Development of a Honey Bee RNA Virus Vector Based on the Genome of a Deformed Wing Virus

 ,
, 


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Construction of the egfp-Tagged DWV cDNA Clones and Generation of the Clone-Derived Viruses
2.2. Honey Bees
2.3. RT-qPCR and Virus Progeny Analysis
2.4. Detection of GFP and DWV Structural Proteins in Honey Bees by Western Blot and Immunofluorescence Microscopy
3. Results
3.1. Design of GFP-Tagged DWV cDNA Constructs
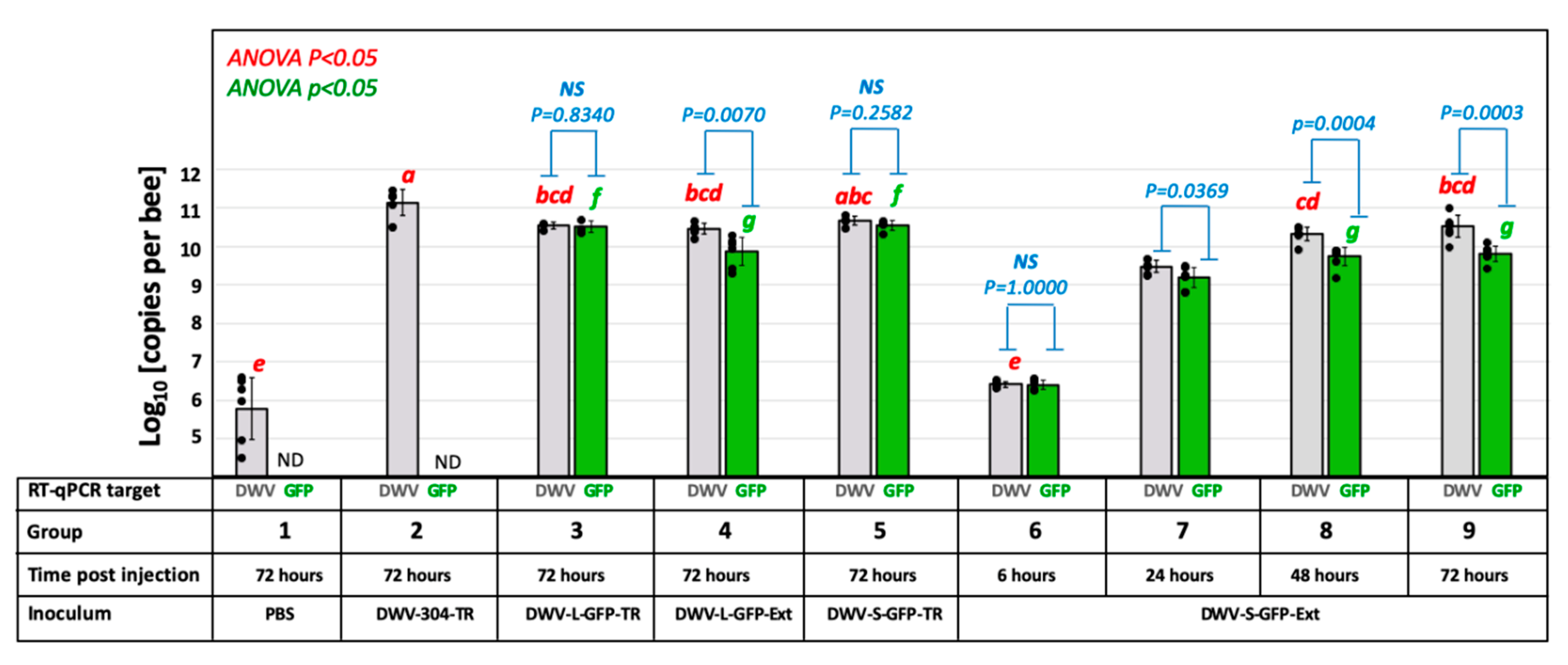
3.2. Infectivity the cDNA Clone-Derived GFP-Tagged DWV in Honey Bee Pupae
3.3. Analysis of Encapsidation of the egfp-Tagged DWV RNA Genomes
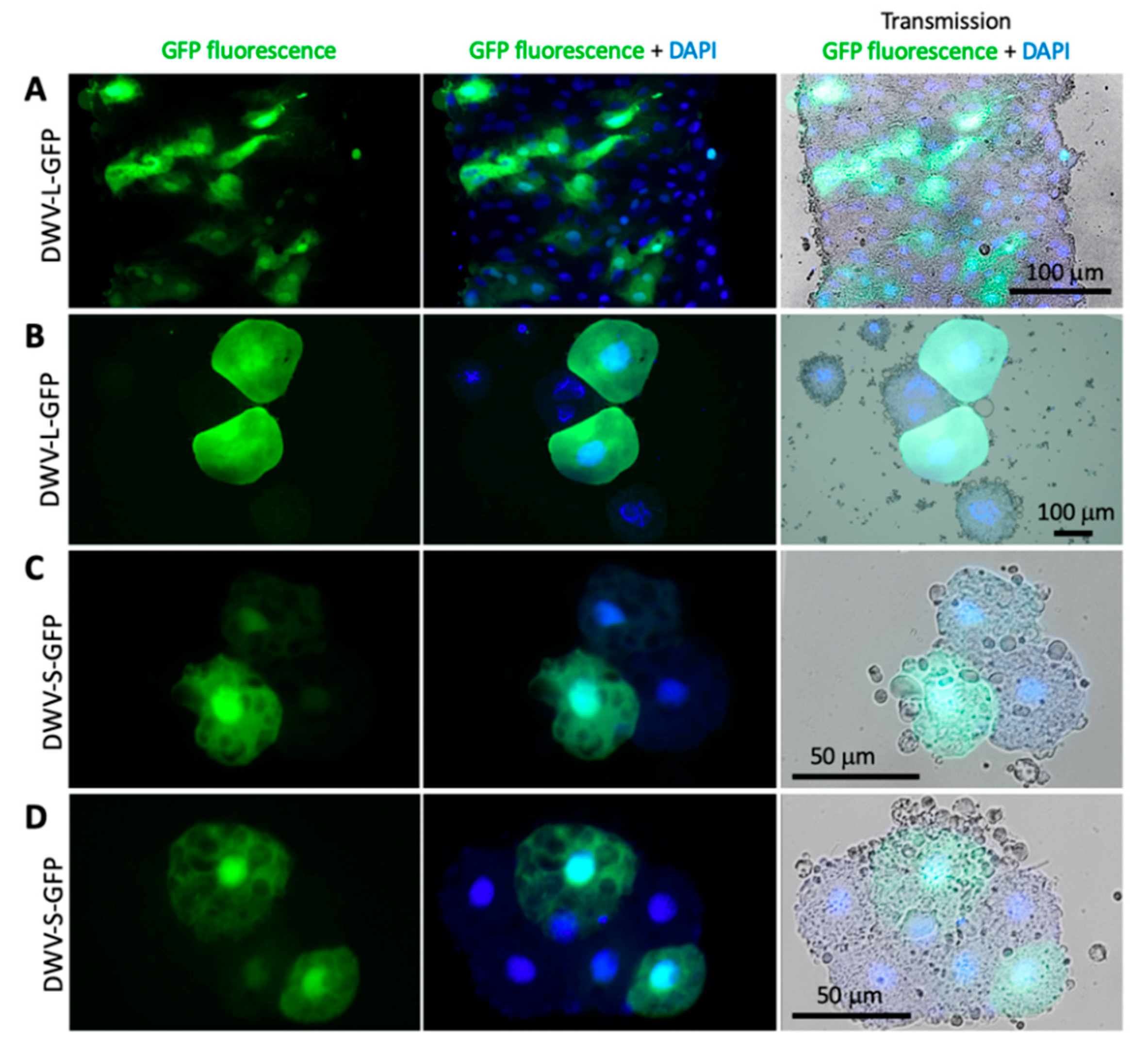
3.4. Expression of eGFP Reporter from the DWV RNA Vectors
3.5. Genetic Stability of egfp-Tagged DWV RNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mogler, M.A.; Kamrud, K.I. RNA-based viral vectors. Expert Rev. Vaccines 2014, 14, 283–312. [Google Scholar] [CrossRef] [PubMed]
- Ylä-Pelto, J.; Tripath, L.; Susi, P. Therapeutic use of native and recombinant enteroviruses. Viruses 2016, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Donson, J.; Kearney, C.M.; Hilf, M.E.; Dawson, W.O. Systemic expression of a bacterial gene by a Tobacco mosaic virus-based vector. Proc. Natl. Acad. Sci. USA 1991, 88, 7204–7208. [Google Scholar] [CrossRef] [PubMed]
- Pogue, G.P.; Lindbo, J.A.; Garger, S.J.; Fitzmaurice, W.P. Making an ally from an enemy: Plant virology and the new agriculture. Annu. Rev. Phytopathol. 2002, 40, 45–74. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D. VIGS vectors for gene silencing: Many targets, many tools. Annu. Rev. Plant Biol. 2004, 55, 495–519. [Google Scholar] [CrossRef] [PubMed]
- Oparka, K.J.; Boevink, P.; Santa Cruz, S. Studying the movement of plant viruses using green fluorescent protein. Trends Plant Sci. 1996, 12, 412–418. [Google Scholar] [CrossRef]
