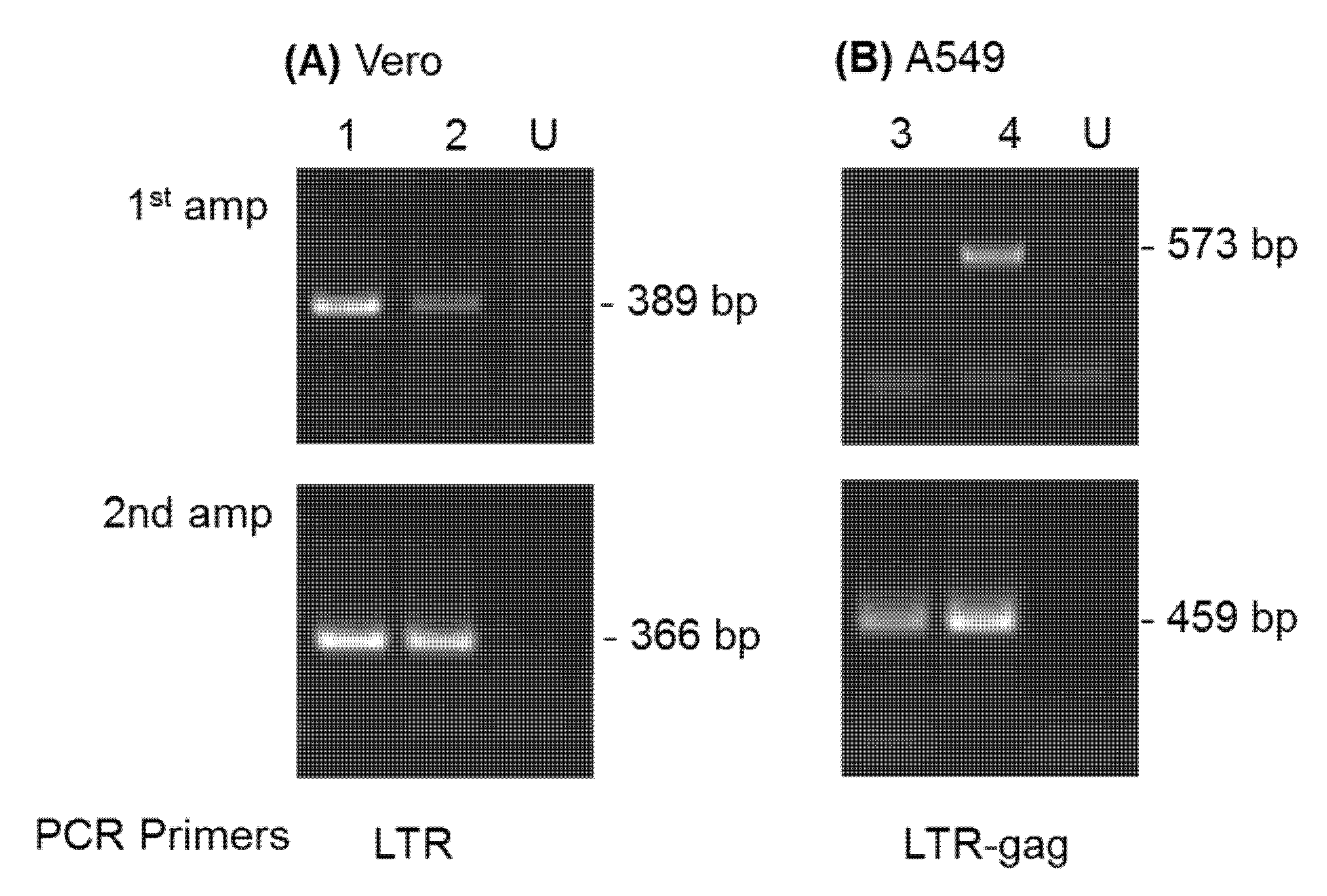
3.1. Characterization of SFVcae_FV2014 Virus Stock
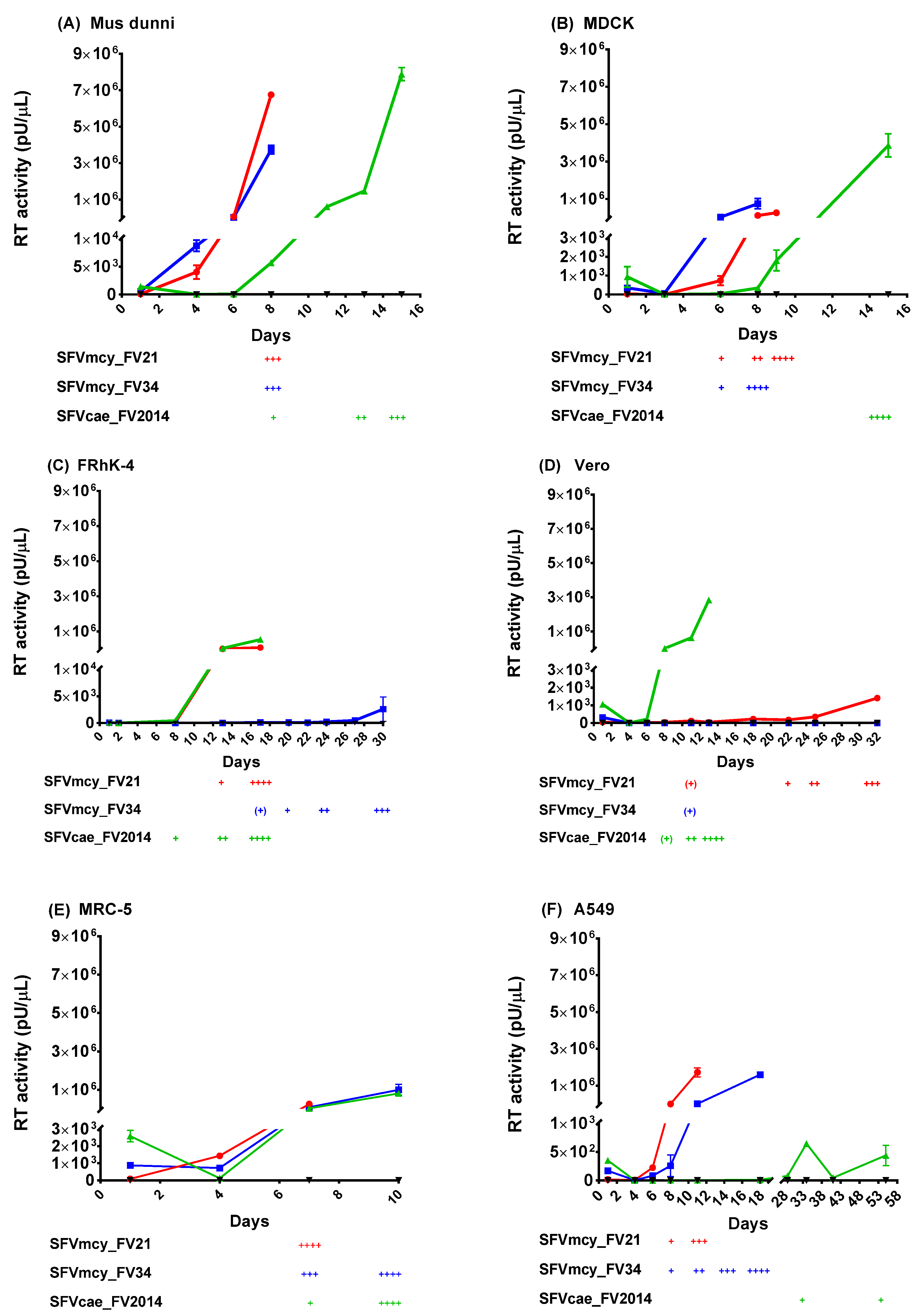
3.2. Studies of SFVcae_FV2014 Replication
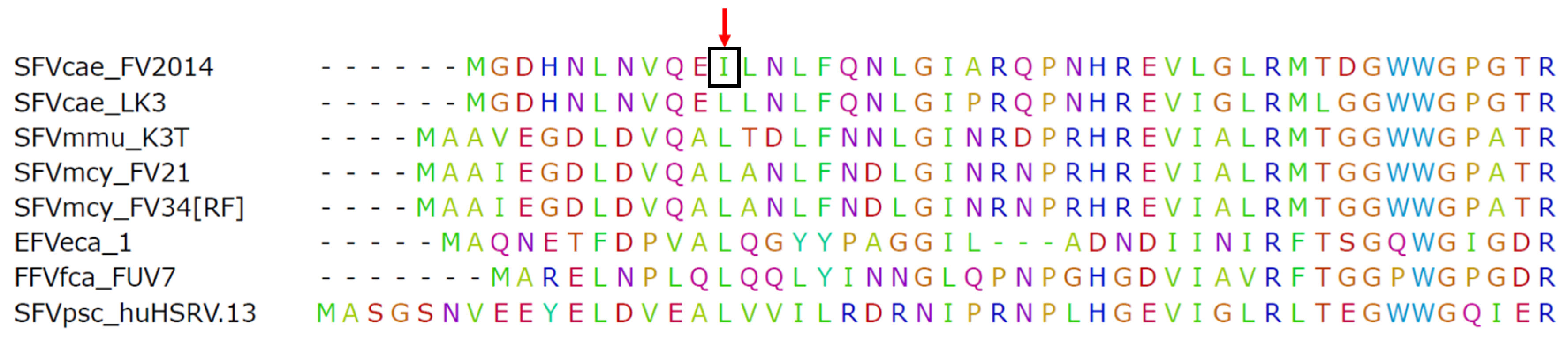
3.3. SFVcae_FV2014 Genome Analysis
3.3.1. Structure and Sequence Comparison
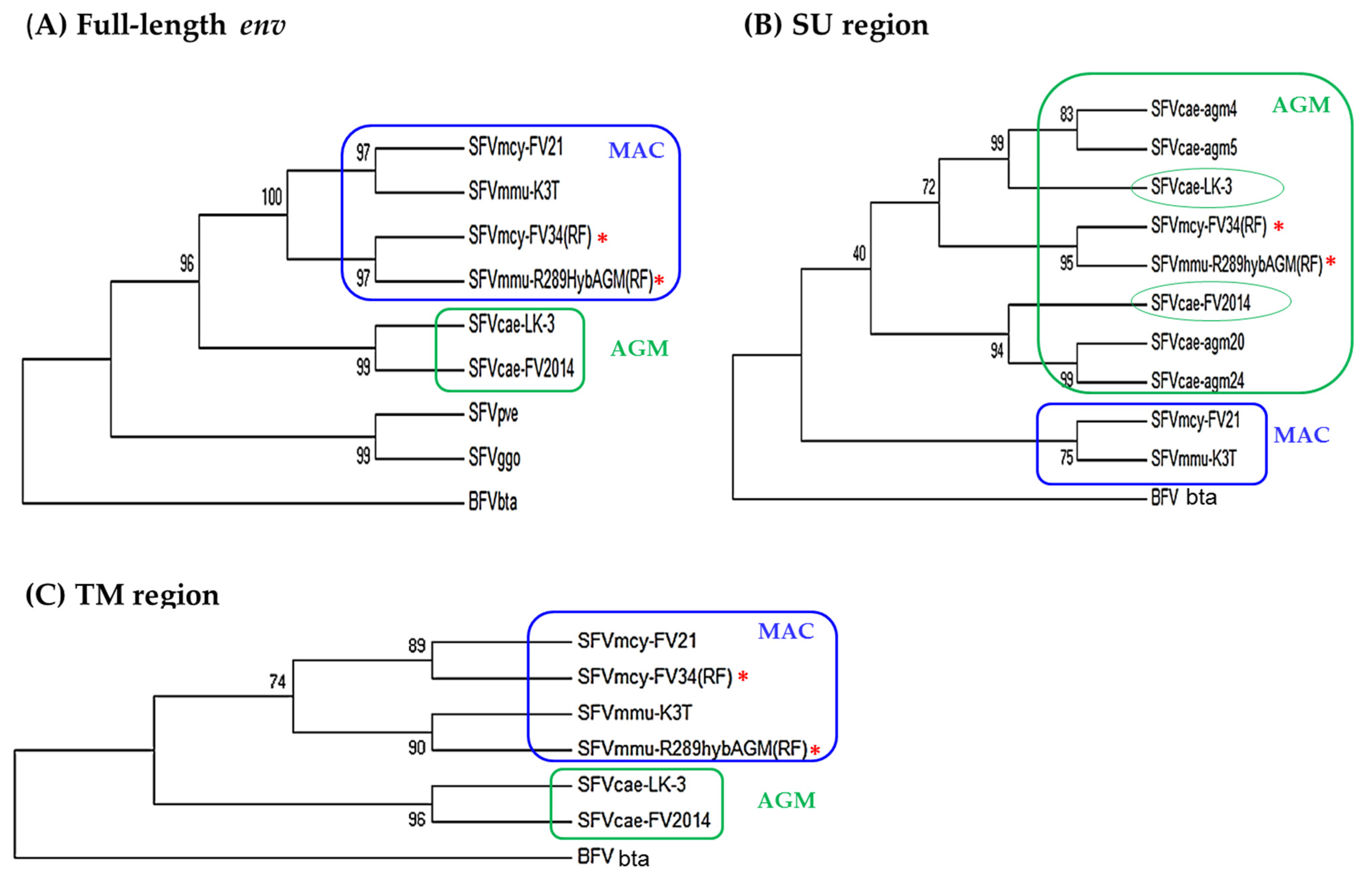
3.3.2. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Khan, A.S.; Bodem, J.; Buseyne, F.; Gessain, A.; Johnson, W.; Kuhn, J.H.; Kuzmak, J.; Lindemann, D.; Linial, M.L.; Lochelt, M.; et al. Spumaretroviruses: Updated taxonomy and nomenclature. Virology 2018, 516, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Stiles, G.E.; Bittle, J.L.; Cabasso, V.J. Comparison of simian foamy virus strains including a new serological type. Nature 1964, 201, 1350–1351. [Google Scholar] [CrossRef] [PubMed]
- Johnston, P.B. A second immunologic type of simian foamy virus: Monkey throat infections and unmasking by both types. J. Infect. Dis. 1961, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Johnston, P.B. Strain fv-21 of simian foamy virus type 1 was cloned and sequenced after isolation from the taiwan monkey macaca cyclopsis. J. Microbiol. Immunol. Infect. 2000, 33, 60–61. [Google Scholar]
- Neumann-Haefelin, D.; Rethwilm, A.; Bauer, G.; Gudat, F.; zur Hausen, H. Characterization of a foamy virus isolated from cercopithecus aethiops lymphoblastoid cells. Med. Microbiol. Immunol. 1983, 172, 75–86. [Google Scholar] [CrossRef]
- Schweizer, M.; Corsten, B.; Neumann-Haefelin, D. Heterogeneity of primate foamy virus genomes. Arch. Virol. 1988, 99, 125–134. [Google Scholar] [CrossRef]
- Renne, R.; Friedl, E.; Schweizer, M.; Fleps, U.; Turek, R.; Neumann-Haefelin, D. Genomic organization and expression of simian foamy virus type 3 (sfv-3). Virology 1992, 186, 597–608. [Google Scholar] [CrossRef]