- Nouri, S.; Matsumura, E.E.; Kuo, Y.W.; Falk, B.W. Insect-specific viruses: From discovery to potential translational applications. Curr. Opin. Virol. 2018, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, R.; Cheng, L.-L.; Bartholomay, L.C.; Christensen, B.M. Flock house virus replicates and expresses green fluorescent protein in mosquitoes. J. Gen. Virol. 2003, 84, 1789–1797. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Andino, R. Non-specific dsRNA-mediated antiviral response in the honey bee. PLoS ONE 2013, 8, e77263. [Google Scholar] [CrossRef]
- Lanzi, G.; de Miranda, J.R.; Boniotti, M.B.; Cameron, C.E.; Lavazza, A.; Capucci, L.; Camazine, S.M.; Rossi, C. Molecular and biological characterization of deformed wing virus of honeybees (Apis mellifera L.). J. Virol. 2006, 80, 4998–5009. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Childers, A.K.; Lopez, D.; Grubbs, K.; Posada-Florez, F.; Weaver, D.; Girten, W.; vanEngelsdorp, D.; Chen, Y.; Evans, J.D. Dynamic evolution in the key honey bee pathogen deformed wing virus: Novel insights into virulence and competition using reverse genetics. PLoS Biol. 2019, 17, e3000502. [Google Scholar] [CrossRef] [PubMed]
- Cormack, B.P.; Valdivia, R.; Falkow, S. FACS-optimized mutants of the green fluorescent protein (GFP). Gene 1996, 173, 33–38. [Google Scholar] [CrossRef]
- Shi, X.; Harrison, R.L.; Hollister, J.R.; Mohammed, A.; Fraser, M.J.; Jarvis, D.L. Construction and characterization of new piggyBac vectors for constitutive or inducible expression of heterologous gene pairs and the identification of a previously unrecognized activator sequence in piggyBac. BMC Biotechnol. 2007, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Childers, A.K.; Chen, Y.; Madella, S.; Nessa, A.; vanEngelsdorp, D.; Evans, J.D. Recent spread of Varroa destructor virus-1, a honey bee pathogen, in the United States. Sci. Rep. 2017, 7, 17447. [Google Scholar] [CrossRef] [PubMed]
- Fannon, J.M.; Ryabov, E.V. Iflavirus (Deformed Wing Virus). In Molecular Detection of Animal Viral Pathogens; Liu, D., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 37–46. [Google Scholar]