- Pinto-Santini, D.M.; Stenbak, C.R.; Linial, M.L. Foamy virus zoonotic infections. Retrovirology 2017, 14, 55. [Google Scholar] [CrossRef]
- Schweizer, M.; Schleer, H.; Pietrek, M.; Liegibel, J.; Falcone, V.; Neumann-Haefelin, D. Genetic stability of foamy viruses: Long-term study in an african green monkey population. J. Virol. 1999, 73, 9256–9265. [Google Scholar] [CrossRef]
- Liu, W.; Worobey, M.; Li, Y.; Keele, B.F.; Bibollet-Ruche, F.; Guo, Y.; Goepfert, P.A.; Santiago, M.L.; Ndjango, J.-B.; Neel, C.; et al. Molecular ecology and natural history of simian foamy virus infection in wild-living chimpanzees. PLoS Pathog. 2008, 4, e1000097. [Google Scholar] [CrossRef]
- Galvin, T.A.; Ahmed, I.A.; Shahabuddin, M.; Bryan, T.; Khan, A.S. Identification of recombination in the envelope gene of simian foamy virus serotype 2 isolated from macaca cyclopis. J. Virol. 2013, 87, 8792–8797. [Google Scholar] [CrossRef] [PubMed]
- Blochmann, R.; Curths, C.; Coulibaly, C.; Cichutek, K.; Kurth, R.; Norley, S.; Bannert, N.; Fiebig, U. A novel small animal model to study the replication of simian foamy virus In Vivo. Virology 2014, 448, 65–73. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nandakumar, S.; Bae, E.H.; Khan, A.S. Complete genome sequence of a naturally occurring simian foamy virus isolate from rhesus macaque (sfvmmu_k3t). Genome Announc. 2017, 5, e00827-17. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.S.; Sears, J.F.; Muller, J.; Galvin, T.A.; Shahabuddin, M. Sensitive assays for isolation and detection of simian foamy retroviruses. J. Clin. Microbiol. 1999, 37, 2678–2686. [Google Scholar] [CrossRef]
- Karber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn Schmiedebergs Arch. Exp. Pathol. Pharmakol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Sears, J.F.; Khan, A.S. Single-tube fluorescent product-enhanced reverse transcriptase assay with ampliwax (stf-pert) for retrovirus quantitation. J. Virol. Methods 2003, 108, 139–142. [Google Scholar] [CrossRef]
- Nandakumar, S.; Bae, E.H.; Khan, A.S. Complete genome sequence of the african green monkey simian foamy virus serotype 3 strain fv2014. Genome Announc. 2017. [Google Scholar] [CrossRef]
- Quail, M.A.; Smith, M.; Coupland, P.; Otto, T.D.; Harris, S.R.; Connor, T.R.; Bertoni, A.; Swerdlow, H.P.; Gu, Y. A tale of three next generation sequencing platforms: Comparison of ion torrent, pacific biosciences and illumina miseq sequencers. BMC Genom. 2012, 13, 341. [Google Scholar] [CrossRef]
- Saib, A.; Koken, M.H.; van der Spek, P.; Peries, J.; de The, H. Involvement of a spliced and defective human foamy virus in the establishment of chronic infection. J. Virol. 1995, 69, 5261–5268. [Google Scholar] [CrossRef]
- Giron, M.L.; de The, H.; Saib, A. An evolutionarily conserved splice generates a secreted env-bet fusion protein during human foamy virus infection. J. Virol. 1998, 72, 4906–4910. [Google Scholar] [CrossRef]
- Khan, A.S.; Kumar, D. Simian foamy virus infection by whole-blood transfer in rhesus macaques: Potential for transfusion transmission in humans. Transfusion 2006, 46, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Maurer, B.; Bannert, H.; Darai, G.; Flugel, R.M. Analysis of the primary structure of the long terminal repeat and the gag and pol genes of the human spumaretrovirus. J. Virol. 1988, 62, 1590–1597. [Google Scholar] [CrossRef] [PubMed]
- Aiewsakun, P.; Richard, L.; Gessain, A.; Mouinga-Ondeme, A.; Vicente Afonso, P.; Katzourakis, A. Modular nature of simian foamy virus genomes and their evolutionary history. Virus Evol. 2019, 5, vez032. [Google Scholar] [CrossRef] [PubMed]
- Switzer, W.M.; Salemi, M.; Shanmugam, V.; Gao, F.; Cong, M.E.; Kuiken, C.; Bhullar, V.; Beer, B.E.; Vallet, D.; Gautier-Hion, A.; et al. Ancient co-speciation of simian foamy viruses and primates. Nature 2005, 434, 376–380. [Google Scholar] [CrossRef]