- Kang, S.-H.; Bak, A.; Kim, O.-K.; Folimonova, S.Y. Membrane association of a nonconserved viral protein confers virus ability to extend its host range. Virology 2015, 482, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Škubník, K.; Nováček, J.; Füzik, T.; Přidal, A.; Paxton, R.J.; Plevka, P. Structure of deformed wing virus, a major honey bee pathogen. Proc. Natl. Acad. Sci. USA 2017, 114, 3210–3215. [Google Scholar] [CrossRef]
- Carrillo-Tripp, J.; Dolezal, A.G.; Goblirsch, M.J.; Miller, W.A.; Toth, A.L.; Bryony, C.; Bonning, B.C. In vivo and in vitro infection dynamics of honey bee viruses. Sci. Rep. 2016, 6, 22265. [Google Scholar] [CrossRef]
- Liu, Y.; Wimmer, E.; Paul, A.V. Cis-acting RNA elements in human and animal plus-strand RNA viruses. Biochim. Biophys. Acta 2009, 1789, 495–517. [Google Scholar] [CrossRef]
- Sasaki, J.; Taniguchi, K. The 5-end sequence of the genome of Aichi virus, a picornavirus, contains an element critical for viral RNA encapsidation. J. Virol. 2003, 77, 3542–3548. [Google Scholar] [CrossRef]
- Valles, S.M.; Chen, Y.; Firth, A.E.; Guérin, D.M.; Hashimoto, Y.; Herrero, S.; de Miranda, J.R.; Ryabov, E. ICTV Virus Taxonomy Profile: Iflaviridae. J. Gen. Virol. 2017, 98, 527–528. [Google Scholar] [CrossRef]
- Liu, H.; Wei, C.; Zhong, Y.; Li, Y. Rice black-streaked dwarf virus minor core protein P8 is a nuclear dimeric protein and represses transcription in tobacco protoplasts. FEBS Lett. 2007, 581, 2534–2540. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Brettell, L.E. Deformed Wing Virus in Honeybees and Other Insects. Annu. Rev. Virol. 2019, 6, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Qu, F.; Morris, T.J. Efficient Infection of Nicotiana benthamiana by Tomato bushy stunt virus is Facilitated by the Coat Protein and Maintained by p19 Through Suppression of Gene Silencing. MPMI 2002, 15, 193–202. [Google Scholar] [CrossRef]
- Qu, F.; Morris, T.J. Encapsidation of Turnip Crinkle Virus Is Defined by a Specific Packaging Signal and RNA Size. J. Virol. 1997, 71, 1428–1435. [Google Scholar] [CrossRef]