- Feeroz, M.M.; Soliven, K.; Small, C.T.; Engel, G.A.; Andreina Pacheco, M.; Yee, J.L.; Wang, X.; Kamrul Hasan, M.; Oh, G.; Levine, K.L.; et al. Population dynamics of rhesus macaques and associated foamy virus in bangladesh. Emerg. Microbes Infect. 2013, 2, e29. [Google Scholar] [CrossRef]
- Calattini, S.; Wanert, F.; Thierry, B.; Schmitt, C.; Bassot, S.; Saib, A.; Herrenschmidt, N.; Gessain, A. Modes of transmission and genetic diversity of foamy viruses in a macaca tonkeana colony. Retrovirology 2006, 3, 23. [Google Scholar] [CrossRef]
- Ghersi, B.M.; Jia, H.; Aiewsakun, P.; Katzourakis, A.; Mendoza, P.; Bausch, D.G.; Kasper, M.R.; Montgomery, J.M.; Switzer, W.M. Wide distribution and ancient evolutionary history of simian foamy viruses in new world primates. Retrovirology 2015, 12, 89. [Google Scholar] [CrossRef]
- Katzourakis, A.; Aiewsakun, P.; Jia, H.; Wolfe, N.D.; LeBreton, M.; Yoder, A.D.; Switzer, W.M. Discovery of prosimian and afrotherian foamy viruses and potential cross species transmissions amidst stable and ancient mammalian co-evolution. Retrovirology 2014, 11, 61. [Google Scholar] [CrossRef]
- Betsem, E.; Rua, R.; Tortevoye, P.; Froment, A.; Gessain, A. Frequent and recent human acquisition of simian foamy viruses through apes’ bites in central africa. PLoS Pathog. 2011, 7, e1002306. [Google Scholar] [CrossRef]
- Calattini, S.; Nerrienet, E.; Mauclere, P.; Georges-Courbot, M.C.; Saib, A.; Gessain, A. Natural simian foamy virus infection in wild-caught gorillas, mandrills and drills from cameroon and gabon. J. Gen. Virol. 2004, 85, 3313–3317. [Google Scholar] [CrossRef]
- Calattini, S.; Nerrienet, E.; Mauclere, P.; Georges-Courbot, M.C.; Saib, A.; Gessain, A. Detection and molecular characterization of foamy viruses in Central African chimpanzees of the Pan troglodytes troglodytes and Pan troglodytes vellerosus subspecies. J. Med. Primatol. 2006, 35, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Mouinga-Ondémé, A.; Betsem, E.; Caron, M.; Makuwa, M.; Sallé, B.; Renault, N.; Saïb, A.; Telfer, P.; Marx, P.; Gessain, A.; et al. Two distinct variants of simian foamy virus in naturally infected mandrills (Mandrillus sphinx) and cross-species transmission to humans. Retrovirology 2010, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Rethwilm, A.; Bodem, J. Evolution of foamy viruses: The most ancient of all retroviruses. Viruses 2013, 5, 2349–2374. [Google Scholar] [CrossRef] [PubMed]
- Leendertz, S.A.; Junglen, S.; Hedemann, C.; Goffe, A.; Calvignac, S.; Boesch, C.; Leendertz, F.H. High prevalence, coinfection rate, and genetic diversity of retroviruses in wild red colobus monkeys (Piliocolobus badius badius) in tai national park, cote d’ivoire. J. Virol. 2010, 84, 7427–7436. [Google Scholar] [CrossRef]
- Richard, L.; Rua, R.; Betsem, E.; Mouinga-Ondémé, A.; Kazanji, M.; Leroy, E.; Njouom, R.; Buseyne, F.; Afonso, P.V.; Gessain, A. Cocirculation of two env molecular variants, of possible recombinant origin, in gorilla and chimpanzee simian foamy virus strains from central africa. J. Virol. 2015, 89, 12480–12491. [Google Scholar] [CrossRef]
- Winkler, I.G.; Flugel, R.M.; Lochelt, M.; Flower, R.L. Detection and molecular characterisation of feline foamy virus serotypes in naturally infected cats. Virology 1998, 247, 144–151. [Google Scholar] [CrossRef]
- Yu, S.F.; Stone, J.; Linial, M.L. Productive persistent infection of hematopoietic cells by human foamy virus. J. Virol. 1996, 70, 1250–1254. [Google Scholar] [CrossRef]
- Mergia, A.; Leung, N.J.; Blackwell, J. Cell tropism of the simian foamy virus type 1 (sfv-1). J. Med. Primatol. 1996, 25, 2–7. [Google Scholar] [CrossRef]
- Campbell, M.; Renshaw-Gegg, L.; Renne, R.; Luciw, P.A. Characterization of the internal promoter of simian foamy viruses. J. Virol. 1994, 68, 4811–4820. [Google Scholar] [CrossRef]