- Fievet, J.; Tentcheva, D.; Gauthier, L.; de Miranda, J.; Cousserans, F.; Colin, M.E.; Bergoin, M. Localization of deformed wing virus infection in queen and drone Apis mellifera L. Virol. J. 2006, 3, 16. [Google Scholar] [CrossRef]
- Lamp, B.; Url, A.; Seitz, K.; Eichhorn, J.; Riedel, C.; Sinn, L.J.; Indik, S.; Köglberger, H.; Rümenapf, T. Construction and Rescue of a Molecular Clone of Deformed Wing Virus (DWV). PLoS ONE 2016, 11, e0164639. [Google Scholar] [CrossRef]
- Posada-Florez, F.; Ryabov, E.V.; Heerman, M.C.; Chen, Y.; Evans, J.D.; Cook, S.C.; Sonenshine, S.C. A novel system for maintaining Varroa destructor mites on artificial diets and its application for studying mites as a vector for honey bee viruses. BioRxiv 2019. [Google Scholar] [CrossRef]
- Gusachenko, O.N.; Woodford, L.; Balbirnie-Cumming, K.; Ryabov, E.V.; Evans, D.J. Deformed Wing Virus spillover from honey bees to bumble bees: A reverse genetic study. BioRxiv 2019. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Genersch, E. RT-PCR analysis of Deformed wing virus in honeybees (Apis mellifera) and mites (Varroa destructor). J. Gen. Virol. 2005, 86, 3419–3424. [Google Scholar] [CrossRef] [PubMed]
- Santillán-Galicia, M.T.; Carzaniga, R.; Ball, B.B.; Alderson, P.G. Immunolocalization of deformed wing virus particles within the mite Varroa destructor. J. Gen. Virol. 2008, 89, 1685–1689. [Google Scholar] [CrossRef] [PubMed]
- Gisder, S.; Aumeier, P.; Genersch, E. Deformed wing virus: Replication and viral load in mites (Varroa destructor). J. Gen. Virol. 2009, 90, 463–467. [Google Scholar] [CrossRef]
- Posada-Florez, F.; Childers, A.K.; Heerman, M.C.; Egekwu, N.I.; Cook, S.C.; Chen, Y.; Evans, J.D.; Ryabov, E.V. Deformed wing virus type A, a major honey bee pathogen, is vectored by the mite Varroa destructor in a non-propagative manner. Sci. Rep. 2019, 9, 12445. [Google Scholar] [CrossRef]
- Dobrikova, E.Y.; Florez, P.; Gromeier, M. Structural determinants of insert retention of poliovirus expression vectors with recombinant IRES elements. Virology 2003, 311, 241–253. [Google Scholar] [CrossRef][Green Version]