- Schweizer, M.; Fleps, U.; Jackle, A.; Renne, R.; Turek, R.; Neumann-Haefelin, D. Simian foamy virus type 3 (sfv-3) in latently infected vero cells: Reactivation by demethylation of proviral DNA. Virology 1993, 192, 663–666. [Google Scholar] [CrossRef]
- Clarke, J.K.; Samuels, J.; Dermott, E.; Gay, F.W. Carrier cultures of simian foamy virus. J. Virol. 1970, 5, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Hotta, J.; Loh, P.C. Enhanced production of a human spumavirus (retroviridae) in semi-permissive cell cultures after treatment with 5-azacytidine. J. Gen. Virol. 1987, Pt 4, 1183–1186. [Google Scholar] [CrossRef]
- Rhodes-Feuillette, A.; Mahouy, G.; Lasneret, J.; Flandrin, G.; Peries, J. Characterization of a human lymphoblastoid cell line permanently modified by simian foamy virus type 10. J. Med. Primatol. 1987, 16, 277–289. [Google Scholar] [PubMed]
- Falcone, V.; Leupold, J.; Clotten, J.; Urbanyi, E.; Herchenroder, O.; Spatz, W.; Volk, B.; Bohm, N.; Toniolo, A.; Neumann-Haefelin, D.; et al. Sites of simian foamy virus persistence in naturally infected african green monkeys: Latent provirus is ubiquitous, whereas viral replication is restricted to the oral mucosa. Virology 1999, 257, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.M.; Picker, L.J.; Axthelm, M.K.; Linial, M.L. Expanded tissue targets for foamy virus replication with simian immunodeficiency virus-induced immunosuppression. J. Virol. 2006, 80, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.M.; Picker, L.J.; Axthelm, M.K.; Hudkins, K.; Alpers, C.E.; Linial, M.L. Replication in a superficial epithelial cell niche explains the lack of pathogenicity of primate foamy virus infections. J. Virol. 2008, 82, 5981–5985. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Name | Source | Host Species | Tissue Origin | Cell Type |
|---|---|---|---|---|