- Chung, B.-N.; Tomas Canto, T.; Palukaitis, P. Stability of recombinant plant viruses containing genes of unrelated plant viruses. J. Gen. Virol. 2007, 88, 1347–1355. [Google Scholar] [CrossRef]
- Tromas, N.; Zwart, M.P.; Forment, J.; Elena, S.F. Shrinkage of Genome Size in a Plant RNA Virus upon Transfer of an Essential Viral Gene into the Host Genome. Genome Biol. Evol. 2014, 6, 538–550. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V.; Krupovic, M. Origins and evolution of viruses of eukaryotes: The ultimate modularity. Virology 2015, 479, 2–25. [Google Scholar] [CrossRef]
- Elena, S.F. Evolutionary transitions during RNA virus experimental evolution. Philos. Trans. R. Soc. B 2016, 371, 20150441. [Google Scholar] [CrossRef]
- Leonard, S.P.; Powell, J.E.; Perutka, J.; Geng, P.; Heckmann, L.C.; Horak, R.D.; Davies, B.W.; Ellington, A.D.; Barrick, J.E.; Moran, N.A. Engineered symbionts activate honey bee immunity and limit pathogens. Science 2020, 367, 573–576. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryabov, E.V.; Christmon, K.; Heerman, M.C.; Posada-Florez, F.; Harrison, R.L.; Chen, Y.; Evans, J.D. Development of a Honey Bee RNA Virus Vector Based on the Genome of a Deformed Wing Virus. Viruses 2020, 12, 374. https://doi.org/10.3390/v12040374
Ryabov EV, Christmon K, Heerman MC, Posada-Florez F, Harrison RL, Chen Y, Evans JD. Development of a Honey Bee RNA Virus Vector Based on the Genome of a Deformed Wing Virus. Viruses. 2020; 12(4):374. https://doi.org/10.3390/v12040374
Chicago/Turabian StyleRyabov, Eugene V., Krisztina Christmon, Matthew C. Heerman, Francisco Posada-Florez, Robert L. Harrison, Yanping Chen, and Jay D. Evans. 2020. "Development of a Honey Bee RNA Virus Vector Based on the Genome of a Deformed Wing Virus" Viruses 12, no. 4: 374. https://doi.org/10.3390/v12040374
APA StyleRyabov, E. V., Christmon, K., Heerman, M. C., Posada-Florez, F., Harrison, R. L., Chen, Y., & Evans, J. D. (2020). Development of a Honey Bee RNA Virus Vector Based on the Genome of a Deformed Wing Virus. Viruses, 12(4), 374. https://doi.org/10.3390/v12040374