| Mus dunni | [14] | wild mouse | tail | fibroblast |
| MDCK (NBL-2) | ATCC, CCL-34 | dog | kidney | epithelial |
| FRhK-4 | ATCC, CRL-1688 | rhesus macaque | fetal, kidney | epithelial |
| Vero | ATCC, CCL-81 | African green | kidney | epithelial |
| monkey | ||||
| MRC-5 | ATCC, CCL-171 | human | lung, fetal | fibroblast, |
| diploid | ||||
| A549 | ATCC, CCL-185 | human | lung, carcinoma | epithelial |
| Virus | Previous Designation | Species of Virus Isolation | Accession Number 1 |
|---|---|---|---|
| SFVmcy_FV21 | SFVmcy-1, (SFV serotype I) | Taiwanese macaque | NC_010819 |
| SFVmcy_FV34[RF] | SFVmcy-2, (SFV serotype II) | Taiwanese macaque | KF026286 |
| SFVmmu_K3T | SFVmmu-K3T | Rhesus macaque | KF026288 |
| SFVmmu_R289HybAGM[RF] | SFV-R289HybAGM | Rhesus macaque | JN801175 |
| SFVcae_FV2014 | SFV 3 | African green monkey | MF582544 |
| (SFV serotype III) | |||
| SFVcae_LK3 | SFVagm-3 | African green monkey | NC_010820 |
| SFVcae_agm4 | agm4 | African green monkey | AJ244075 |
| SFVcae_agm5 | agm5 | African green monkey | AJ244067 |
| SFVcae_agm20 | agm20 | African green monkey | AJ244091 |
| SFVcae_agm24 | agm24 | African green monkey | AJ244090 |
| SFVpve | SFVcpz | Chimpanzee | NC_001364 |
| SFVpsc_huHRSV.13 | HFV | Chimpanzee | KX08159 |
| SFVggo | SFVgor | Gorilla | HM245790 |
| BFVbta | BFV | Cow | NC_001831 |
| EFVeca_1 | EFVeca | Equine | AF201902 |
| FFVfca_FUV7 | FFVfca | Feline | Y08851 |
| Viral Regions | Location 1 | SFVcae_FV2014 LTR/gene 2 | ORF 3 | Location 1 | SFVcae_LK3 LTR/gene 2 | ORF 3 |
|---|---|---|---|---|---|---|
| LTR | 1–1710 | 1710 | 1–1708 | 1708 | ||
| gag | 1827–3758 | 1932 | 643 | 1825–3756 | 1932 | 643 |
| pol | 3706–7137 | 3432 | 1143 | 3704–7135 | 3432 | 1143 |
| env | 7085–10045 | 2961 | 986 | 7083–10031 | 2949 | 982 |
| tas | 10,015–10,905 | 891 | 296 | 10,001–10,897 | 897 | 298 |
| bet | 10,015–10,285, 10,581–11,824 | 1515 | 504 | 10,001–10,271, 10,567–11,705 | 1410 | 469 |
| LTR | 11,418–13,127 | 1710 | 11,404–13,111 | 1708 |
| SFV Isolates | % Sequence Identity | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LTR | gag | pol | env | env-LP | env-SU | env-TM | tas | bet | ||
| SFVcae_LK3 | nt | 88.71 | 81.5 | 86.3 | 79.1 | 79.6 | 71.1 | 86.6 | 90.2 | 88.8 |
| aa | 85.4 | 91.6 | 81.5 | 86.5 | 67.3 | 93.9 | 91.3 | 88.5 | ||
| SFVmcy_FV21 | nt | 66.9 | 68.1 | 81.7 | 75.0 | 71.4 | 73.8 | 77.0 | 66.1 | 61.8 |
| aa | 65.8 | 86.1 | 78.2 | 73.0 | 73.5 | 84.8 | 52.2 | 51.4 | ||
| SFVmcy_FV34(RF) | nt | 65.8 | 68.8 | 81.9 | 75.0 | 72.2 | 69.1 | 76.6 | 65.4 | 64.4 |
| aa | 66.7 | 86.3 | 78.2 | 73.8 | 63.2 | 83.9 | 52.5 | 50.9 | ||
| SFVmmu_R289(RF) | nt | 67.6 | 69.6 | 81.5 | 73.4 | 73.8 | 68.3 | 77.8 | 66.7 | 62.2 |
| aa | 67.6 | 86.4 | 74.5 | 76.2 | 63.7 | 85.3 | 53.7 | 51.2 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuentes, S.M.; Bae, E.H.; Nandakumar, S.; Williams, D.K.; Khan, A.S. Genome Analysis and Replication Studies of the African Green Monkey Simian Foamy Virus Serotype 3 Strain FV2014. Viruses 2020, 12, 403. https://doi.org/10.3390/v12040403
Fuentes SM, Bae EH, Nandakumar S, Williams DK, Khan AS. Genome Analysis and Replication Studies of the African Green Monkey Simian Foamy Virus Serotype 3 Strain FV2014. Viruses. 2020; 12(4):403. https://doi.org/10.3390/v12040403
Chicago/Turabian StyleFuentes, Sandra M., Eunhae H. Bae, Subhiksha Nandakumar, Dhanya K. Williams, and Arifa S. Khan. 2020. "Genome Analysis and Replication Studies of the African Green Monkey Simian Foamy Virus Serotype 3 Strain FV2014" Viruses 12, no. 4: 403. https://doi.org/10.3390/v12040403
APA StyleFuentes, S. M., Bae, E. H., Nandakumar, S., Williams, D. K., & Khan, A. S. (2020). Genome Analysis and Replication Studies of the African Green Monkey Simian Foamy Virus Serotype 3 Strain FV2014. Viruses, 12(4), 403. https://doi.org/10.3390/v12040403